Long-Term Determinants of the Seroprevalence of Toxoplasma gondii in a Wild Ungulate Community

 ,
,  , ,
, ,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
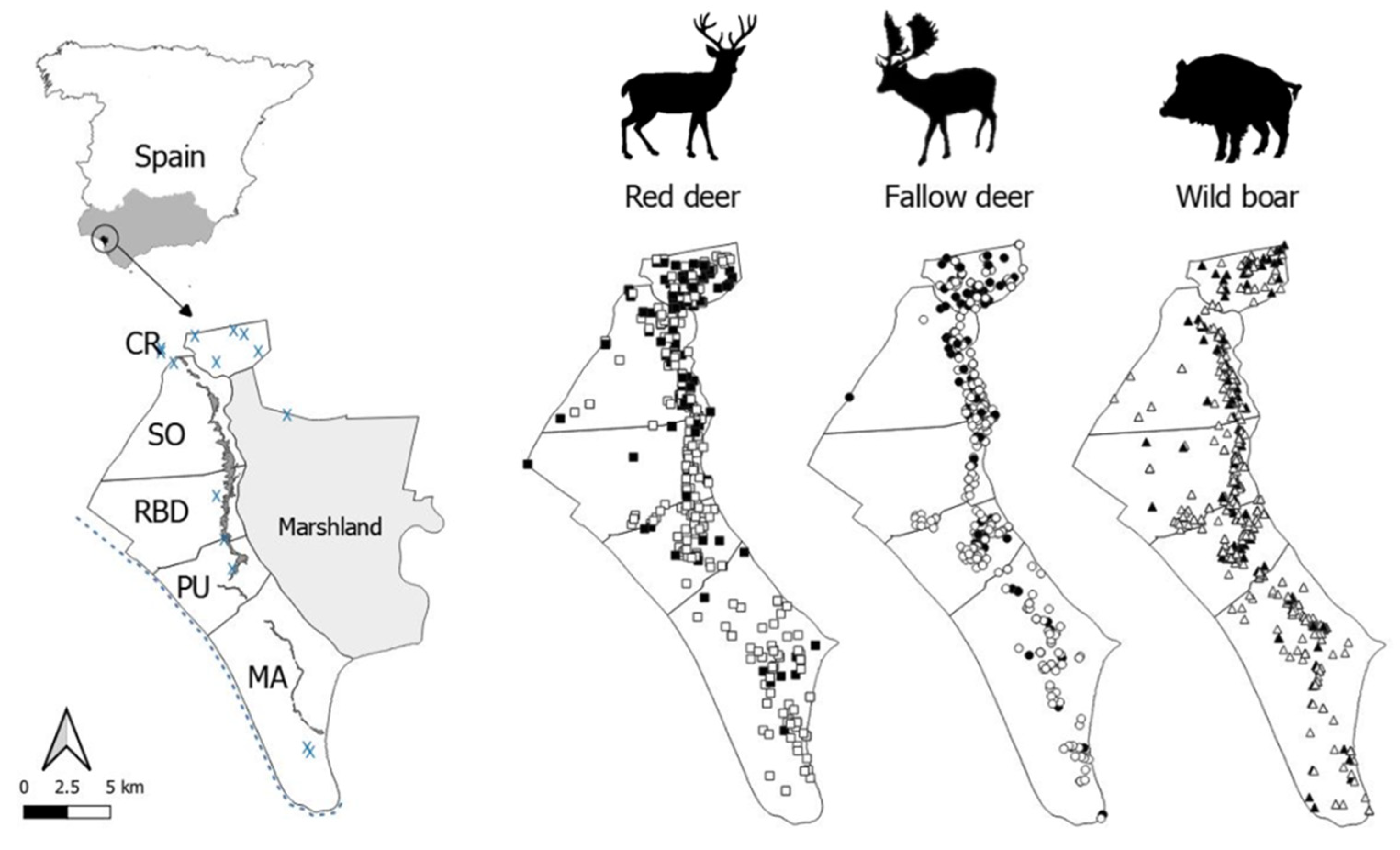
2.1. Study Area
2.2. Animal Sampling
2.3. Serological Testing
2.4. Data Collection
2.4.1. Individual Factors
2.4.2. Environmental Factors
2.4.3. Population Factors
2.4.4. Stochastic Factors
2.5. Risk Factor Analysis
3. Results
3.1. General
3.2. Factors Determining the Seroprevalence of T. gondii
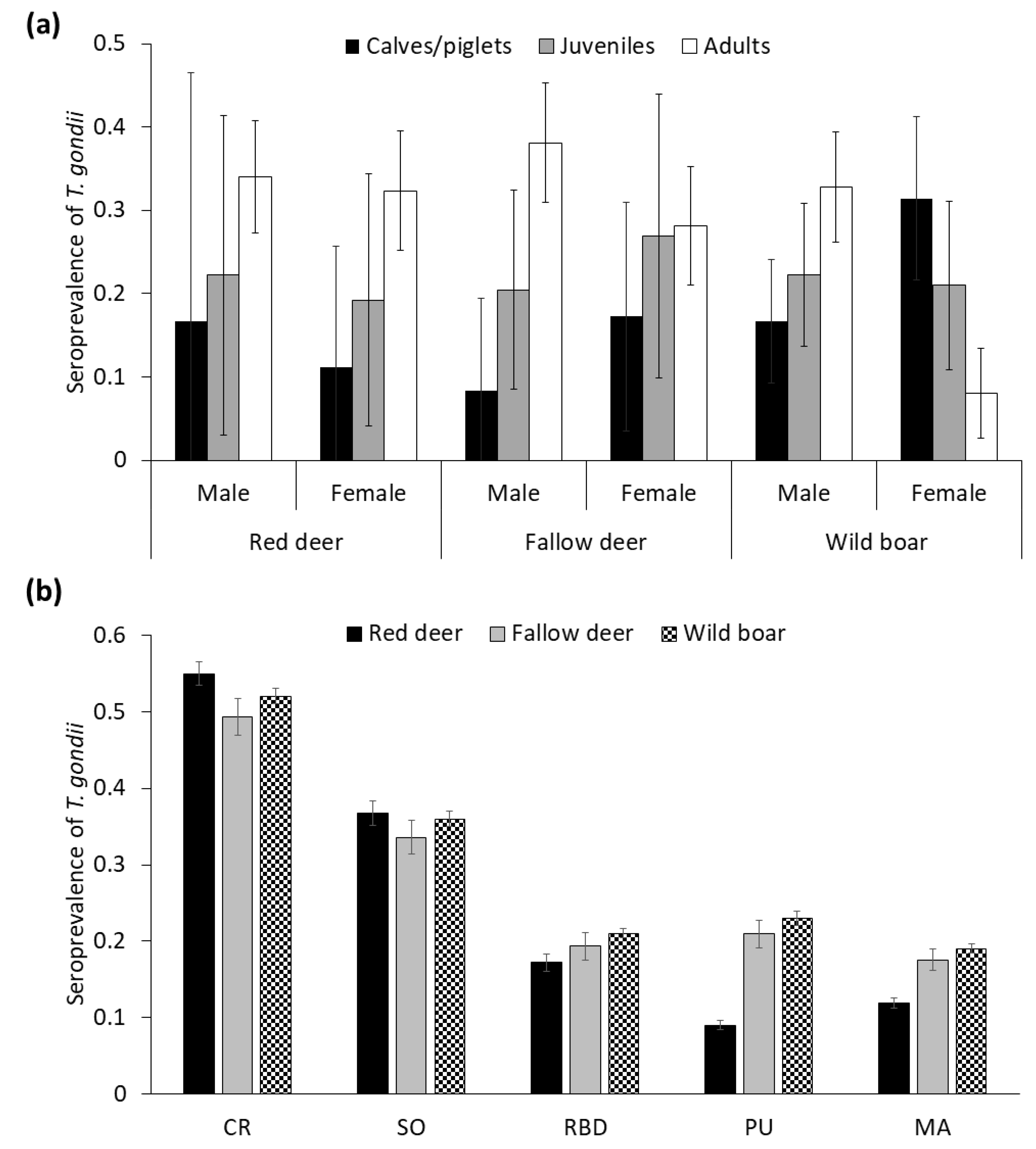
3.2.1. Individual Factors
3.2.2. Environmental Factors
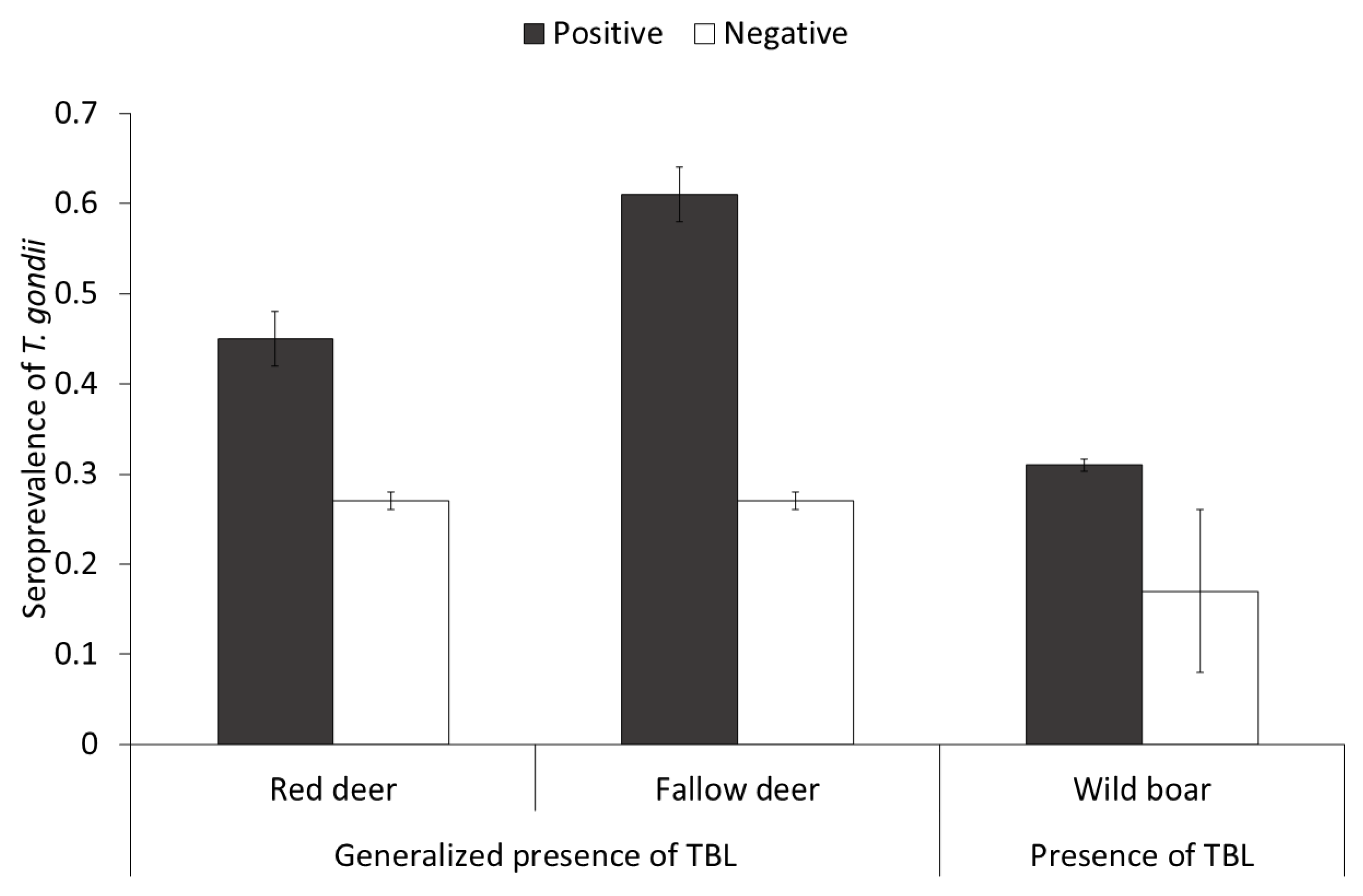
3.2.3. Population Factors
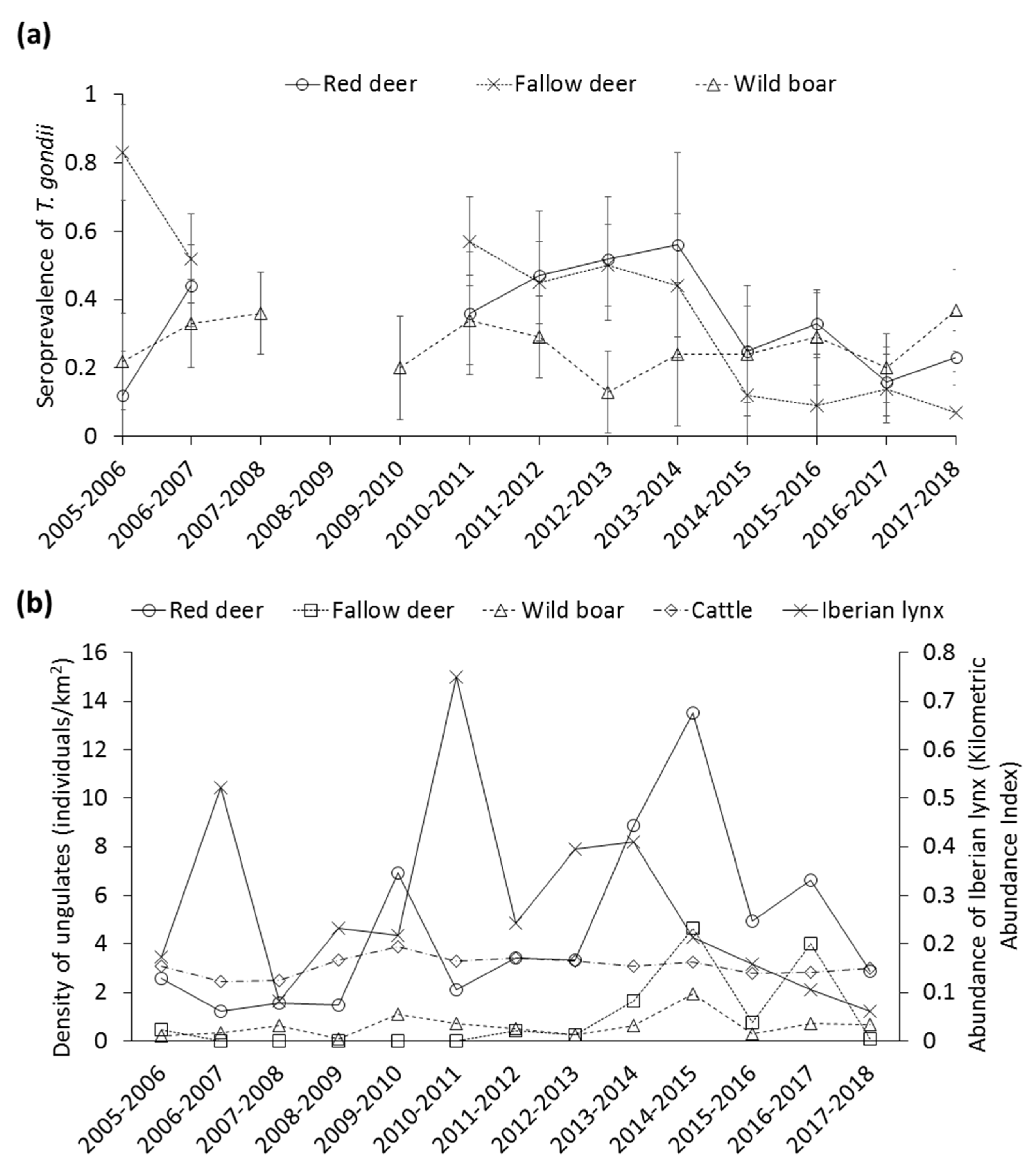
3.2.4. Stochastic Factors
4. Discussion
4.1. General Patterns of the Seroprevalence of T. gondii
4.2. Individual Factors
4.3. Environmental Factors
4.4. Population Factors
4.5. Stochastic Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Hill, D.E.; Dubey, J.P. Toxoplasmosis. US Geol. Surv. Circ. 2014, 1389, 84. [Google Scholar] [CrossRef]
- Gauss, C.; Dubey, J.; Vidal, D.; Fons, F.R.; Vicente, J.; Marco, I.; Lavín, S.; Gortazar, C.; Almeria, S.; Vidal, D. Seroprevalence of Toxoplasma gondii in wild pigs (Sus scrofa) from Spain. Vet. Parasitol. 2005, 131, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Panadero, R.; Painceira, A.; López, C.; Vázquez, L.; Paz, A.; Díaz, P.; DaCal, V.; Cienfuegos, S.; Fernández, G.; Lago, N.; et al. Seroprevalence of Toxoplasma gondii and Neospora caninum in wild and domestic ruminants sharing pastures in Galicia (Northwest Spain). Res. Vet. Sci. 2010, 88, 111–115. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Cabezón, O.; Hernández, E.; Martínez-Cruz, M.S.; Martínez-Moreno, Á.; Martínez-Moreno, J. Toxoplasma gondii in Ruminant Species (Cattle, Sheep, and Goats) from Southern Spain. J. Parasitol. 2013, 99, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Calero-Bernal, R.; Pérez-Martín, J.E.; Reina, D.; Serrano, F.J.; Frontera, E.; Fuentes, I.; Dubey, J.P. Detection of Zoonotic Protozoa Toxoplasma gondii and Sarcocystis suihominis in Wild Boars from Spain. Zoonoses Public Health 2016, 63, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Cuenca, J.C.; Díaz-Cao, J.M.; Martínez-Moreno, Á.; Cano-Terriza, D.; Jiménez-Ruiz, S.; Almería, S.; García-Bocanegra, I. Seroepidemiology of Toxoplasma gondii in extensively raised Iberian pigs in Spain. Prev. Vet. Med. 2020, 175, 104854. [Google Scholar] [CrossRef] [PubMed]
- Almeria, S.; Cabezón, O.; Paniagua, J.; Cano-Terriza, D.; Jiménez-Ruiz, S.; Arenas-Montes, A.; Dubey, J.P.; García-Bocanegra, I. Toxoplasma gondii in sympatric domestic and wild ungulates in the Mediterranean ecosystem. Parasitol. Res. 2018, 117, 665–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Martín, D.; García-Bocanegra, I.; Almería, S.; Castro-Scholten, S.; Dubey, J.P.; Amaro-López, M.A.; Cano-Terriza, D. Epidemiological surveillance of Toxoplasma gondii in small ruminants in southern Spain. Prev. Vet. Med. 2020, 183. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Simon-Grifé, M.; Dubey, J.P.; Casal, J.; Martín, G.E.; Cabezón, O.; Perea, A.; Almería, S. Seroprevalence and risk factors associated with Toxoplasma gondii in domestic pigs from Spain. Parasitol. Int. 2010, 59, 421–426. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Dubey, J.; Simon-Grifé, M.; Cabezón, O.; Casal, J.; Allepuz, A.; Napp, S.; Almeria, S. Seroprevalence and risk factors associated with Toxoplasma gondii infection in pig farms from Catalonia, north-eastern Spain. Res. Vet. Sci. 2010, 89, 85–87. [Google Scholar] [CrossRef]
- Sobrino, R.; Cabezón, O.; Millán, J.; Pabón, M.; Arnal, M.; Luco, D.; Gortazar, C.; Dubey, J.P.; Almeria, S. Seroprevalence of Toxoplasma gondii antibodies in wild carnivores from Spain. Vet. Parasitol. 2007, 148, 187–192. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.; Robertson, A.; Silk, M.J. Wildlife disease ecology from the individual to the population: Insights from a long-term study of a naturally infected European badger population. J. Anim. Ecol. 2017, 87, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Barroso, P.; Barasona, J.Á.; Acevedo, P.; Palencia, P.; Carro, F.; Negro, J.J.; Torres, M.J.; Gortazar, C.; Soriguer, R.C.; Vicente, J. Long-Term Determinants of Tuberculosis in the Ungulate Host Community of Doñana National Park. Pathogens 2020, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Barasona, J.A.; Acevedo, P.; Ruiz-Fons, J.F.; Boadella, M.; Diez-Delgado, I.; Beltran-Beck, B.; González-Barrio, D.; Queirós, J.; Montoro, V.; et al. Temporal Trend of Tuberculosis in Wild Ungulates from Mediterranean Spain. Transbound. Emerg. Dis. 2013, 60, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Barroso, P.; Acevedo, P.; Vicente, J. The importance of long-term studies on wildlife diseases and their interfaces with humans and domestic animals: A review. Transbound. Emerg. Dis. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Life + IBERLINCE. Life + IBERLINCE, Recuperación de la Distribución Histórica del Lince Ibérico (Lynx pardinus) en España y Portugal. (LIFE10NAT/ES/570). Available online: http://www.iberlince.eu/index.php/esp/lince-iberico-esp/ (accessed on 18 June 2020).
- García-Bocanegra, I.; Dubey, J.; Martinez, F.; Vargas, A.; Cabezón, O.; Zorrilla, I.; Arenas, A.; Almeria, S. Factors affecting seroprevalence of Toxoplasma gondii in the endangered Iberian lynx (Lynx pardinus). Vet. Parasitol. 2010, 167, 36–42. [Google Scholar] [CrossRef]
- Millán, J.; Candela, M.G.; Palomares, F.; Cubero, M.J.; Rodriguez, A.W.; Barral, M.; De La Fuente, J.; Almeria, S.; León-Vizcaíno, L. Disease threats to the endangered Iberian lynx (Lynx pardinus). Vet. J. 2009, 182, 114–124. [Google Scholar] [CrossRef]
- Fernández-Delgado, C. Conservation management of a European natural area: Doñana National Park, Spain. In Principles of Conservation Biology, 2nd ed.; Meffe, G.K., Carroll, C.R., Eds.; Sinauer Associates: Sunderland, MA, USA, 1997; pp. 458–467. [Google Scholar]
- Barasona, J.Á.; Latham, M.C.; Acevedo, P.; Armenteros, J.A.; Latham, A.D.M.; Gortazar, C.; Carro, F.; Soriguer, R.C.; Vicente, J. Spatiotemporal interactions between wild boar and cattle: Implications for cross-species disease transmission. Vet. Res. 2014, 45, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Delgado, R.; Aragonés, D.; Afán, I.; Bustamante, J. Long-Term Monitoring of the Flooding Regime and Hydroperiod of Doñana Marshes with Landsat Time Series (1974–2014). Remote. Sens. 2016, 8, 775. [Google Scholar] [CrossRef] [Green Version]
- Valverde, J.A. Estructura de una Comunidad de Vertebrados Terrestres; Monografías de la Estación Biológica de Doñana; CSIC: Madrid, Spain, 1967.
- Junta de Andalucia. Memoria de Actividades y Resultados del Espacio Natural de Doñana; Consejeria de Medio Ambiente y Ordenación del Territorio: Andalucia, Spain, 2013.
- EEC. Council Directive 86/609/EEC of 24 November 1986 on the approximation of laws, regulations and administrative provisions of the Member States regarding the protection of animals used for experimental and other scientific purposes. Off. J. Eur. Communities 1986, 358, 1–29. [Google Scholar]
- MAPA. Real Decreto 223/1988, de 14 de Marzo, Sobre Protección de los Animales Utilizados Para Experimentación y otros Fines Científicos; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1988; pp. 8509–8512.
- Buchanan, K.; Burt de Perera, T.; Carere, C.; Carter, T.; Hailey, A.; Hubrecht, R.; Jennings, D.; Metcalfe, N.; Pitcher, T.; Peron, F.; et al. ASAB Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 2012, 83, 301–309. [Google Scholar]
- Jiménez-Ruiz, S.; Arenas-Montes, A.; Cano-Terriza, D.; Paniagua, J.; Pujols, J.; Miró, F.; Fernández-Aguilar, X.; González, M.Á.; Franco, J.J.; García-Bocanegra, I. Blood extraction method by endocranial venous sinuses puncture in hunted wild ruminants. Eur. J. Wildl. Res. 2016, 62, 775–780. [Google Scholar] [CrossRef]
- Dubey, J.P.; Desmonts, G. Serological responses of equids fed Toxoplasma gondii oocysts. Equine Vet. J. 1987, 19, 337–339. [Google Scholar] [CrossRef]
- Dubey, J.P.; Cerqueira-Cézar, C.K.; Murata, F.H.A.; Verma, S.K.; Kwok, O.C.H.; Pedersen, K.; Rosenthal, B.M.; Su, C. White-tailed deer (Odocoileus virginianus) are a reservoir of a diversity of Toxoplasma gondii strains in the USA and pose a risk to consumers of undercooked venison. Parasitology 2020, 147, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Murata, F.; Cezar, C.; Kwok, O. Epidemiologic and public health significance of Toxoplasma gondii infections in venison: 2009–2020. J. Parasitol. 2021, in press. [Google Scholar]
- Dubey, J.P.; Cerqueira-Cézar, C.K.; Murata, F.H.A.; Verma, S.K.; Kwok, O.C.H.; Pedersen, K.; Rosenthal, B.M.; Su, C. Genotyping of viable Toxoplasma gondii from the first national survey of feral swine revealed evidence for sylvatic transmission cycle, and presence of highly virulent parasite genotypes. Parasitology 2019, 147, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Cerqueira-Cézar, C.K.; Murata, F.H.; Kwok, O.C.; Hill, D.; Yang, Y.R.; Su, C. WITHDRAWN: All about Toxoplasma gondii infections in pigs: The past decade. Vet. Parasitol. 2020, 288, 109182. [Google Scholar] [CrossRef]
- Gauss, C.; Dubey, J.P.; Vidal, D.; Cabezón, O.; Ruiz-Fons, F.; Vicente, J.; Marco, I.; Lavin, S.; Gortazar, C.; Almería, S. Prevalence of Toxoplasma gondii antibodies in red deer (Cervus elaphus) and other wild ruminants from Spain. Vet. Parasitol. 2006, 136, 193–200. [Google Scholar] [CrossRef]
- Gamarra, J.; Cabezón, O.; Pabón, M.; Arnal, M.; Luco, D.; Dubey, J.; Gortázar, C.; Almeria, S. Prevalence of antibodies against Toxoplasma gondii in roe deer from Spain. Vet. Parasitol. 2008, 153, 152–156. [Google Scholar] [CrossRef]
- Saenz de Buruaga, M.; Lucio-Calero, A.; Purroy, F.J. Reconocimiento de Sexo y Edad en Especies Cinegéticas, 1st ed.; Edilesa: Madrid, Spain, 2001; ISBN 9788480123716. [Google Scholar]
- Byrne, A.; Graham, J.; McConville, J.; Milne, G.; Guelbenzu-Gonzalo, M.; McDowell, S. Liver fluke (Fasciola hepatica) co-infection with bovine tuberculosis in cattle: A prospective herd-level assessment of herd bTB risk in dairy enterprises. Transbound. Emerg. Dis. 2019, 66, 1727–1736. [Google Scholar] [CrossRef]
- Vicente, J.; Höfle, U.; Garrido, J.; Fernández-De-Mera, I.G.; Juste, R.; Barral, M.; Gortazar, C. Wild boar and red deer display high prevalences of tuberculosis-like lesions in Spain. Vet. Res. 2006, 37, 107–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braza, F.; Alvarez, F. Habitat use by Red Deer and Fallow Deer in Doñana National Park. Misc. Zool. 1987, 11, 363–367. [Google Scholar]
- Barasona, J.Á.; Mulero-Pázmány, M.; Acevedo, P.; Negro, J.J.; Torres, M.J.; Gortázar, C.; Vicente, J. Unmanned Aircraft Systems for Studying Spatial Abundance of Ungulates: Relevance to Spatial Epidemiology. PLOS ONE 2014, 9, e115608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- QGIS Development Team. Quantum GIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: https://qgis.org/es/site/ (accessed on 16 May 2020).
- REDIAM Consejería de Medio Ambiente y Ordenación del Territorio. Andalucía, España: Red de Información Ambiental de Andalucía, REDIAM. Available online: http://www.juntadeandalucia.es/medioambiente/site/rediam (accessed on 20 September 2007).
- Miller, M.; Gardner, I.; Kreuder, C.; Paradies, D.; Worcester, K.; Jessup, D.; Dodd, E.; Harris, M.; Ames, J.; Packham, A.; et al. Coastal freshwater runoff is a risk factor for Toxoplasma gondii infection of southern sea otters (Enhydra lutris nereis). Int. J. Parasitol. 2002, 32, 997–1006. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasmosis—A waterborne zoonosis. Vet. Parasitol. 2004, 126, 57–72. [Google Scholar] [CrossRef]
- Villari, S.; Vesco, G.; Petersen, E.; Crispo, A.; Buffolano, W. Risk factors for toxoplasmosis in pigs bred in Sicily, Southern Italy. Vet. Parasitol. 2009, 161, 1–8. [Google Scholar] [CrossRef]
- Cabal, A.; Vicente, J.; Alvarez, J.; Barasona, J.A.; Boadella, M.; Dominguez, L.; Gortazar, C. Human influence and biotic homogenization drive the distribution of Escherichia coli virulence genes in natural habitats. Microbiologyopen 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Buckland, S.; Anderson, D.; Burnham, K.; Laake, J.; Borchers, D.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Acevedo, P.; Fons, F.R.; Vicente, J.; Reyes-García, A.R.; Alzaga, V.; Gortazar, C. Estimating red deer abundance in a wide range of management situations in Mediterranean habitats. J. Zool. 2008, 276, 37–47. [Google Scholar] [CrossRef]
- Thomas, L.; Buckland, S.T.; Rexstad, E.A.; Laake, J.L.; Strindberg, S.; Hedley, S.L.; Bishop, J.R.B.; Marques, T.A.; Burnham, K.P. Distance software: Design and analysis of distance sampling surveys for estimating population size. J. Appl. Ecol. 2010, 47, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- EBD-CSIC Equipo de Seguimiento del Espacio Nacional de Doñana. Available online: http://icts.ebd.csic.es/datos-meteorologicos (accessed on 5 July 2019).
- Dubey, J.P.; Beattie, C.P. Toxoplasmosis of Animals and Man; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. GLM and GAM for Count Data. In Environmental Forum; Springer: New York, NY, USA, 2009; Volume 21, pp. 209–243. ISBN 9780387874579. [Google Scholar]
- Göloglu, F.; Pott, A. Results on the Crosscorrelation and Autocorrelation of Sequences. In Proceedings of the Sequences and Their Applications-SETA 2008 5th International Conference, Lexington, KY, USA, 14–18 September 2008; Springer: Berlin/Heidelberg, Germany, 2008; pp. 95–105. [Google Scholar]
- R Core Team. R Core Team R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Jokelainen, P.; Näreaho, A.; Hälli, O.; Heinonen, M.; Sukura, A. Farmed wild boars exposed to Toxoplasma gondii and Trichinella spp. Vet. Parasitol. 2012, 187, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Reiterová, K.; Špilovská, S.; Blaňarová, L.; Derdáková, M.; Čobádiová, A.; Hisira, V. Wild boar (Sus scrofa)—Reservoir host of Toxoplasma gondii, Neospora caninum and Anaplasma phagocytophilum in Slovakia. Acta Parasitol. 2016, 61, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Fons, F.R.; Vicente, J.; Vidal, D.; Höfle, U.; Villanúa, D.; Gauss, C.; Segalés, J.; Almeria, S.; Montoro, V.; Gortazar, C.; et al. Seroprevalence of six reproductive pathogens in European wild boar (Sus scrofa) from Spain: The effect on wild boar female reproductive performance. Theriogenology 2006, 65, 731–743. [Google Scholar] [CrossRef] [Green Version]
- Coelho, C.; Vieira-Pinto, M.; Faria, A.S.; Vale-Gonçalves, H.M.; Veloso, O.; Paiva-Cardoso, M.D.N.; Mesquita, J.R.; Lopes, A.P. Serological evidence of Toxoplasma gondii in hunted wild boar from Portugal. Vet. Parasitol. 2014, 202, 310–312. [Google Scholar] [CrossRef]
- Beral, M.; Rossi, S.; Aubert, D.; Gasqui, P.; Terrier, M.-E.; Klein, F.; Villena, I.; Abrial, D.; Gilot-Fromont, E.; Richomme, C.; et al. Environmental Factors Associated with the Seroprevalence of Toxoplasma gondii in Wild Boars (Sus scrofa), France. EcoHealth 2012, 9, 303–309. [Google Scholar] [CrossRef]
- Rocchigiani, G.; Nardoni, S.; D’Ascenzi, C.; Nicoloso, S.; Picciolli, F.; Papini, R.; Mancianti, F. Seroprevalence of Toxoplasma gondii and Neospora caninum in red deer from Central Italy. Ann. Agric. Environ. Med. 2016, 23, 699–701. [Google Scholar] [CrossRef] [Green Version]
- Bártová, E.; Sedlák, K.; Pavlik, I.; Literak, I. Prevalence of Neospora caninum and Toxoplasma gondii Antibodies in Wild Ruminants from the Countryside or Captivity in the Czech Republic. J. Parasitol. 2007, 93, 1216–1218. [Google Scholar] [CrossRef]
- Calero-Bernal, R.; Saugar, J.M.; Frontera, E.; Pérez-Martín, J.E.; Habela, M.A.; Serrano, F.J.; Reina, D.; Fuentes, I. Prevalence and genotype identification of Toxoplasma gondii in wild animals from southwestern Spain. J. Wildl. Dis. 2015, 51, 233–238. [Google Scholar] [CrossRef]
- Wilson, A.G.; Lapen, D.R.; Mitchell, G.W.; Provencher, J.F.; Wilson, S. Interaction of diet and habitat predicts Toxoplasma gondii infection rates in wild birds at a global scale. Glob. Ecol. Biogeogr. 2020, 29, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Opsteegh, M.; Swart, A.; Fonville, M.; Dekkers, L.; Van Der Giessen, J. Age-Related Toxoplasma gondii Seroprevalence in Dutch Wild Boar Inconsistent with Lifelong Persistence of Antibodies. PLOS ONE 2011, 6, e16240. [Google Scholar] [CrossRef] [PubMed]
- Gortazar, C.; Torres, M.J.; Vicente, J.; Acevedo, P.; Reglero, M.; De La Fuente, J.; Negro, J.J.; Aznar-Martín, J. Bovine Tuberculosis in Doñana Biosphere Reserve: The Role of Wild Ungulates as Disease Reservoirs in the Last Iberian Lynx Strongholds. PLOS ONE 2008, 3, e2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jokelainen, P.; Näreaho, A.; Knaapi, S.; Oksanen, A.; Rikula, U.; Sukura, A. Toxoplasma gondii in wild cervids and sheep in Finland: North-south gradient in seroprevalence. Vet. Parasitol. 2010, 171, 331–336. [Google Scholar] [CrossRef]
- Garrote, G.; De Ayala, R.P.; Pereira, P.; Robles, F.; Guzman, N.; García, F.J.; Iglesias, M.C.; Hervás, J.; Fajardo, I.; Simón, M.; et al. Estimation of the Iberian lynx (Lynx pardinus) population in the Doñana area, SW Spain, using capture–recapture analysis of camera-trapping data. Eur. J. Wildl. Res. 2011, 57, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Millán, J.; Casanova, J.C.; Millán, J.; Casanova, J.C. Helminth parasites of the endangered Iberian lynx (Lynx pardinus) and sympatric carnivores. J. Helminthol. 2007, 81, 377–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente, J.; Palomares, F.; De Ibañez, A.R.; Ortiz, J.; De Ibañez, R.R. Epidemiology of Ancylostoma spp. in the endangered Iberian lynx (Lynx pardinus) in the Doñana National Park, south-west Spain. J. Helminthol. 2004, 78, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P. The history and life cycle of Toxoplasma gondii. In Toxoplasma Gondii; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–19. [Google Scholar]
- Antolová, D.; Reiterová, K.; Dubinský, P. Seroprevalence of Toxoplasma gondii in wild boars (Sus scrofa) in the Slovak Republic. Ann. Agric. Environ. Med. 2007, 14, 71–73. [Google Scholar]
- Mashaly, M.; Nabih, N.; Fawzy, I.M.; El Henawy, A.A. Tuberculosis/toxoplasmosis co-infection in Egyptian patients: A reciprocal impact. Asian Pac. J. Trop. Med. 2017, 10, 315–319. [Google Scholar] [CrossRef]
- Hwang, E.H.; Ahn, P.G.; Lee, D.M.; Kim, H.S. Cerebral Toxoplasmosis Combined with Disseminated Tuberculosis. J. Korean Neurosurg. Soc. 2012, 51, 316–319. [Google Scholar] [CrossRef]
- Delibes, M. Feeding ecology of the Spanish lynx in the Coto Doñana. Acta Theriol. (Warsz). 1980, 25, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-García, R.; Barroso, P.; Perez-Olivares, J.; Montoro, V.; Vicente, J. Consumption of Big Game Remains by Scavengers: A Potential Risk as Regards Disease Transmission in Central Spain. Front. Vet. Sci. 2018, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schley, L.; Roper, T.J. Diet of wild boar Sus scrofa in Western Europe, with particular reference to consumption of agricultural crops. Mammal Rev. 2003, 33, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Lochmiller, R.L.; Deerenberg, C. Trade-offs in evolutionary immunology: Just what is the cost of immunity? Oikos 2000, 88, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Malo, A.F.; Roldan, E.; Garde, J.; Soler, A.; Vicente, J.; Gortazar, C.; Gomendio, M. What does testosterone do for red deer males? Proc. R. Soc. B Boil. Sci. 2008, 276, 971–980. [Google Scholar] [CrossRef]
- Mauget, R.; Campan, R.; Spitz, F.; Dardaillom, M.; Janeau, G.; Pepin, D. Synthise des connaissances actuelles sur la biologie du sanglier, perspectives de recherche. In Proceedings of the Symposium International Sanglier, Les colloques de l’INRA, Tolouse, France, 24–26 April 1984. [Google Scholar]
- Rougier, S.; Montoya, J.G.; Peyron, F. Lifelong Persistence of Toxoplasma Cysts: A Questionable Dogma? Trends Parasitol. 2017, 33, 93–101. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Red Deer | Fallow Deer | Wild Boar | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F df (x,y) | Estimate ± SD | p | F df (x,y) | Estimate ± SD | p | F df (x,y) | Estimate ± SD | p | |
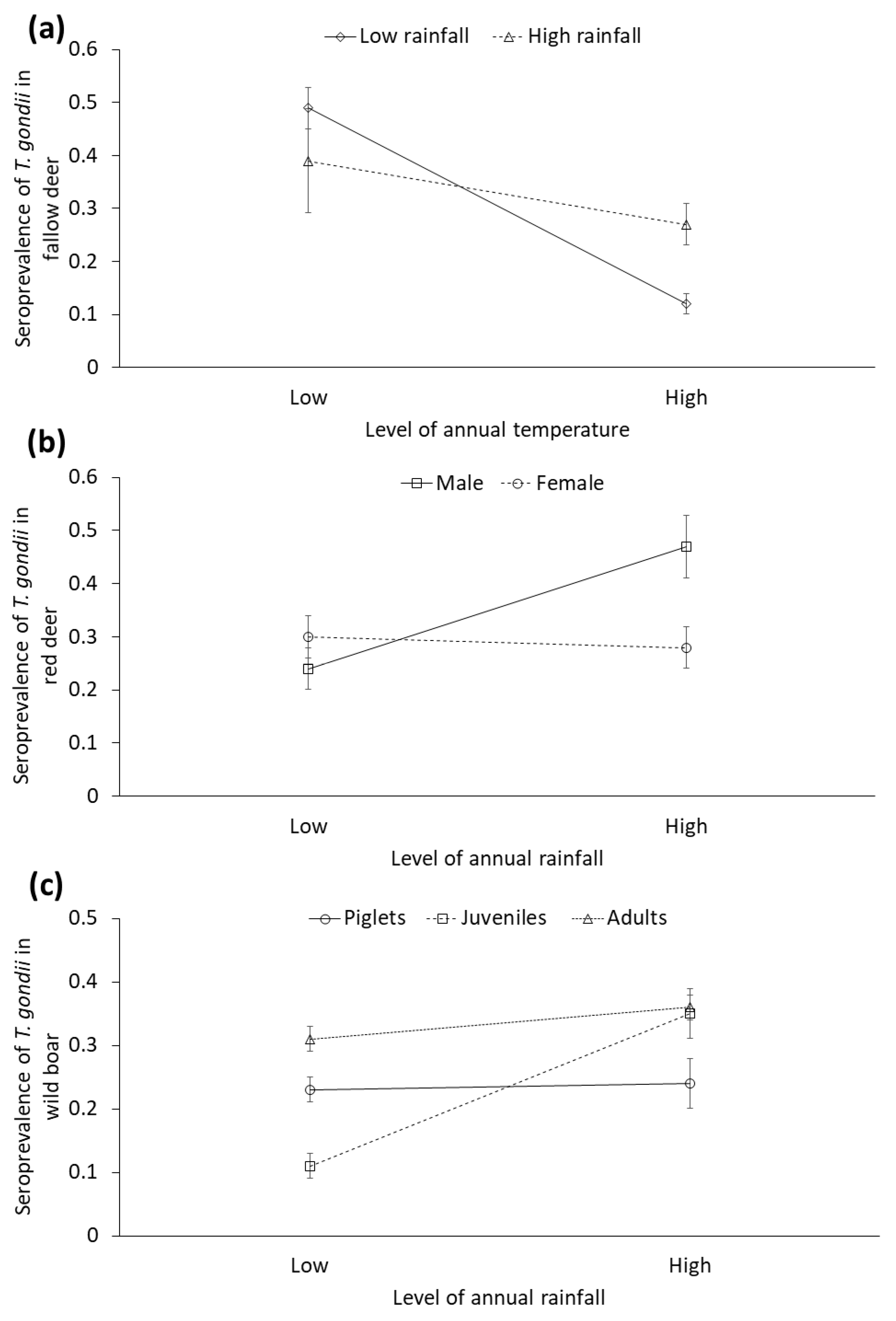
| Sex 1 | 0.44 (1, 420) | Female: −3.01 ± 0.99 | <0.01 | 0.22 (1, 695) | Female: 0.89 ± 0.38 | 0.03 | |||
| Age 2 | 1.46 (2, 420) | Juveniles: 0.01 ± 0.81 Adults: 1.03 ± 0.70 | 0.04 | 2.86 (2, 695) | Juveniles: −3.76 ± 1.24 Adults: 0.85 ± 0.76 | <0.01 | |||
| Presence of generalized TBL 3 | 2.95 (1, 420) | Positive: 0.59 ± 0.31 | 0.05 | 14.47 (1, 446) | Positive: 1.86 ± 0.48 | <0.01 | |||
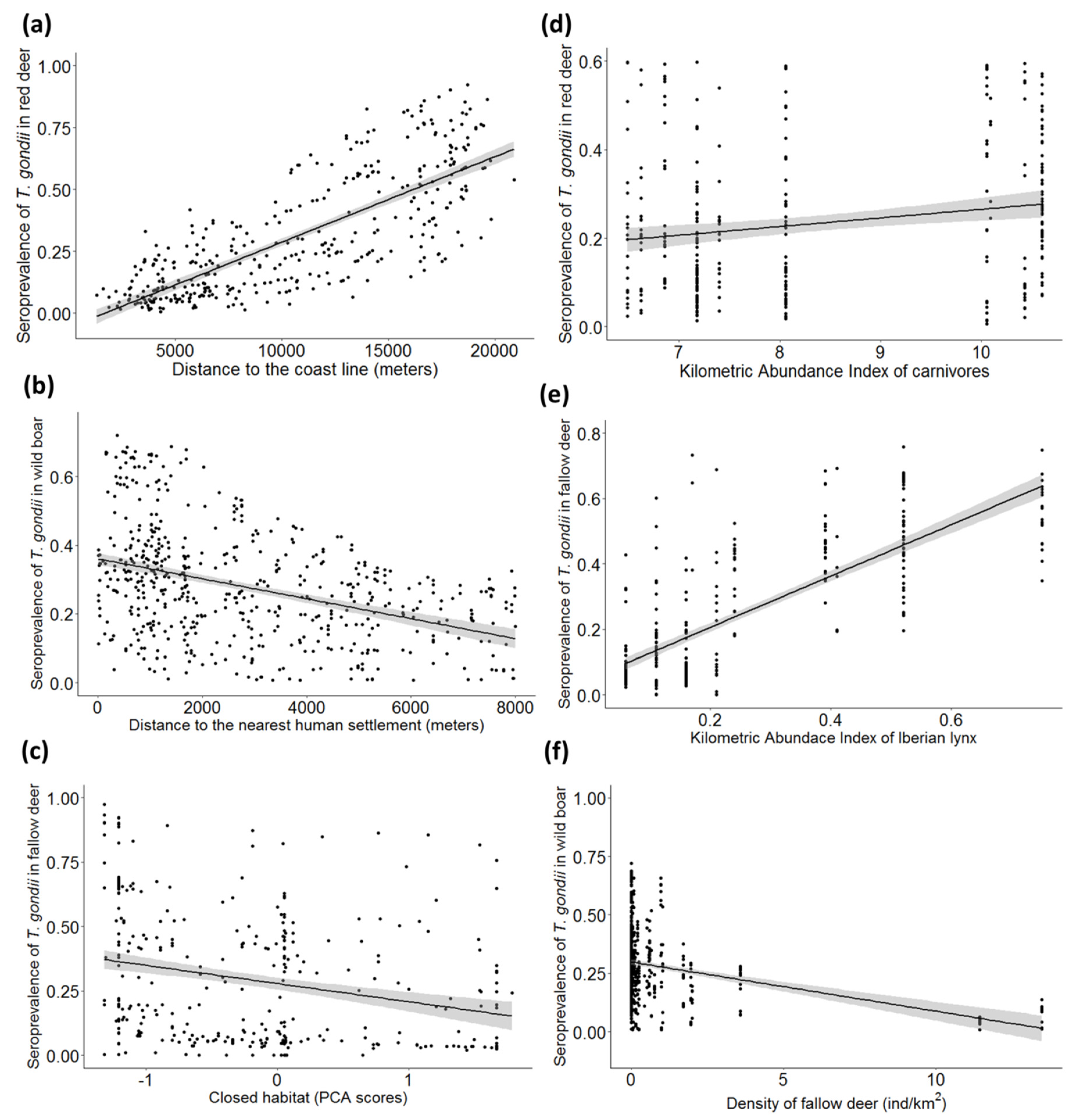
| DCOAST | 25.19 (1, 420) | 0.0002 ± 0.00002 | <0.01 | 5.69 (1, 446) | 0.00007 ± 0.00003 | 0.01 | 26.70 (1, 695) | 0.0001 ± 0.00002 | <0.01 |
| DE | 17.67 (1, 420) | 0.0002 ± 0.00009 | 0.07 | ||||||
| DHS | 2.93 (1, 695) | −0.0001 ± 0.00005 | <0.01 | ||||||
| Closed habitat | 5.75 (1, 446) | −0.31 ± 0.15 | 0.04 | ||||||
| Previous season´s rainfall | 3.24 (1, 420) | 0.01 ± 0.001 | <0.01 | 0.07 (1, 446) | −0.05 ± 0.01 | <0.01 | 2.40 (1, 695) | −0.0006 ± 0.001 | 0.15 |
| Previous season´s temperature | 3.23 (1, 420) | −0.68 ± 0.32 | 0.04 | 3.88 (1, 446) | −1.61 ± 0.57 | <0.01 | |||
| Red deer density | 0.43 (1, 420) | −0.11 ± 0.04 | <0.01 | 1.27 (1, 695) | −0.05 ± 0.03 | 0.07 | |||
| Fallow deer density | 0.43 (1, 420) | 0.11 ± 0.06 | 0.04 | 6.03 (1, 446) | −0.42 ± 0.18 | 0.02 | 7.07 (1, 695) | −0.16 ± 0.07 | 0.02 |
| Wild boar density | 4.1 (1, 695) | 0.34 ± 0.16 | 0.04 | ||||||
| Horse density | 1.14 (1, 420) | 0.08 ± 0.05 | 0.10 | ||||||
| Iberian lynx abundance | 5.81 (1, 446) | 3.56 ± 1.60 | 0.03 | 0 (1, 695) | −0.86 ± 0.45 | 0.06 | |||
| Carnivores abundance | 13.88 (1, 420) | 0.35 ± 0.09 | <0.01 | ||||||
| Sex * Rainfall | 10.18 (1, 420) | Rainfall * Female: −0.01 ± 0.002 | <0.01 | ||||||
| Sex * Age | 2.34 (2, 695) | Female * juveniles: −0.97 ± 0.58 Female * ≥ adults: −1.01 ± 0.45 | 0.07 | ||||||
| Temperature * Rainfall | 1.95 (1, 446) | 0.003 ± 0.0005 | <0.01 | ||||||
| Rainfall * Age 2 | 7.19 (2, 695) | Rainfall * juveniles: 0.01 ± 0.002 Rainfall * adults: 0.0003 ± 0.001 | <0.01 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barroso, P.; García-Bocanegra, I.; Acevedo, P.; Palencia, P.; Carro, F.; Jiménez-Ruiz, S.; Almería, S.; Dubey, J.P.; Cano-Terriza, D.; Vicente, J. Long-Term Determinants of the Seroprevalence of Toxoplasma gondii in a Wild Ungulate Community. Animals 2020, 10, 2349. https://doi.org/10.3390/ani10122349
Barroso P, García-Bocanegra I, Acevedo P, Palencia P, Carro F, Jiménez-Ruiz S, Almería S, Dubey JP, Cano-Terriza D, Vicente J. Long-Term Determinants of the Seroprevalence of Toxoplasma gondii in a Wild Ungulate Community. Animals. 2020; 10(12):2349. https://doi.org/10.3390/ani10122349
Chicago/Turabian StyleBarroso, Patricia, Ignacio García-Bocanegra, Pelayo Acevedo, Pablo Palencia, Francisco Carro, Saúl Jiménez-Ruiz, Sonia Almería, Jitender P. Dubey, David Cano-Terriza, and Joaquín Vicente. 2020. "Long-Term Determinants of the Seroprevalence of Toxoplasma gondii in a Wild Ungulate Community" Animals 10, no. 12: 2349. https://doi.org/10.3390/ani10122349
APA StyleBarroso, P., García-Bocanegra, I., Acevedo, P., Palencia, P., Carro, F., Jiménez-Ruiz, S., Almería, S., Dubey, J. P., Cano-Terriza, D., & Vicente, J. (2020). Long-Term Determinants of the Seroprevalence of Toxoplasma gondii in a Wild Ungulate Community. Animals, 10(12), 2349. https://doi.org/10.3390/ani10122349