Morphological Correlates of Personality in Female Asian Particolored Bats (Vespertilio sinensis)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Bat Capture and Husbandry
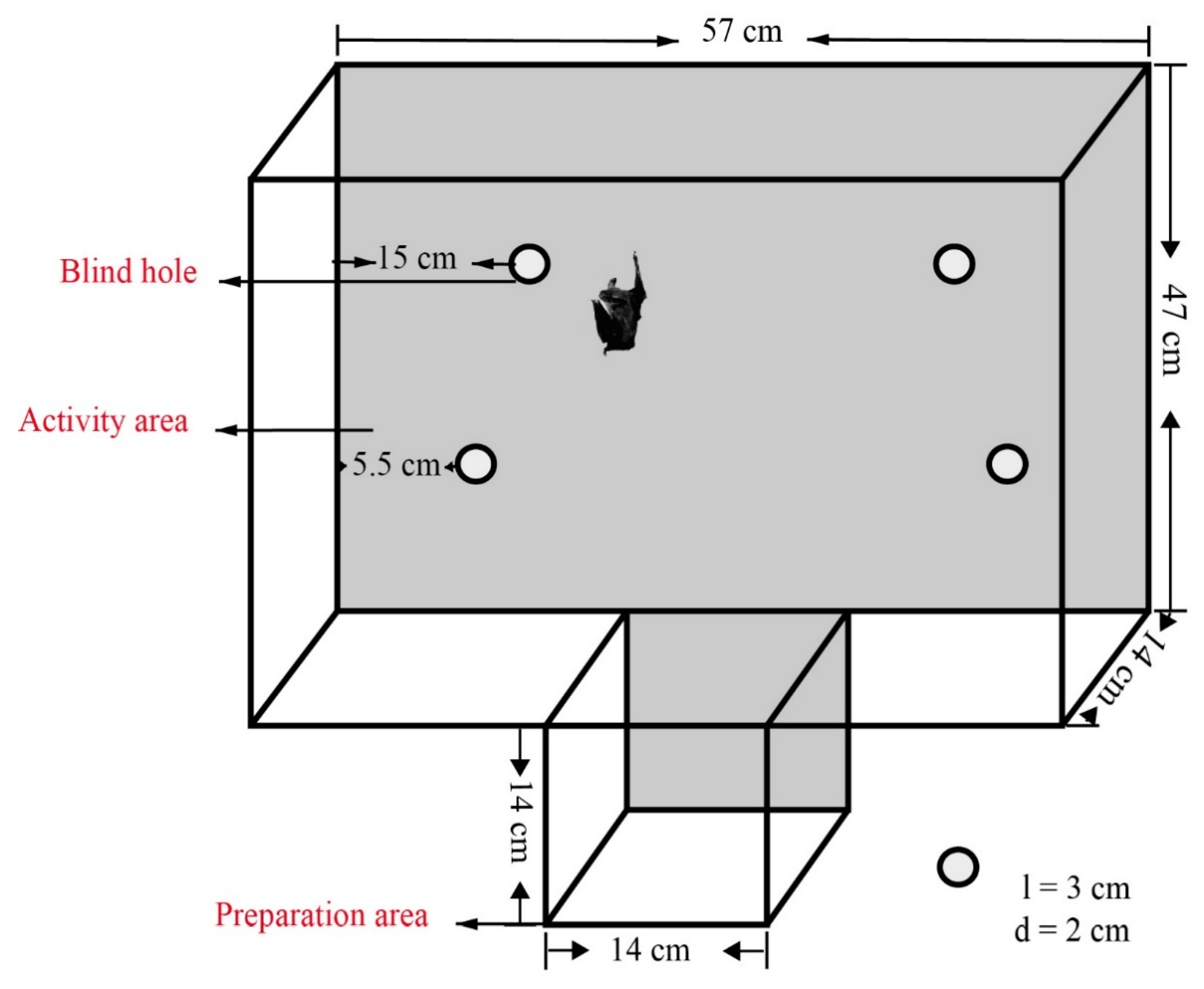
2.2. Hole-Board Test
2.3. Aggression Test
2.4. Morphological Measurements
2.5. Personality Traits Analysis
- (1)
- Latency to enter: the amount of time it takes to enter the activity area from the preparation area.
- (2)
- Frequency of head dips: the number of times bats explored the hole.
- (3)
- Latency to head dip in holes (edge): the time it takes from entering the active area to exploring the edge hole for the first time.
- (4)
- Latency to head hip in holes (center): the time it takes from entering the activity area to exploring the central hole for the first time.
- (5)
- Locomotion: the proportion of time spent climbing and crawling in relation to the total duration of the experiment in the activity area.
- (6)
- Echolocation: the proportion of time spent echolocating in relation to the total duration of the experiment in the activity area.
- (7)
- Grooming: the proportion of time spent grooming in relation to the total duration of the experiment in the activity area.
- (8)
- Urination and defecation: the total number of urination and defecation events.
2.6. Statistical Analysis
2.7. Ethical Statements
3. Results
3.1. Characteristics of Personality
3.2. The Relationships Among Different Personalities
3.3. The Relationships Between Personality and Body Size
4. Discussion
4.1. Characteristics of Personalities
4.2. The Relationships Among Different Personalities
4.3. Effects of Body Size on Personalities
5. Conclusions
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Nettle, D.; PENKE, L. Personality: Bridging the literatures from human psychology and behavioural ecology. Philos. Trans. R. Soc. B 2010, 365, 4043–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.J.; Feeney, W.E.; Marshall, H.H.; Cowlishaw, G.; Heinsohn, R. Animal personality: What are behavioural ecologists measuring? Biol. Rev. 2013, 88, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briffa, M.; Weiss, A. Animal personality. Curr. Biol. 2010, 20, R912–R914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingemanse, N.J.; Wolf, M. Recent models for adaptive personality differences: A review. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3947–3958. [Google Scholar] [CrossRef]
- Sih, A.; Mathot, K.J.; Moiron, M.; Montiglio, P.O.; Wolf, M.; Dingemanse, N.J. Animal personality and state-behaviour feedbacks: A review and guide for empiricists. Trends Ecol. Evol. 2015, 30, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.; Weissing, F.J. An explanatory framework for adaptive personality differences. Philos. Trans. R. Soc. B 2010, 365, 3959–3968. [Google Scholar] [CrossRef]
- Houston, A.I.; McNamara, J.M. Models of Adaptive Behaviour. J. Appl. Ecol. 1999, 38, 491–495. [Google Scholar]
- Kelleher, S.R.; Silla, A.J.; Dingemanse, N.J.; Byrne, P.G. Body size predicts between-individual differences in exploration behaviour in the southern corroboree frog. Anim. Behav. 2017, 129, 161–170. [Google Scholar] [CrossRef]
- Dowling, L.M.; Godin, J.-G.J. Refuge use in a killifish: Influence of body size and nutritional state. Can. J. Zool. 2002, 80, 782–788. [Google Scholar] [CrossRef]
- Mayer, M.; Shine, R.; Brown, G.P. Bigger babies are bolder: Effects of body size on personality of hatchling snakes. Behaviour 2016, 153, 313–323. [Google Scholar] [CrossRef]
- McElreath, R.; Strimling, P. How noisy information and individual asymmetries can make ‘personality’ an adaptation: A simple model. Anim. Behav. 2006, 72, 1135–1139. [Google Scholar] [CrossRef]
- Nyqvist, M.J.; Gozlan, R.E.; Cucherousset, J.; Britton, J.R. Behavioural syndrome in a solitary predator is independent of body size and growth rate. PLoS ONE 2012, 7, e31619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamps, J.A. Growth-mortality tradeoffs and ‘personality traits’ in animals. Ecol. Lett. 2007, 10, 355–363. [Google Scholar] [CrossRef]
- Rodel, H.G.; Meyer, S. Early development influences ontogeny of personality types in young laboratory rats. Dev. Psychobiol. 2011, 53, 601–613. [Google Scholar] [CrossRef]
- Abrahams, M.V.; Cartar, R.V. Within-group variation in the willingness to risk exposure to a predator the influence of species and size. Oikos 2000, 89, 340–344. [Google Scholar] [CrossRef]
- Skalski, G.T.; Gilliam, J.F. Feeding under predation hazard: Testing models of adaptive behavior with stream fish. Am. Nat. 2002, 160, 158–172. [Google Scholar] [CrossRef]
- Krause, J.; Loader, S.P.; McDermott, J.; Ruxton, G.D. Refuge use by fish as a function of body length-related metabolic expenditure and predation risks. Proc. Biol. Sci. 1998, 265, 2373–2379. [Google Scholar] [CrossRef] [Green Version]
- Mathot, K.J.; Nicolaus, M.; Araya-Ajoy, Y.G.; Dingemanse, N.J.; Kempenaers, B.; Grémillet, D. Does metabolic rate predict risk-taking behaviour? A field experiment in a wild passerine bird. Funct. Ecol. 2015, 29, 239–249. [Google Scholar] [CrossRef]
- Bouwhuis, S.; Quinn, J.L.; Sheldon, B.C.; Verhulst, S. Personality and basal metabolic rate in a wild bird population. Oikos 2014, 123, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Sogard, S.M. Size-Selective Mortality in the Juvenile Stage of Teleost Fishes: A Review. Bull. Mar. Sci. 1997, 60, 1129–1157. [Google Scholar]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, M.; Weissing, F.J. Animal personalities: Consequences for ecology and evolution. Trends Ecol. Evol. 2012, 27, 452–461. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; Boer, S.F.D.; Vegt, B.J.V.D.; Reenen, C.G.V.; Hopster, H.; Jong, I.C.D.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Dall, S.R.X.; Houston, A.I.; McNamara, J.M. The behavioural ecology of personality: Consistent individual differences from an adaptive perspective. Ecol. Lett. 2004, 7, 734–739. [Google Scholar] [CrossRef]
- Colléter, M.; Brown, C. Personality traits predict hierarchy rank in male rainbowfish social groups. Anim. Behav. 2011, 81, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Devost, I.; Jones, T.B.; Cauchoix, M.; Montreuil-Spencer, C.; Morand-Ferron, J. Personality does not predict social dominance in wild groups of black-capped chickadees. Anim. Behav. 2016, 122, 67–76. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; de Goede, P. The relation between dominance and exploratory behavior is context-dependent in wild great tits. Behav. Ecol. 2004, 15, 1023–1030. [Google Scholar] [CrossRef]
- Fox, R.A.; Ladage, L.D.; Roth, T.C.; Pravosudov, V.V. Behavioral Profile Predicts Dominance Status in Mountain Chickadees. Anim. Behav. 2009, 77, 1441–1448. [Google Scholar] [CrossRef] [Green Version]
- Buwalda, B.; Koolhaas, J.M.; de Boer, S.F. Trait aggressiveness does not predict social dominance of rats in the Visible Burrow System. Physiol. Behav. 2017, 178, 134–143. [Google Scholar] [CrossRef]
- Martin, J.G.A.; Réale, D. Temperament, risk assessment and habituation to novelty in eastern chipmunks, Tamias striatus. Anim. Behav. 2008, 75, 309–318. [Google Scholar] [CrossRef]
- Baker, K.R. Comparative Personality Assessment of Three Captive Primate Species Macaca nigra, Macaca sylvanus, and Saimiri sciureus. Int. J. Primatol. 2015, 36, 625–646. [Google Scholar] [CrossRef]
- Brandão, A.; Costa, R.; Rodrigues, E.; Vicente, L. Using behaviour observations to study personality in a group of capuchin monkeys (Cebus apella) in captivity. Behaviour 2019, 156, 203–243. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.M. Insights for Behavioral Ecology from Behavioral Syndromes. Adv. Study Behav. 2008, 38, 227–281. [Google Scholar] [PubMed] [Green Version]
- Menzies, A.K.; Timonin, M.E.; McGuire, L.P.; Willis, C.K.R. Personality variation in little brown bats. PLoS ONE 2013, 8, e80230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, H.D.; Gosling, S.D. Personality in Nonhuman Primates A Review and Evaluation of Past Research. Am. J. Primatol. 2010, 72, 653–671. [Google Scholar] [CrossRef] [PubMed]
- Webber, Q.M.R.; McGuire, L.P.; Smith, S.B.; Willis, C.K. Host behaviour, age and sex correlate with ectoparasite prevalence and intensity in a colonial mammal, the little brown bat. Behaviour 2015, 152, 83–105. [Google Scholar] [CrossRef] [Green Version]
- Sagot, M.; Schöner, C.R.; Jago, A.J.; Razik, I.; Chaverri, G. The importance of group vocal behaviour in roost finding. Anim. Behav. 2018, 142, 157–164. [Google Scholar] [CrossRef]
- Chaverri, G.; Gillam, E.H. Repeatability in the contact calling system of Spix’s disc-winged bat (Thyroptera tricolor). R. Soc. Open Sci. 2015, 2, 140197. [Google Scholar] [CrossRef] [Green Version]
- Kilgour, R.J.; Brigham, R.M. The relationships between Behavioural Categories and Social Influences in the Gregarious Big Brown Bat (Eptesicus fuscus). Ethology 2013, 119, 189–198. [Google Scholar] [CrossRef]
- Ammersdorfer, S.; Galinski, S.; Esser, K.H. Effects of aversive experience on the behavior within a custom-made plus maze in the short-tailed fruit bat, Carollia perspicillata. J. Comp. Physiol. 2012, 198, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Nachev, V.; Winter, Y. Behavioral repeatability and choice performance in wild free-flying nectarivorous bats (Glossophaga commissarisi). Behav. Ecol. Sociobiol. 2019, 73, 2. [Google Scholar] [CrossRef]
- Dai, F.; Okazaki, K.; Miyazaki, M.; Maeda, K. The Effect of Roost Environment on Roost Selection by Non-Reproductive and Dispersing Asian Parti-Coloured Bats, Vespertilio sinensis. Mamm. Study 2010, 35, 99–109. [Google Scholar]
- Jin, L.; Wang, J.; Zhang, Z.; Sun, K.; Kanwal, J.S.; Feng, J. Postnatal development of morphological and vocal features in Asian particolored bat, Vespertilio sinensis. Mamm. Biol. 2012, 77, 339–344. [Google Scholar] [CrossRef]
- Rydell, J.; Baagoe, H.J. Vespertilio murinus, Mamm. Species 1994, 467, 1–6. [Google Scholar]
- Bowyer, R.T. Sexual segregation in ruminants: Definitions, hypotheses, and implications for conservation and management. J. Mammal. 2004, 85, 1039–1052. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Jiang, T.; Gu, H.; Liu, H.; Sun, C.; Liu, Y.; Feng, J. Are aggressive vocalizations the honest signals of body size and quality in female Asian particoloured bats? Behav. Ecol. Sociobiol. 2018, 72, 96. [Google Scholar] [CrossRef]
- Zhao, X.; Jiang, T.; Liu, H.; Wang, Y.; Liu, Y.; Feng, J. Acoustic signalling of aggressive intent in the agonistic encounters of female Asian particoloured bats. Anim. Behav. 2019, 149, 65–75. [Google Scholar] [CrossRef]
- Broom, M.; Koenig, A.; Borries, C. Variation in dominance hierarchies among group-living animals: Modeling stability and the likelihood of coalitions. Behav. Ecol. 2009, 20, 844–855. [Google Scholar] [CrossRef]
- Merten, S.; Zwolak, R.; Rychlik, L. Social personality: A more social shrew species exhibits stronger differences in personality types. Anim. Behav. 2017, 127, 125–134. [Google Scholar] [CrossRef]
- Racey, P.A. Ageing and assessment of reproductive status of Pipistrelle bats, Pipistrellus pipistrellus. J. Zool. 2010, 173, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Riebli, T.; Avgan, B.; Bottini, A.-M.; Duc, C.; Taborsky, M.; Heg, D. Behavioural type affects dominance and growth in staged encounters of cooperatively breeding cichlids. Anim. Behav. 2011, 81, 313–323. [Google Scholar] [CrossRef]
- Wilson, A.J.; de Boer, M.; Arnott, G.; Grimmer, A. Integrating personality research and animal contest theory: Aggressiveness in the green swordtail Xiphophorus helleri. PLoS ONE 2011, 6, e28024. [Google Scholar] [CrossRef] [PubMed]
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research. 2018. Available online: https://CRAN.R-project.org/package=psych (accessed on 23 July 2019).
- Timonin, M.E.; Carrière, C.J.; Dudych, A.D.; Latimer, J.G.W.; Unruh, S.T.; Willis, C.K.R. Individual differences in the behavioural responses of meadow voles to an unfamiliar environment are not correlated with variation in resting metabolic rate. J. Zool. 2011, 284, 198–205. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; Computing: Vienna, Austria, 2018. [Google Scholar]
- Kaiser, H.F. Coefficient Alpha for a Principal Component and the Kaiser-Guttman Rule. Psychol. Rep. 1991, 68, 855–858. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Schjelderup-Ebbe, T. Beiträge zur Sozialpsychologie des Haushuhns. Zeitschrift für Psychol. 1922, 88, 225–252. [Google Scholar]
- Liu, H.; Zhao, X.; Wang, Y.; Liu, Y.; Feng, J.; Jiang, T. Effects of body size and prior residence on dominance hierarchies in female Asian particoloured bats. J. Mammal. 2020. accepted. [Google Scholar]
- Lin, F.-J. Solving Multicollinearity in the Process of Fitting Regression Model Using the Nested Estimate Procedure. Qual. Quant. 2008, 42, 417–426. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 133–199. [Google Scholar] [CrossRef]
- Sugiura, N. Further analysts of the data by akaike’ s information criterion and the finite corrections. Commun. Stat.-Theory Methods 1978, 7, 13–26. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference; Burnham, K.P., Anderson, D.R., Eds.; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Bartoń, K. Multi-model inference. Sociol. Method Res. 2016, 33, 261–304. [Google Scholar]
- Lukacs, P.M.; Thompson, W.L.; Kendall, W.L.; Gould, W.R.; Doherty, P.F.; Burnham, K.P.; Anderson, D.R. Concerns regarding a call for pluralism of information theory and hypothesis testing. J. Appl. Ecol. 2007, 44, 456–460. [Google Scholar] [CrossRef]
- Wilson, D.S.; Clark, A.B.; Coleman, K.; Dearstyne, T. Shyness and boldness in humans and other animals. Trends Ecol. Evol. 1994, 9, 442–446. [Google Scholar] [CrossRef]
- Hughes, R.N. Intrinsic exploration in animals motives and measurement. Behav. Process. 1997, 41, 213–226. [Google Scholar] [CrossRef]
- Walsh, R.N.; Cummins, R.A. The open-field test a critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef]
- Archer, J. Tests for emotionality in rats and mice: A review. Anim. Behav. 1973, 21, 205–235. [Google Scholar] [CrossRef]
- Timmermans, P.J.A.; Vochteloo, J.D.; Vossen, J.M.H.; Röder, E.L.; Duijghuisen, J.A.H. Persistent neophobic behaviour in monkeys: A habit or a trait? Behav. Process. 1994, 31, 177–196. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C.; Winkler, H.; Leisler, B. The Significance of Ecological Factors for Exploration and Neophobia in Parrots. Ethology 2002, 108, 249–272. [Google Scholar] [CrossRef]
- Liang, J.; Yang, J.; Xie, H.; Peng, X.; He, X.; Sun, Y.; Zhang, L. Impact of external odor on self-grooming of lesser flat-headed bats, Tylonycteris pachypus. Ecol. Evol. 2019, 9, 8113–8118. [Google Scholar] [CrossRef]
- Antoniadis, E.A.; McDonald, R.J. Amygdala, hippocampus and discriminative fear conditioning to context. Behav. Brain Res. 2000, 108, 1–19. [Google Scholar] [CrossRef]
- Bell, A.M.; Hankison, S.J.; Laskowski, K.L. The repeatability of behaviour: A meta-analysis. Anim. Behav. 2009, 77, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boake, C.R.B. Repeatability its role in evolutionary studies of mating behavior. Evol. Ecol. 1989, 3, 173–182. [Google Scholar] [CrossRef]
- Morinay, J.; Daniel, G.; Gustafsson, L.; Doligez, B. No evidence for behavioural syndrome and genetic basis for three personality traits in a wild bird population. Anim. Behav. 2019, 153, 69–82. [Google Scholar] [CrossRef]
- Verbeek, M.E.M.; Drent, P.J.; Wiepkema, P.R. Consistent individual differences in early exploratory behaviour of male great tits. Anim. Behav. 1994, 48, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Dingemanse, N.J.; Both, C.; Drent, P.J.; van Oers, K.; van Noordwijk, A.J. Repeatability and heritability of exploratory behaviour in great tits from the wild. Anim. Behav. 2002, 64, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Mettke-Hofmann, C.; Rowe, K.C.; Hayden, T.J.; Canoine, V. Effects of experience and object complexity on exploration in garden warblers (Sylvia borin). J. Zool. 2006, 268, 405–413. [Google Scholar] [CrossRef]
- Thompson, R.; Groves, C.P. Habituation: A dual-process theory. Psychol. Rev. 1970, 77, 419. [Google Scholar]
- Via, S.; Gomulkiewicz, R.; Jong, G.D.; Scheiner, S.M.; Schlichting, C.D.; Tienderen, P.H.V. Adaptive phenotypic plasticity consensus and controversy. Trends Ecol. Evol. 1995, 10, 212–217. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Both, C.; Drent, P.J.; Tinbergen, J.M. Fitness consequences of avian personalities in a fluctuating environment. Proc. R. Soc. Lond. Ser. B 2004, 271, 847–852. [Google Scholar] [CrossRef]
- Benesh, D.P.; Valtonen, E.T.; Seppala, O. Multidimensionality and intra-individual variation in host manipulation by an acanthocephalan. Parasitology 2008, 135, 617–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kralj-Fiser, S.; Scheiber, I.B.; Blejec, A.; Moestl, E.; Kotrschal, K. Individualities in a flock of free-roaming greylag geese: Behavioral and physiological consistency over time and across situations. Horm. Behav. 2007, 51, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.B.; Ehlinger, T.J. Pattern and adaptation in individual behavioral differences. In Perspectives in Ethology; Bateson, P.P.G., Klopfer, P.H., Eds.; Plenum: New York, NY, USA, 1987. [Google Scholar]
- Wilson, D.S.; Coleman, K.; Clark, A.B.; Biederman, L. Shy-bold continuum in pumpkinseed sunfish (Lepomis gibbosus): An ecological study of a psychological trait. J. Comp. Psychol. 1993, 107, 250–260. [Google Scholar] [CrossRef]
- Herborn, K.A.; Macleod, R.; Miles, W.T.S.; Schofield, A.N.B.; Alexander, L.; Arnold, K.E. Personality in captivity reflects personality in the wild. Anim. Behav. 2010, 79, 835–843. [Google Scholar] [CrossRef]
- Coleman, K.; Wilson, D.S. Shyness and boldness in pumpkinseed sunfish: Individual differences are context-specific. Anim. Behav. 1998, 56, 927–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingemanse, N.J.; Van der Plas, F.; Wright, J.; Réale, D.; Schrama, M.; Roff, D.A.; Van der Zee, E.; Barber, I. Individual experience and evolutionary history of predation affect expression of heritable variation in fish personality and morphology. Proc. R. Soc. B. Biol. Sci. 2009, 276, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Ward, A.J.W.; Thomas, P.; Hart, P.J.B.; Krause, J. Correlates of boldness in three-spined sticklebacks (Gasterosteus aculeatus). Behav. Ecol. Sociobiol. 2004, 55, 561–568. [Google Scholar] [CrossRef]
- van Oers, K.; Klunder, M.; Drent, P.J. Context dependence of personalities: Risk-taking behavior in a social and a nonsocial situation. Behav. Ecol. 2005, 16, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Webster, M.M.; Ward, A.J.W.; Hart, P.J.B. Boldness is influenced by social context in threespine sticklebacks (Gasterosteus aculeatus). Behaviour 2007, 144, 351–371. [Google Scholar]
- Wilson, A.D.M.; Whattam, E.M.; Bennett, R.; Visanuvimol, L.; Lauzon, C.; Bertram, S.M. Behavioral correlations across activity, mating, exploration, aggression, and antipredator contexts in the European house cricket, Acheta domesticus. Behav. Ecol. Sociobiol. 2009, 64, 703–715. [Google Scholar] [CrossRef] [Green Version]
- Koolhaas, J.M.; De Boer, S.F.; Coppens, C.M.; Buwalda, B. Neuroendocrinology of coping styles: Towards understanding the biology of individual variation. Front. Neuroendocrinol. 2010, 31, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Piersma, T.; Davidson, N.C. Confusion of mass and size. The Auk 1991, 108, 441–443. [Google Scholar]
- Brett, J.R. Environmental Factors and Growth. In Fish Physiology; Academic: Cambridge, MA, USA, 1979; pp. 599–675. [Google Scholar]
- Peters, R.H.; Peters, R.H. Physiological correlates of size. In The Ecological Implications of Body Size; Beck, E., Birks, H.J.B., Conner E., F., Eds.; Cambridge University Press: Cambridge, UK, 1983; Volume 22, pp. 45–53. [Google Scholar]
- Careau, V.; Thomas, D.; Humphries, M.M.; Réale, D. Energy metabolism and animal personality. Oikos 2008, 117, 641–653. [Google Scholar] [CrossRef]
- Harris, S.; Ramnarine, I.W.; Smith, H.G.; Pettersson, L.B. Picking personalities apart estimating the influence of prédation, sex and body size on boldness in the guppy Poecilia reticulata. Oikos 2010, 119, 1711–1718. [Google Scholar] [CrossRef]
- Hewes, M.E.; Chaves-Campos, J.; Wright, J. Boldness related to size in the hermit crab Coenobita compressus at undisturbed, but not disturbed beach. Ethology 2018, 124, 570–578. [Google Scholar] [CrossRef]
- Cooper, W.E.C., Jr.; Frederick, W.G. Optimal time to emerge from refuge. Biol. J. Linn. Soc. 2007, 91, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Funghi, C.; Leitão, A.V.; Ferreira, A.C.; Mota, P.G.; Cardoso, G.C.; Fusani, L. Social Dominance in a Gregarious Bird is Related to Body Size But not to Standard Personality Assays. Ethology 2015, 121, 84–93. [Google Scholar] [CrossRef]
- Parker, G.A. Assessment strategy and the evolution of fighting behaviour. J. Theory Biol. 2001, 47, 223–243. [Google Scholar] [CrossRef]
- Smith, J.M.; Parker, G.A. The logic of asymmetric contests. Anim. Behav. 1976, 24, 159–175. [Google Scholar] [CrossRef]
- Gilbert, C.; McCafferty, D.; Le Maho, Y.; Martrette, J.M.; Giroud, S.; Blanc, S.; Ancel, A. One for all and all for one: The energetic benefits of huddling in endotherms. Biol. Rev. 2010, 85, 545–569. [Google Scholar] [CrossRef]
- Luo, B.; Lu, G.; Chen, K.; Guo, D.; Huang, X.; Liu, Y.; Feng, J. Social calls honestly signal female competitive ability in Asian particoloured bats. Anim. Behav. 2017, 127, 101–108. [Google Scholar] [CrossRef]
- David, M.; Auclair, Y.; Cézilly, F. Personality predicts social dominance in female zebra finches, Taeniopygia guttata, in a feeding context. Anim. Behav. 2011, 81, 219–224. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Variables | Components | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| Latency to enter | −0.26 | 0.77 | 0.06 |
| Frequency of head dips | −0.92 | −0.08 | −0.02 |
| Latency to head dip in holes (edge) | 0.78 | 0.24 | 0.07 |
| Latency to head dip in holes (center) | 0.77 | −012 | −0.05 |
| Locomotion | −0.58 | −0.63 | −0.21 |
| Echolocation | 0.35 | 0.72 | −0.21 |
| Grooming | 0.31 | 0.03 | 0.82 |
| Urination and defecation | −0.43 | −0.06 | 0.60 |
| Standard deviance (% of total variance) | 0.36 | 0.20 | 0.14 |
| Cumulative proportion of total variance | 0.36 | 0.56 | 0.70 |
| Model | Predictive Variables | df | LogL | AICc | Δi | wi |
|---|---|---|---|---|---|---|
| 1 | BM (+) | 4 | −69.398 | 147.6 | 0.00 | 0.461 |
| 2 | HBL (−), BM (+) | 5 | −68.739 | 148.6 | 1.14 | 0.261 |
| 3 | FAL (+), BM (+) | 5 | −69.354 | 150.0 | 2.37 | 0.141 |
| 4 | FAL (+), HBL (−), BM (+) | 6 | −68.673 | 151.2 | 3.57 | 0.078 |
| 5 | Intercept | 3 | −73.280 | 153.1 | 5.41 | 0.031 |
| Estimate | SE | Adjusted SE | z | 95% CI | |
|---|---|---|---|---|---|
| (Intercept) | −2.425 | 2.415 | 2.465 | 0.984 | (−7.263, 2.241) |
| BM | 0.181 | 0.063 | 0.065 | 2.798 | (0.054, 0.308) |
| HBL | −0.043 | 0.040 | 0.041 | 1.049 | (−0.124, 0.033) |
| FAL | 0.025 | 0.069 | 0.071 | 0.351 | (−0.115, 0.159) |
| Model | Predictive Variables | df | LogL | AICc | Δi | wi |
|---|---|---|---|---|---|---|
| 1 | HBL (−), BM (+) | 6 | 34.472 | −54.0 | 0.00 | 0.303 |
| 2 | Intercept | 4 | 31.281 | −53.3 | 0.78 | 0.206 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Shi, B.; Zhao, X.; Feng, J.; Jiang, T. Morphological Correlates of Personality in Female Asian Particolored Bats (Vespertilio sinensis). Animals 2020, 10, 289. https://doi.org/10.3390/ani10020289
Wang Y, Shi B, Zhao X, Feng J, Jiang T. Morphological Correlates of Personality in Female Asian Particolored Bats (Vespertilio sinensis). Animals. 2020; 10(2):289. https://doi.org/10.3390/ani10020289
Chicago/Turabian StyleWang, Yuze, Biye Shi, Xin Zhao, Jiang Feng, and Tinglei Jiang. 2020. "Morphological Correlates of Personality in Female Asian Particolored Bats (Vespertilio sinensis)" Animals 10, no. 2: 289. https://doi.org/10.3390/ani10020289