Recovery from Hypersaline-Stress-Induced Immunity Damage and Intestinal-Microbiota Changes through Dietary β-glucan Supplementation in Nile tilapia (Oreochromis niloticus)





Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Diets
2.3. Experimental Fish and Management Procedure
2.4. Sample Collection
2.5. Hematological Assay
2.6. Immune-Related Gene Expression
2.7. Intestinal Microbiota Analysis in Nile tilapia
2.8. Statistical Analysis
3. Results
3.1. Survival, Growth Performance, and Relative Spleen Weight
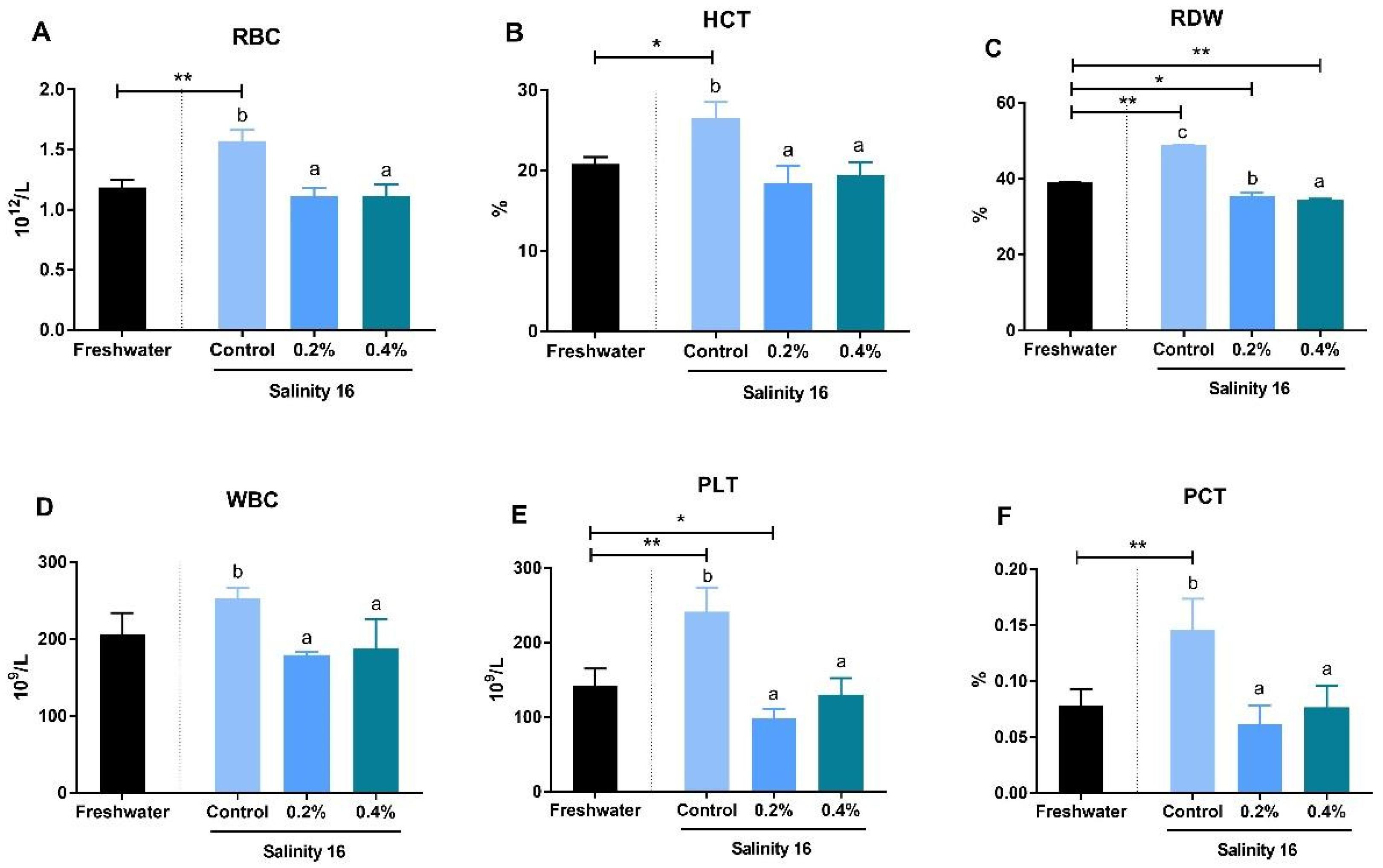
3.2. Hematological Parameters
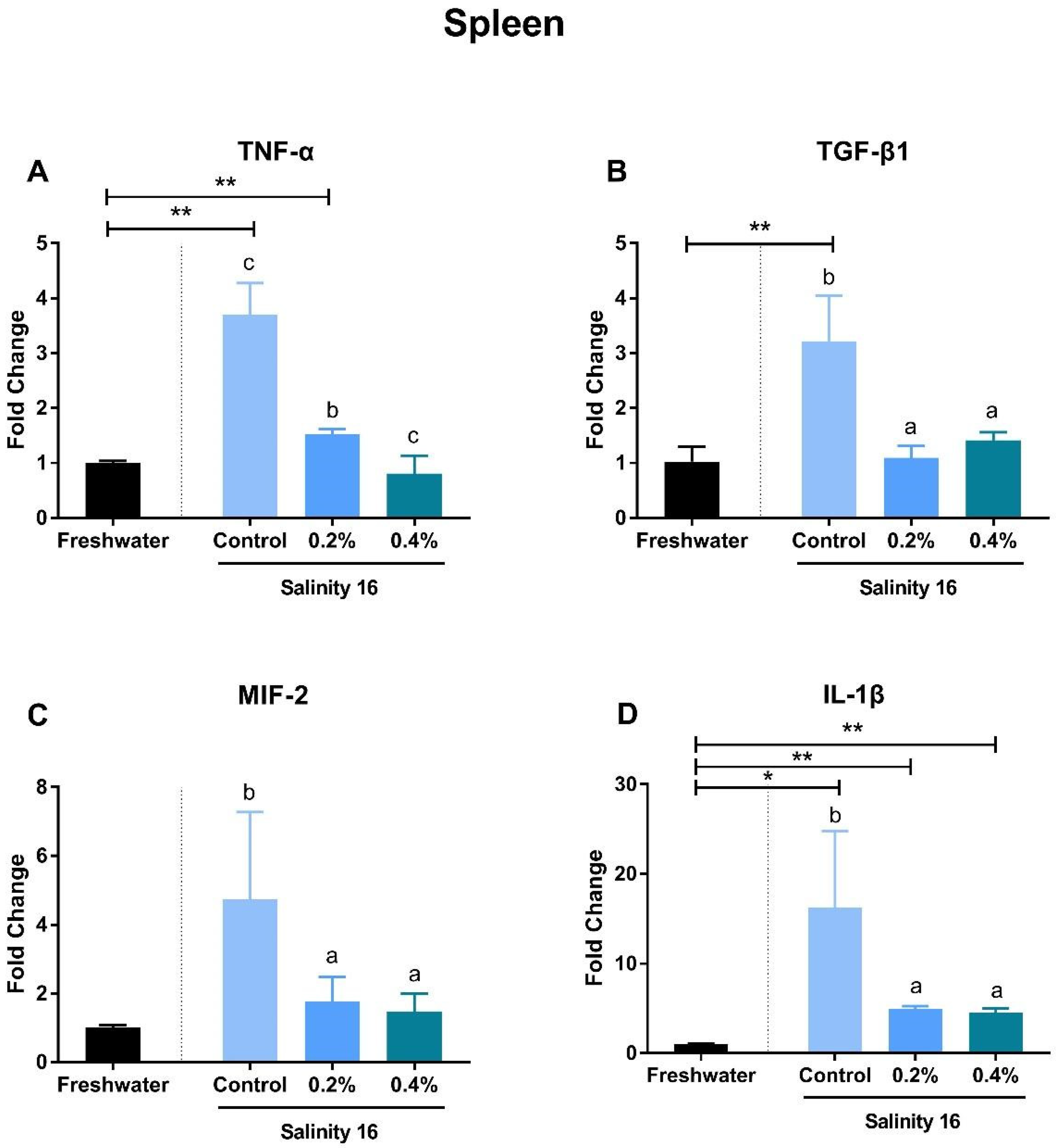
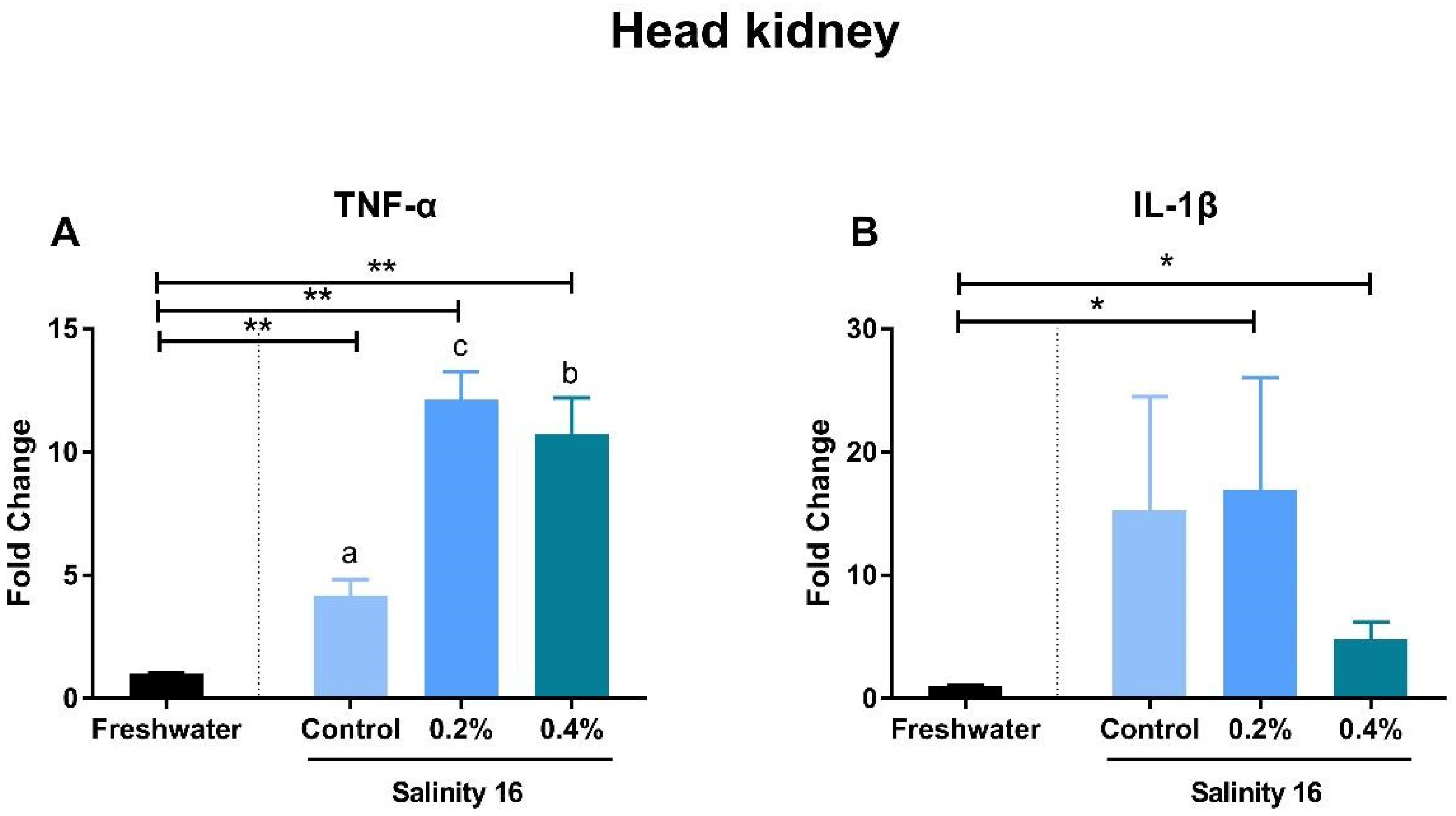
3.3. Gene Expression of Immune-Inflammatory Cytokines
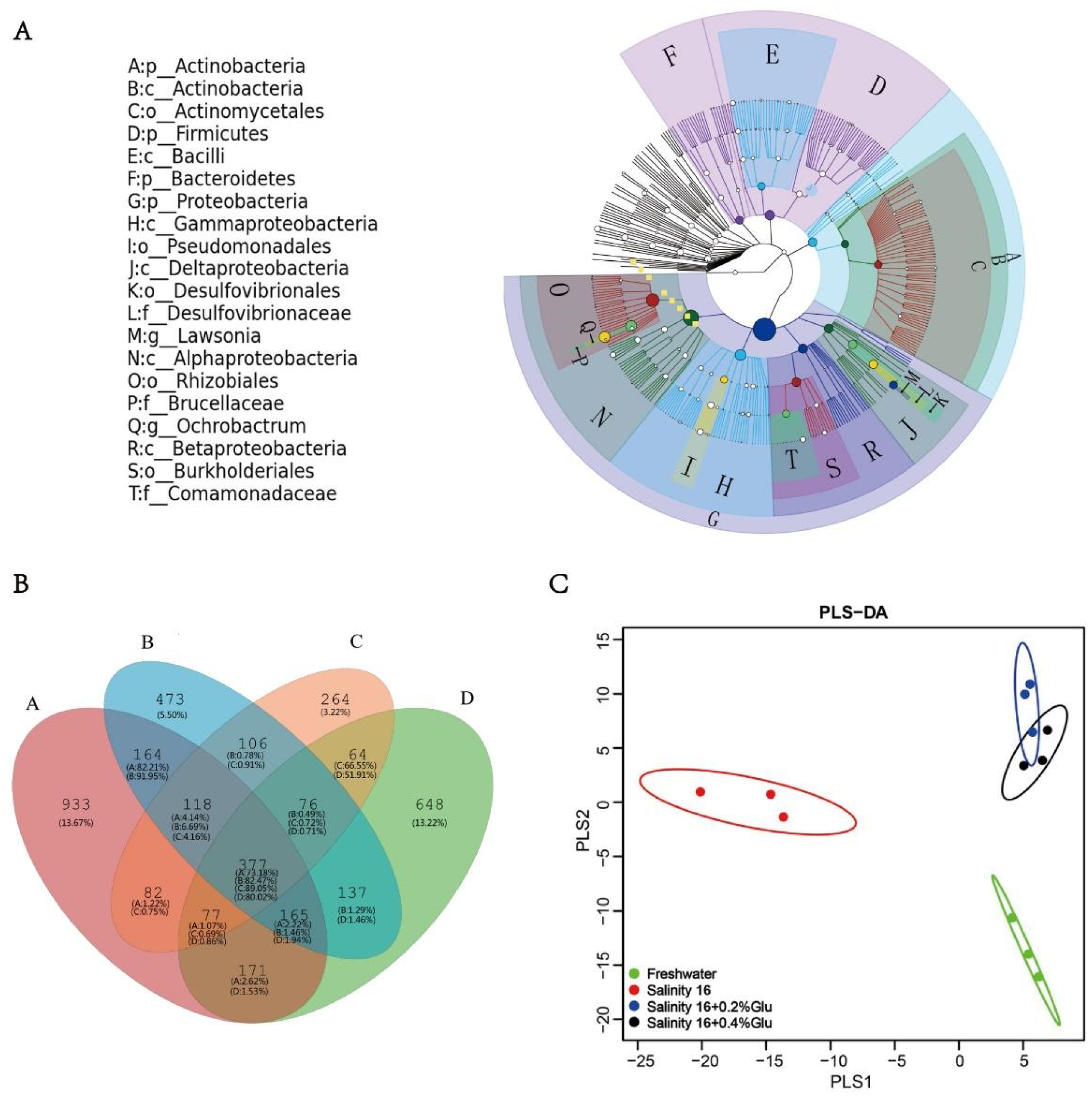
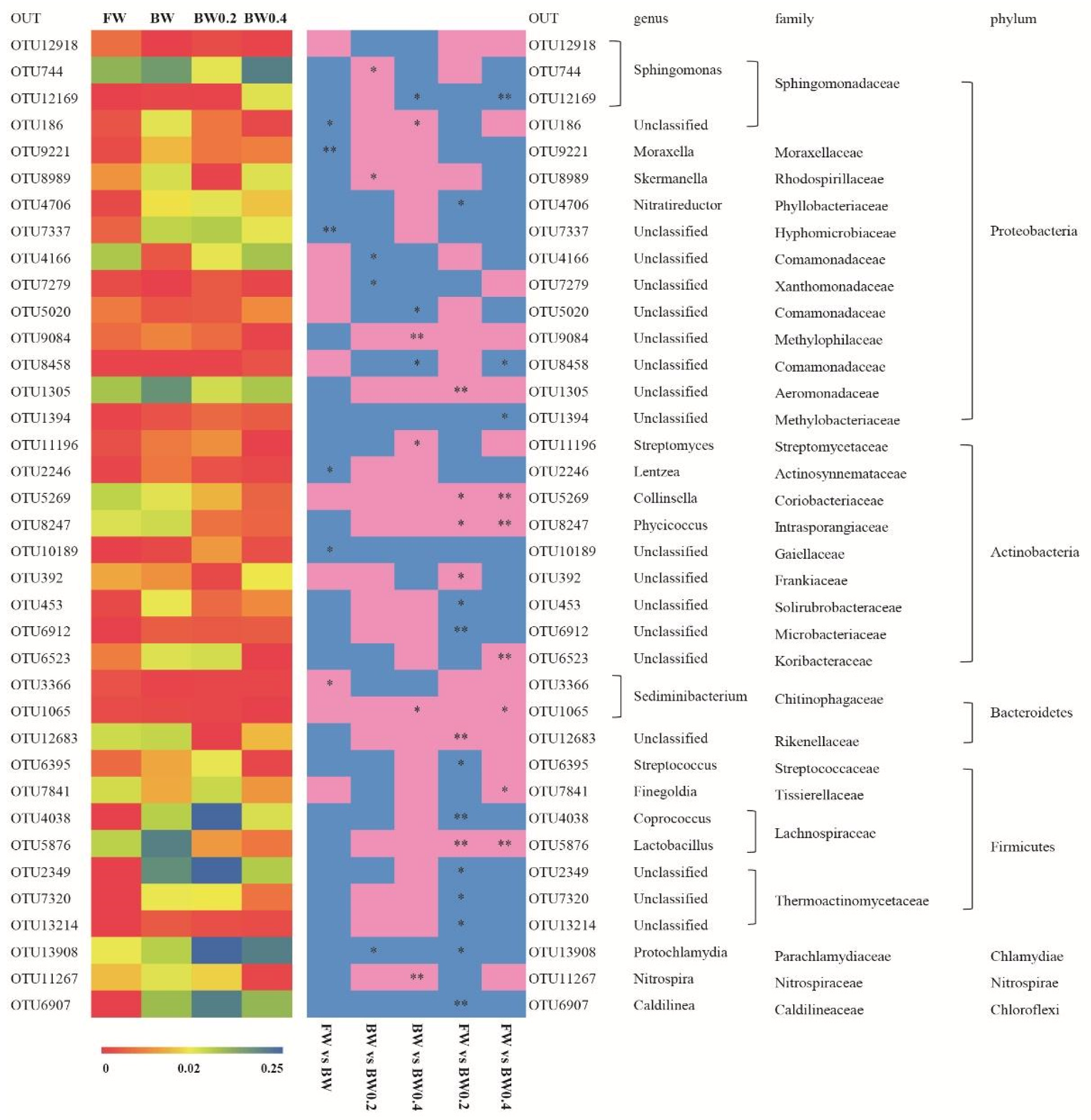
3.4. Differences in Bacterial Community Composition, Diversity, and Structure
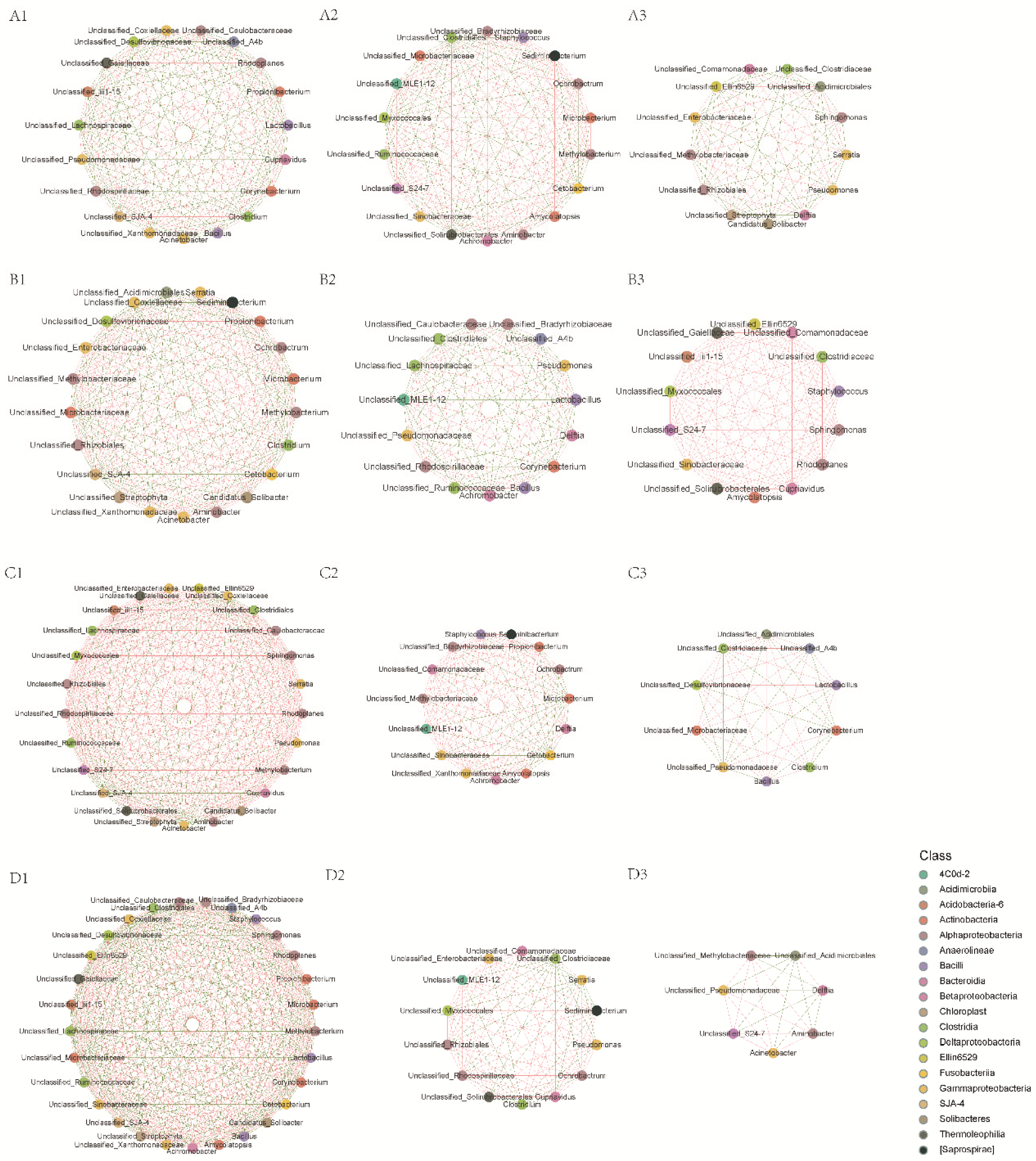
3.5. Intestinal Microbiota Interspecies Interaction
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vetvicka, V.J.; Vetvickova, J. Physiological effects of different types of β-glucan. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 2007, 151, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-Glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Sun, Y.; Zou, S.W.; Duan, B.C.; Sun, M.Y.; Xu, X.J. Yeast β-glucan suppresses the chronic inflammation and improves the microenvironment in adipose tissues of ob/ob mice. J. Agric. Food Chem. 2018, 66, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.F.; Chan, W.K.; Sze, M.Y. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilarski, F.; de Oliveira, C.A.F.; de Souza, F.P.B.D.; Zanuzzo, F.S. Different β-glucans improve the growth performance and bacterial resistance in Nile tilapia. Fish Shellfish Immunol. 2017, 70, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.A.O.; Eweedah, N.M.; Moustafa, E.M.; Shahin, M.G. Effects of aspergillus oryzae and β-glucan on growth and oxidative and immune responses of Nile tilapia, Oreochromis niloticus. Probiotics Antimicro. 2019, 425, 9. [Google Scholar] [CrossRef]
- Udayangani, R.; Dananjaya, S.; Fronte, B.; Kim, C.H.; Lee, J.; Zoysa, M.D. Feeding of nano scale oats β-glucan enhances the host resistance against Edwardsiella tarda and protective immune modulation in zebrafish larvae. Fish Shellfish Immunol. 2017, 60, 72–77. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; El Basuini, M.F.; Hossain, M.S.; Nhu, T.H.; Moss, A.S.; Dossou, S.; Wei, H. Dietary supplementation of β-glucan improves growth performance, the innate immune response and stress resistance of red sea bream, Pagrus major. Aquac. Nurt. 2017, 23, 148–159. [Google Scholar] [CrossRef]
- Chen, D.; Ainsworth, A.J. Glucan administration potentiates immune defence mechanisms of channel catfish, Ictalurus punctatus Rafinesque. J. Fish. Dis. 2006, 15, 295–304. [Google Scholar] [CrossRef]
- Ai, Q.H.; Mai, K.S.; Zhang, L.; Tan, B.P.; Zhang, W.B.; Xu, W.; Li, H.T. Effects of dietary β-1, 3 glucan on innate immune response of large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2007, 22, 394–402. [Google Scholar] [CrossRef]
- Murthy, H.S.; Li, P.; Lawrence, A.L.; Gatlin, D.M. Dietary β-glucan and nucleotide effects on growth, survival and immune responses of pacific white shrimp, Litopenaeus vannamei. J. Appl. Aquac. 2009, 21, 160–168. [Google Scholar] [CrossRef]
- Li, H.F.; Xu, C.; Zhou, L.; Dong, Y.F.; Su, Y.J.; Wang, X.D.; Jian, G.Q.I.N.; Chen, L.Q.; Li, E.C. Beneficial effects of dietary beta-glucan on growth and health status of Pacific white shrimp Litopenaeus vannamei at low salinity. Fish Shellfish Immunol. 2019, 91, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Chen, J.L.; Chung, S.S.M.; Xu, B.J. A critical review on the impacts of β-glucans on gut microbiota and human health. J. Nutr. Bio. 2018, 61, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Sima, P.; Vannucci, L.; Vetvicka, V. β-glucans and cholesterol (Review). Int. J. Mol. Med. 2018, 41, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.O.; Flint, H.J.; Duncan, S.H. The influence of diet on the gut microbiota. Pharmacol. Res. 2013, 69, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Li, E.C.; Xu, C.; Wang, X.D.; Wang, S.F.; Zhao, Q.; Zhang, M.L.; Jian, G.Q.I.N.; Chen, L.Q. Gut microbiota and its modulation for healthy farming of Pacific white shrimp, Litopenaeus vannamei. Rev. Fish. Sci. Aquac. 2018, 26, 381–399. [Google Scholar] [CrossRef]
- Jung-Schroers, V.; Adamek, M.; Jung, A.; Harris, S.; Harris, S.; Dóza, Ö.-S.; Baumer, A.; Steinhagen, D. Feeding of β-1,3/1,6-glucan increases the diversity of the intestinal microflora of carp (Cyprinus carpio). Aquacult Nutr. 2015, 22, 1026–1039. [Google Scholar] [CrossRef]
- Miest, J.J.; Arndt, C.; Adamek, M.; Steinhagen, D.; Reusch, T.B.H. Dietary β-glucan (MacroGard®) enhances survival of first feeding turbot (Scophthalmus maximus) larvae by altering immunity, metabolism and microbiota. Fish Shellfish Immunol. 2016, 48, 94–104. [Google Scholar] [CrossRef]
- Rufchaie, R.; Hoseinifar, S.H. Effects of dietary commercial yeast glucan on innate immune response, hematological parameters, intestinal microbiota and growth performance of white fish (Rutilus frisii kutum) fry. Croat. J. Fisheries. 2014, 72, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Kühlwein, H.; Emery, M.J.; Rawling, M.D.; Harper, G.M.; Merrifield, S.J.; Davies, S.J. Effects of a dietary β-(1, 3) (1, 6)-D-glucan supplementation on intestinal microbial communities and intestinal ultrastructure of mirror carp (Cyprinus carpio L.). J. Appl. Microbiol. 2013, 115, 1091–1106. [Google Scholar] [CrossRef]
- Wongsasak, U.; Chaijamrus, S.; Kumkhong, S.; Boonanuntanasarn, S. Effects of dietary supplementation with β-glucan and synbiotics on immune gene expression and immune parameters under ammonia stress in Pacific white shrimp. Aquaculture 2015, 436, 179–187. [Google Scholar] [CrossRef]
- Brocker, C.; Thompson, D.C.; Vasilis, V. The role of hyperosmotic stress in inflammation and disease. Biomol. Concepts 2012, 3, 345–364. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Li, E.C.; Suo, Y.T.; Su, Y.J.; Lu, M.H.; Zhao, Q.; Jian, G.Q.I.N.; Chen, L.Q. Histological and transcriptomic responses of two immune organs, the spleen and head kidney, in Nile tilapia (Oreochromis niloticus) to long-term hypersaline stress. Fish Shellfish Immunol. 2018, 76, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, A.; Diez, B.; Cejalvo, T.; Gutiérrezde, F.C.; Cortés, A. Ontogeny of the immune system of fish. Fish Shellfish Immunol. 2006, 20, 126–136. [Google Scholar] [CrossRef]
- Fishelson, L. Cytomorphological alterations of the thymus, spleen, head-kidney, and liver in cardinal fish (Apogonidae, Teleostei) as bioindicators of stress. J. Morphol. 2006, 267, 57–69. [Google Scholar] [CrossRef]
- Jiang, I.F.; Kumar, V.B.; Lee, D.N.; Weng, C.F. Acute osmotic stress affects Tilapia (Oreochromis mossambicus) innate immune responses. Fish Shellfish Immunol. 2008, 25, 841–846. [Google Scholar] [CrossRef]
- Tellez-Bañuelos, M.C.; Santerre, A.; Casas-Solis, J.; Bravo-Cuellar, A.; Zaitseva, G. Oxidative stress in macrophages from spleen of Nile tilapia (Oreochromis niloticus) exposed to sublethal concentration of endosulfan. Fish Shellfish Immunol. 2009, 27, 105–111. [Google Scholar] [CrossRef]
- Hussein, M.M.A.; Elsadaawy, H.A.; El-Murr, A.; Ahmed, M.M.; Bedawy, A.M.; Tukur, H.A.; Swelum, A.A.A.; Saadeldin, I.M. Endosulfan toxicity in Nile tilapia (Oreochromis niloticus) and the use of lycopene as an ameliorative agent. Comp. Bioche. Phys. C 2019, 224, 108573. [Google Scholar] [CrossRef]
- Suresh, N. Effect of cadmium chloride on liver, spleen and kidney melano macrophage centres in Tilapia mossambica. J. Environ. Biol. 2009, 30, 505–508. [Google Scholar] [CrossRef]
- Zarejabad, A.M.; Jalali, M.A.; Sudagar, M.; Pouralimotlagh, S. Hematology of great sturgeon (Huso huso Linnaeus, 1758) juvenile exposed to brackish water environment. Fish. Physiol. Biochem. 2010, 36, 655–659. [Google Scholar] [CrossRef]
- Xu, L.W.; Feng, J.; Guo, Z.X.; Lin, H.Z.; Guo, G.X. Effect of salinity on hematology and gill Na+-K+ ATPase activity of juvenile cobia, Rachycentron canadum Linnaeus. Mar. Environ. Sci. 2008, 27, 602–606. [Google Scholar] [CrossRef]
- Kavya, K.S.; Jadesh, M.; Kulkarni, R.S. Hematology and serum biochemical changes in response to change in saline concentration in fresh water fish Notopterus notopterus. World Sci. News 2016, 32, 49–60. [Google Scholar]
- Jarvis, P.L.; Ballantyne, J.S. Metabolic responses to salinity acclimation in juvenile shortnose sturgeon Acipenser brevirostrum. Aquaculture 2003, 219, 891–909. [Google Scholar] [CrossRef]
- Zhang, M.L.; Sun, Y.H.; Liu, Y.K.; Qiao, F.; Chen, L.Q.; Liu, W.T.; Du, Z.Y.; Li, E.C. Response of gut microbiota to salinity change in two euryhaline aquatic animals with reverse salinity preference. Aquaculture 2016, 454, 72–80. [Google Scholar] [CrossRef]
- Readman, G.D.; Owen, S.F.; Knowles, T.G.; Murrell, J.C. Species specific anaesthetics for fish anaesthesia and euthanasia. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laria, C.; Saoca, C.; Guerrera, M.C.; Ciulli, S.; Brundo, M.V.; Piccione, G.; Lanteri, G. Occurrence of diseases in fish used for experimental research. Lab. Anim. 2019, 53, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.K.; Singh, S.K.; Akhtar, M.S.; Behera, B.K.; Kumar, K.; Pal, A.K.; et al. Beta-glucan: An ideal immunostimulant in aquaculture (a review). Fish. Physiol. Biochem. 2013, 39, 431–457. [Google Scholar] [CrossRef]
- Lin, S.M.; Pan, Y.; Luo, L.; Luo, L. Effects of dietary β-1,3-glucan, chitosan or raffinose on the growth, innate immunity and resistance of koi (Cyprinus carpio koi). Fish Shellfish Immunol. 2011, 31, 788–794. [Google Scholar] [CrossRef]
- Ji, L.Q.; Sun, G.X.; Li, J.; Wang, Y.; Du, Y.S.; Li, X.; Liu, Y. Effect of dietary β-glucan on growth, survival and regulation of immune processes in rainbow trout (Oncorhynchus mykiss) infected by, Aeromonas salmonicida. Fish Shellfish Immunol. 2017, 64, 56–67. [Google Scholar] [CrossRef]
- El-Murr Abd elhakeem, I.; Hakim, Y.A.E.; Neamat-Allah, A.N.F.; Baeshen, M.; Ali, H.A. Immune-protective, antioxidant and relative genes expression impacts of β-glucan against fipronil toxicity in Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2019, 94, 427–433. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Ma, H.M.; Zhang, W.B.; Ai, Q.H.; Mai, K.S.; Xu, W.; Wang, X.J.; Liu, Z.G. Effects of dietary β-glucan on the growth, immune responses and resistance of sea cucumber, Apostichopus japonicus against Vibrio splendidus infection. Aquaculture 2011, 315, 269–274. [Google Scholar] [CrossRef]
- Divya, M.; Gopi, N.; Iswarya, A.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Almanaa, T.N.; Vaseeharan, B. β-glucan extracted from eukaryotic single-celled microorganism Saccharomyces cerecisiae: Dietary supplementation and enhanced ammonia stress tolerance on Oreochromis mossambicus. Microb. Pathog. 2020, 139, 103917. [Google Scholar] [CrossRef] [PubMed]
- Bagni, M.; Romano, N.; Finoia, M.G.; Abelli, L.; Scapigliati, G.; Tiscar, P.G.; Sarti, M.; Marino, G. Short- and long-term effects of a dietary yeast beta-glucan (Macrogard) and alginic acid (Ergosan) preparation on immune response in sea bass (Dicentrarchus labrax). Fish Shellfish Immunol. 2005, 18, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.L.; Limbu, S.M.; Lv, H.B.; Ma, Q.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. The comparisons in protective mechanisms and efficiencies among dietary α-lipoic acid, β-glucan and l-carnitine on Nile tilapia infected by Aeromonas hydrophila. Fish Shellfish Immunol. 2018, 86, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.Z.; Yu, R.H.; Zhang, Y.Y.; Hu, B.Q.; Jian, S.Q.; Wen, C.G.; Kajbaf, K.; Kumar, V.; Yang, G. Effects of dietary supplementation with β-glucan and Bacillus subtilis on growth, fillet quality, immune capacity, and antioxidant status of Pengze crucian carp (Carassius auratus var. Pengze). Aquaculture 2019, 508, 106–112. [Google Scholar] [CrossRef]
- Abdelhamid, F.M.; Elshopakey, G.E.; Aziza, A.E. Ameliorative effects of dietary Chlorella vulgaris and β-glucan against diazinon-induced toxicity in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 96, 213–222. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486–503. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, C.A.F.; Vetvicka, V.; Zanuzzo, F.S. β-Glucan successfully stimulated the immune system in different jawed vertebrate species. Comp. Immunol. Diagn. Microbiol. Infect. Dis. 2019, 62, 1–6. [Google Scholar] [CrossRef]
- Huu, H.D.; Sang, H.M.; Thanh, T.N.T. Dietary β-glucan improved growth performance, Vibrio counts, haematological parameters and stress resistance of pompano fish, Trachinotus ovatus Linnaeus, 1758. Fish Shellfish Immunol. 2016, 54, 402–410. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Mukherjee, S.C. Effect of dietary beta-1,3 glucan on immune responses and disease resistance of healthy and aflatoxin B1-induced immunocompromised rohu (Labeo rohita Hamilton). Fish Shellfish Immunol. 2001, 11, 683–695. [Google Scholar] [CrossRef]
- Flavell, R.A. The relationship of inflammation and initiation of autoimmune disease: Role of TNF super family members. Curr. Top. Microbiol. Immun. 2002, 266, 1–9. [Google Scholar] [CrossRef]
- Pallai, A.; Kiss, B.; Vereb, G.; Armaka, M.; Kollias, G.; Szekanecz, Z.; Szondy, Z. Transmembrane TNF—Reverse signaling inhibits lipopolysaccharide-induced proinflammatory cytokine formation in macrophages by inducing TGF-: Therapeutic implications. J. Immunol. 2016, 196, 1146–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netea, M.G.; Simon, A.; De Veerdonk, F.V.; Kullberg, B.J.; Van der Meer, J.W.; Joosten, L.A. IL-1beta processing in host defense: Beyond the inflammasomes. PLoS Pathol. 2010, 6, e1000661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetz, F.W.; Planas, J.V.; MacKenzie, S. Tumor necrosis factors. Dev. Compar. Immunol. 2004, 28, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Frost, P.; Miest, J.; Pionnier, N.; Irnazarow, I.; Hoole, D. Reduced inflammatory response to Aeromonas salmonicida infection in common carp (Cyprinus carpio L.) fed with β-glucan supplements. Fish Shellfish Immunol. 2012, 32, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, A.; De Baetselier, P.; Roelants, K.; Trez, C.D.; Magez, S.; Ginderachter, J.A.V.; Raes, G.; Bucala, R.; Stijlemans, B. The non-mammalian MIF superfamily. Immunobiology 2016, 222, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Douxfils, J.; Fierro-Castro, C.; Mandiki, S.N.M.; Emile, W.; Tort, L.; Kestemont, P. Dietary β-glucans differentially modulate immune and stress-related gene expression in lymphoid organs from healthy and, Aeromonas hydrophila-infected rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2017, 63, 285–296. [Google Scholar] [CrossRef]
- Carlos, C.V.; Rodriguez, M.E.M.; Arizaga, G.G.C.; Jara, J.S.; Angulo, C.; Becerril, M.R. Immobilizing yeast β-glucan on zinc-layered hydroxide nanoparticle improves innate immune response in fish leukocytes. Fish Shellfish Immunol. 2018, 82, 504–513. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Ganzle, M.G.; Mosenthin, R.; Zijlstra, R.T. Oat β-glucan and dietary calcium and phosphorus differentially modify intestinal expression of proinflammatory cytokines and monocarboxylate transporter 1 and cecal morphology in weaned pigs. J. Nutr. 2012, 142, 668–674. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, R.F.; Berthon, B.S.; Jensen, M.E.; Baines, K.J.; Wood, L.G. Short-chain fatty acids, prebiotics, synbiotics, and systemic inflammation: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2017, 106, 930–945. [Google Scholar] [CrossRef] [Green Version]
- Carballo, C.; Pinto, P.I.S.; Mateus, A.P.; Berbel, C.; Guerreiro, C.C.; Martinez-Blanch, J.F.; Codoner, F.M.; Mantecon, L.; Power, D.M.; Manchado, M. Yeast β-glucan and microalgal extracts modulate the immune response and gut microbiome in Senegalese sole (Solea senegalensis). Fish Shellfish Immunol. 2019, 92, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xu, Z.J.; Tian, X.L.; Dong, S.L.; Peng, M. Intestinal microbiota and immune related genes in sea cucumber (Apostichopus japonicus) response to dietary β-glucan supplementation. Biochem. Bioph. Res. Co. 2015, 458, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Siriyappagouder, P.; Galindo-Villegas, J.; Lokesh, J.; Mulero, V.; Fernandes, J.M.O.; Kiron, V. Exposure to yeast shapes the intestinal bacterial community assembly in zebrafish larvae. Front. Microbiol. 2018, 9, 1868. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, C.Z.; Xie, J.; Xu, C.; Zhao, Q.; Jian, G.Q.I.N.; Chen, L.Q.; Li, E.C. Intestinal bacterial signatures of the “cotton shrimp-like” disease explain the change of growth performance and immune responses in Pacific white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2019, 92, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Gatesoupe, F.J. Updating the importance of lactic acid bacteria in fish farming: Natural occurrence and pro-biotic treatments. J. Mol. Microb. Biotech. 2008, 14, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaskari, J.; Kontula, P.; Siitonen, A.; Somer, H.J.; Sandholm, T.M.; Poutanen, K. Oat β-glucan and xylan hydrolysates as selective substrates for Bifidobacterium and Lactobacillus strains. App. Microbiol. Biotechnol. 1998, 49, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Foysal, M.J.; Alam, M.; Kawser, A.Q.M.R.; Hasan, F.; Rahman, M.M.; Tay, C.Y.; Prodhan, M.S.H.; Gupta, S.K. Meta-omics technologies reveals beneficiary effects of Lactobacillus plantarum as dietary supplements on gut microbiota, immune response and disease resistance of Nile tilapia (Oreochromis niloticus). Aquaculture 2020, 520, 734974. [Google Scholar] [CrossRef]
- Johnson, D.B. Biodiversity and ecology of acidophilic microorganisms. FEMS Microbiol. Ecol. 1998, 27, 307–317. [Google Scholar] [CrossRef]
- Yang, X.Z.; Xu, M.J.; Huang, G.Y.; Zhang, C.; Pang, Y.Y.; Cheng, Y.X. Effect of dietary L-tryptophan on the survival, immune response and gut microbiota of the Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2019, 84, 1007–1017. [Google Scholar] [CrossRef]
- Yu, L.; Rahman, S.U.; Huang, Y.Y.; Zhang, Y.F.; Ming, P.F.; Zhu, L.; Chu, X.Y.; Li, J.C.; Feng, S.B.; Wang, X.H.; et al. Green tea polyphenols decrease weight gain, ameliorate alteration of gut microbiota, and mitigate intestinal inflammation in canines with high-fat-diet-induced obesity. J. Nutr. Biochem. 2020, 78, 108324. [Google Scholar] [CrossRef]
- Tzelepis, G.; Karlsson, M. Killer toxin-like chitinases in filamentous fungi: Structure, regulation and potential roles in fungal biology. Fungal Biol. Rev. 2018, 33, 123–132. [Google Scholar] [CrossRef]
- Oh, M.; Zhang, T.; Seljelid, R.; Li, X. Treptomyces cerevisiae 1,3/1,6 beta-glucan prevents airway hyperreactivity and pulmonary inflammation in a murine asthma model. J. Allergy Clin. Immunol. 2007, 119, S6. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black Soldier Fly (Hermetia illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2019, 518, 734659. [Google Scholar] [CrossRef]
- Zheng, Y.; Wu, W.; Hu, G.D.; Qin, L.P.; Meng, S.L.; Song, C.; Fan, L.M.; Zhao, Z.X.; Bing, X.W.; Chen, J.Z. Gut microbiota analysis of juvenile genetically improved farmed tilapia (Oreochromis niloticus) by dietary supplementation of different resveratrol concentrations. Fish Shellfish Immunol. 2018, 77, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Almeidaa, A.R.; Alvesb, M.; Dominguesa, I.; Henriques, I. The impact of antibiotic exposure in water and zebrafish gut microbiomes: A 16S rRNA gene-based metagenomic analysis. Ecotox. Environ. Safe. 2019, 186, 109771. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.Z.; Lv, L.X.; Wu, W.R.; Li, Y.T.; Shi, D.; Fang, D.Q.; Guo, F.F.; Jiang, H.Y.; Yan, R.; Ye, W.C.; et al. Butyrate protects mice against methionine-choline-deficient diet-induced nonalcoholic steatohepatitis by improving gut barrier function, attenuating inflammation and reducing endotoxin levels. Front. Microbiol. 2018, 9, 1967. [Google Scholar] [CrossRef] [Green Version]
- Geraylou, Z.; Souffreau, C.; Rurangwa, E.; Maes, G.E.; Spanier, K.I.; Courtin, C.M.; Delcour, J.A.; Buyse, J.; Ollevier, F. Prebiotic effects of arabinoxylan oligosaccharides on juvenile Siberian sturgeon (Acipenser baerii) with emphasis on the modulation of the gut microbiota using 454 pyrosequencing. FEMS Microbiol. Ecol. 2013, 86, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Yang, Y. Phylogenetic molecular ecological network of soil microbial communities in response to elevated CO2. MBio 2011, 2, e00122–11. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Jiang, Y.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [Green Version]
- Pande, S.; Kaftan, F.; Lang, S.; Svatoš, A.; Germerodt, S.; Kost, C. Privatization of cooperative benefits stabilizes mutualistic cross-feeding interactions in spatially structured environments. ISME J. 2016, 10, 1413. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | β-glucan Contents (%) | ||
|---|---|---|---|
| 0 | 0.20 | 0.40 | |
| Fish meal | 80.00 | 80.00 | 80.00 |
| Soybean meal | 450.00 | 450.00 | 450.00 |
| Maize meal | 250.00 | 250.00 | 250.00 |
| Wheat middlings | 74.00 | 74.00 | 74.00 |
| Maize oil | 36.00 | 36.00 | 36.00 |
| Dicalcium | 10.00 | 10.00 | 10.00 |
| Vitamin premix a | 5.00 | 5.00 | 5.00 |
| Mineral premix b | 5.00 | 5.00 | 5.00 |
| Cellulose | 60.00 | 57.73 | 55.46 |
| Carboxymethyl cellulose | 30.00 | 30.00 | 30.00 |
| β-glucan c | 0.00 | 2.27 | 4.54 |
| Total | 1000.00 | 1000.00 | 1000.00 |
| Gene | 5′-3′ Primer Sequence | GenBank Accession No. |
|---|---|---|
| IL-1β | F: GAGCACAGAATTCCAGGATGAAAG | XM_019365841.1 |
| R: TGAACTGAGGTGGTCCAGCTGT | ||
| MIF-2 | F: AGCAGAAGCAGGAAGGCGAAGA | XM_003444573.4 |
| R: CGGTACATCACCTCTGGCAACATT | ||
| TGF-β1 | F: AAGAGGAGGAGGAATACTTTGCCA | XM_003459454 |
| R: GAAGCTCATTGAGATGACTTTGGG | ||
| TNF-α | F: CAGAAGCACTAAAGGCGAAGAACA | NM_001279533 |
| R: TTCTAGATGGATGGCTGCCTTG | ||
| EF-1α | F: ATCAAGAAGATCGGCTACAACCCT | XM_005469373.3 |
| R: ATCCCTTGAACCAGCTCATCTTGT |
| Items | Freshwater | Brackish Water (16 psu)-β-glucan (%) | ||
|---|---|---|---|---|
| 0 | 0.2 | 0.4 | ||
| Initial weight | 1.27 ± 0.01 | 1.28 ± 0.01 | 1.28 ± 0.01 | 1.27 ± 0.01 |
| Final weight | 38.03 ± 2.86 | 41.50 ± 1.53 | 38.67 ± 2.29 | 38.40 ± 2.70 |
| Weight gain (%) | 2974.75 ± 226.02 | 3235.35 ± 109.63 | 3025.67 ± 172.76 | 3008.85 ± 206.30 |
| Relative spleen weight (%) | 0.29 ± 0.01 b | 0.37 ± 0.03 c | 0.24 ± 0.09 a | 0.23 ± 0.02 a |
| Survival (%) | 100 | 100 | 100 | 100 |
| Items | Freshwater | Brackish Water (16 psu)-β-glucan (%) | ||
|---|---|---|---|---|
| 0 | 0.2 | 0.4 | ||
| Richness estimate | ||||
| Chao 1 | 782.01 ± 138.12 ab | 939.67 ± 63.50 b | 758.00 ± 125.74 ab | 564.00 ± 16.07 a |
| ACE | 782.30 ± 138.15 ab | 939.67 ± 63.57 b | 758.11 ± 16.06 ab | 564.11 ± 16.06 a |
| Diversity estimators | ||||
| Shannon | 6.10 ± 0.13 ab | 7.02 ± 0.40 b | 6.01 ± 0.71 ab | 5.46 ± 0.11 a |
| Simpson | 0.94 ± 0.01 ab | 0.97 ± 0.01 b | 0.94 ± 0.02 ab | 0.93 ± 0.01 a |
| Items | Freshwater | 16 psu | 16 psu + 0.2% β-glucan | 16 psu + 0.4% β-glucan |
|---|---|---|---|---|
| Node | 50 | 50 | 50 | 50 |
| Edge | 402 | 406 | 450 | 518 |
| Average degree | 16.08 | 16.24 | 18 | 20.72 |
| Diameter | 1 | 1 | 1 | 1 |
| Graph density | 0.328 | 0.331 | 0.367 | 0.423 |
| Centralization | 0.041 | 0.080 | 0.128 | 0.155 |
| Heterogeneity | 0.153 | 0.198 | 0.347 | 0.426 |
| Average clustering Coefficient | 1 | 1 | 1 | 1 |
| Average path length | 1 | 1 | 1 | 1 |
| Positive/negative association (%) | 53.23/46.77 | 64.53/35.47 | 76/24 | 55.02/44.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Suo, Y.; Wang, X.; Qin, J.G.; Chen, L.; Li, E. Recovery from Hypersaline-Stress-Induced Immunity Damage and Intestinal-Microbiota Changes through Dietary β-glucan Supplementation in Nile tilapia (Oreochromis niloticus). Animals 2020, 10, 2243. https://doi.org/10.3390/ani10122243
Xu C, Suo Y, Wang X, Qin JG, Chen L, Li E. Recovery from Hypersaline-Stress-Induced Immunity Damage and Intestinal-Microbiota Changes through Dietary β-glucan Supplementation in Nile tilapia (Oreochromis niloticus). Animals. 2020; 10(12):2243. https://doi.org/10.3390/ani10122243
Chicago/Turabian StyleXu, Chang, Yantong Suo, Xiaodan Wang, Jian G Qin, Liqiao Chen, and Erchao Li. 2020. "Recovery from Hypersaline-Stress-Induced Immunity Damage and Intestinal-Microbiota Changes through Dietary β-glucan Supplementation in Nile tilapia (Oreochromis niloticus)" Animals 10, no. 12: 2243. https://doi.org/10.3390/ani10122243
APA StyleXu, C., Suo, Y., Wang, X., Qin, J. G., Chen, L., & Li, E. (2020). Recovery from Hypersaline-Stress-Induced Immunity Damage and Intestinal-Microbiota Changes through Dietary β-glucan Supplementation in Nile tilapia (Oreochromis niloticus). Animals, 10(12), 2243. https://doi.org/10.3390/ani10122243