RNA Sequencing (RNA-Seq) Based Transcriptome Analysis in Immune Response of Holstein Cattle to Killed Vaccine against Bovine Viral Diarrhea Virus Type I

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Vaccination
2.2. Serological Antibody Detection
2.3. RNA Isolation, Library Preparation, and RNA Sequencing (RNA-seq)
2.4. Reads Trimming, Mapping, and Assembly of Sequenced RNA Reads
2.5. Differential Expression Genes (DEGs) Analysis and Clustering
2.6. Functional Annotation and Enrichment Analysis
3. Results
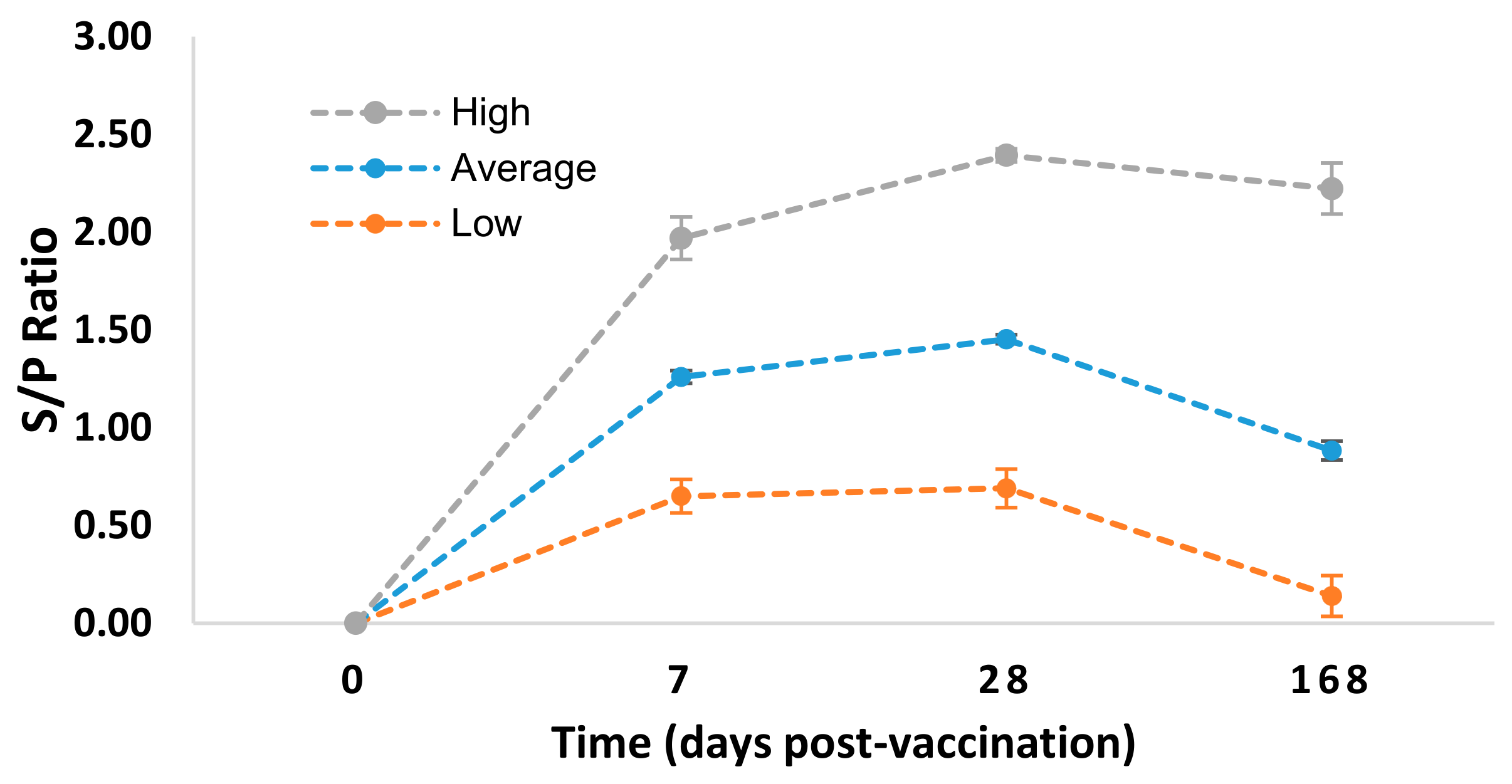
3.1. Experimental Animals
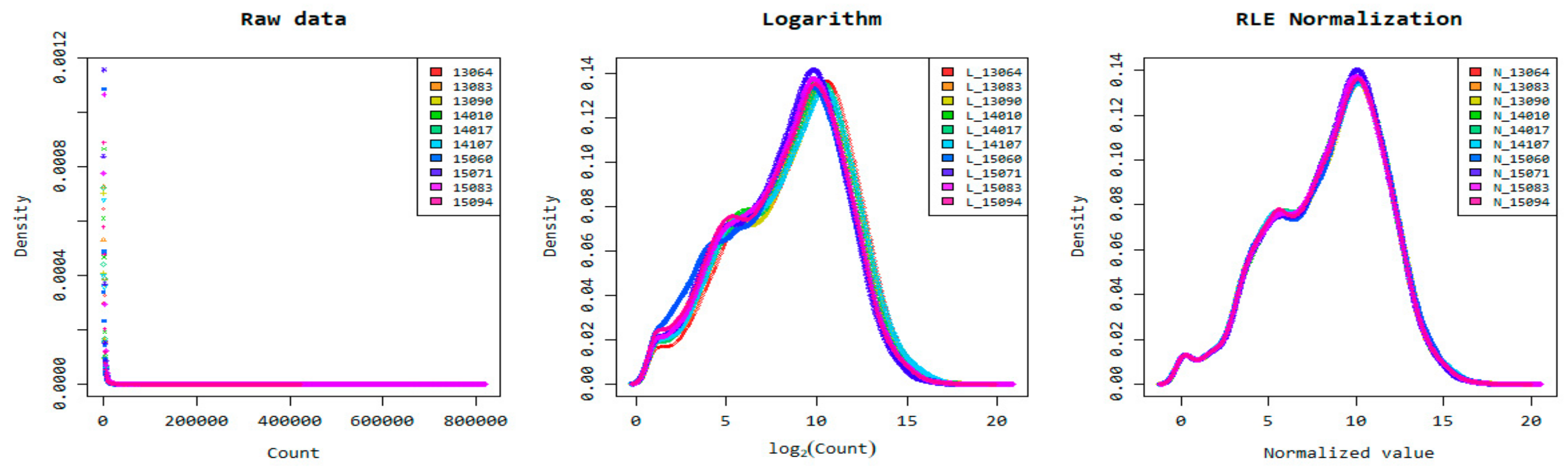
3.2. Transcriptome Sequencing Data
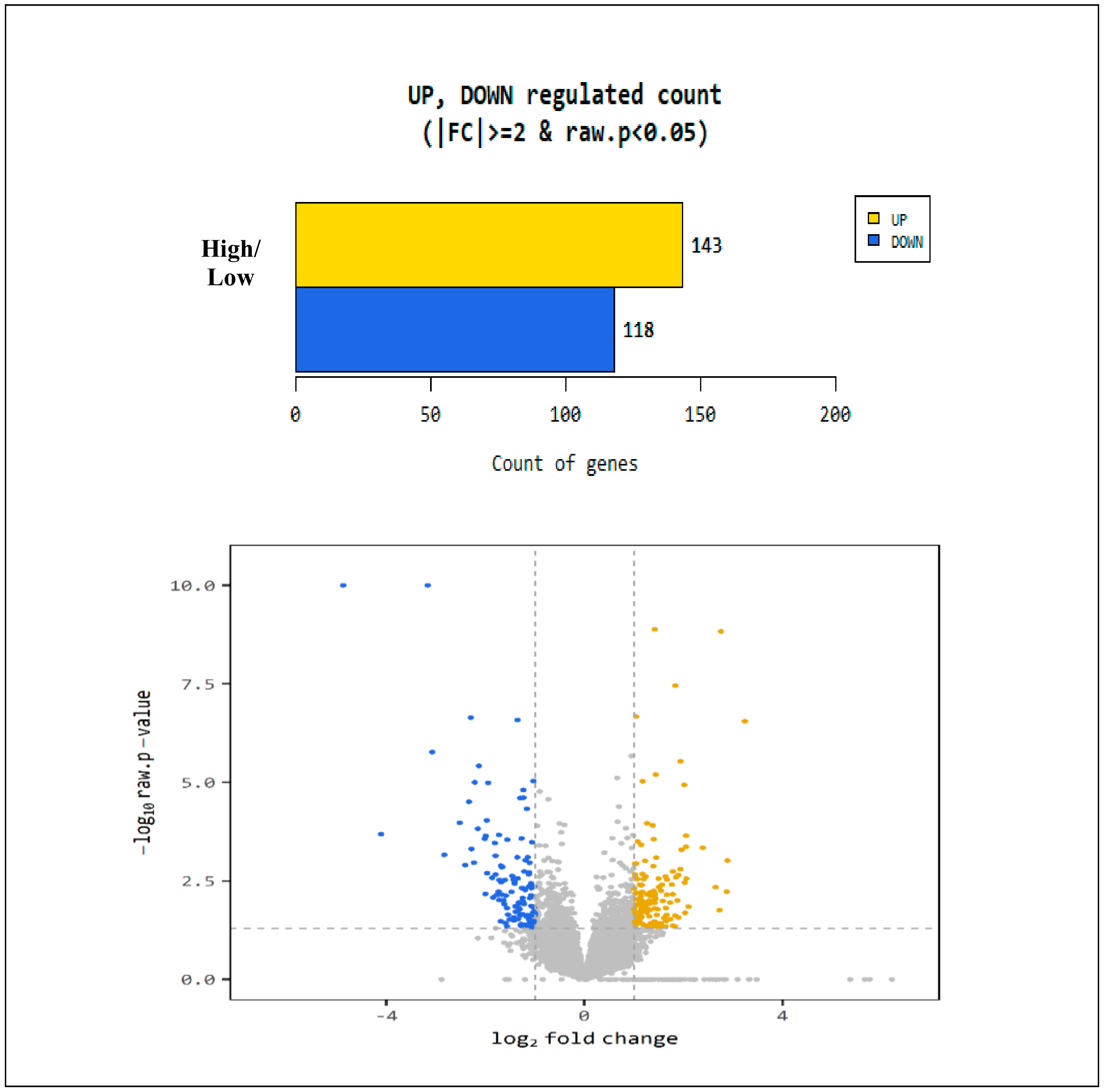
3.3. Differentially Expressed Genes
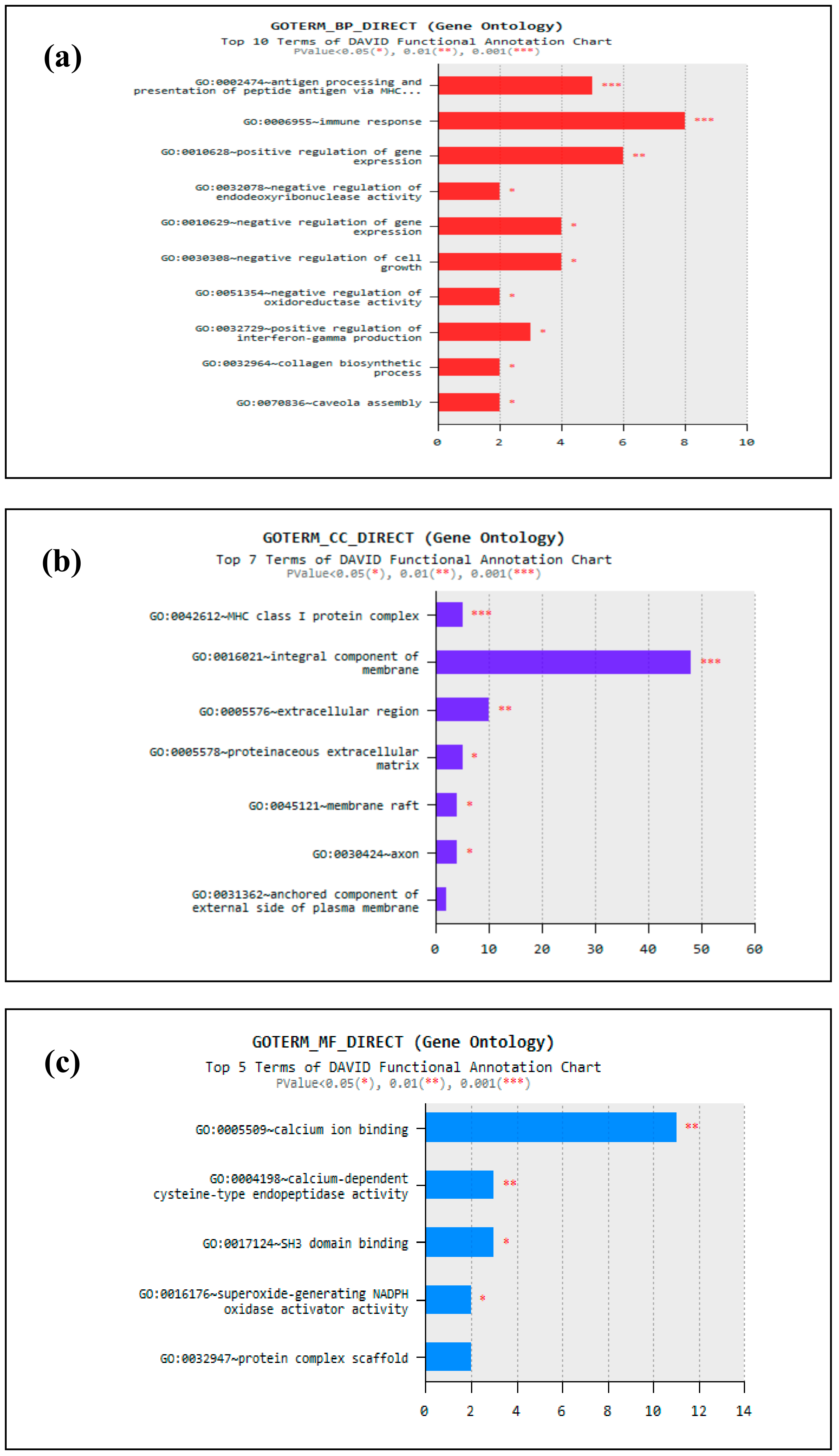
3.4. Functional Enrichment Analysis of Identified DEGs
3.5. KEGG Pathway Enrichment Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Grooms, D.L. Reproductive consequences of infection with bovine viral diarrhea virus. Vet. Clin. N. Am-Food A. 2004, 20, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Khodakaram-Tafti, A.; Farjanikish, G.H. Persistent bovine viral diarrhea virus (BVDV) infection in cattle herds. Iran. J. Vet. Res. 2017, 18, 154–163. [Google Scholar] [CrossRef]
- Pinior, B.; Firth, C.L.; Richter, V.; Lebl, K.; Trauffler, M.; Dzieciol, M.; Hutter, S.E.; Burgstaller, J.; Obritzhauser, W.; Winter, P.; et al. A systematic review of financial and economic assessments of bovine viral diarrhea virus (BVDV) prevention and mitigation activities worldwide. Prev. Vet. Med. 2017, 137, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Mao, L.; Shu, X.; Liu, R.; Hao, F.; Li, J.; Liu, M.; Yang, L.; Zhang, W.; Sun, M.; et al. Transcriptome analysis reveals differential immune related genes expression in bovine viral diarrhea virus-2 infected goat peripheral blood mononuclear cells (PBMCs). BMC Genomics 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, J.; Clarke, M.C.; Howard, C.J.; Pocock, D.H. Pathogenesis and epidemiology of bovine virus diarrhoea virus infection of cattle. Ann. Rech. Vet. 1987, 18, 157–166. [Google Scholar]
- Hansen, T.R.; Smirnova, N.P.; Van Campen, H.; Shoemaker, M.L.; Ptitsyn, A.A.; Bielefeldt-Ohmann, H. Maternal and fetal response to fetal persistent infection with bovine viral diarrhea virus. Am. J. Reprod. Immunol. 2010, 64, 295–306. [Google Scholar] [CrossRef]
- Smirnova, N.P.; Bielefeldt-Ohmann, H.; Van Campen, H.; Austin, K.J.; Han, H.; Montgomery, D.L.; Shoemaker, M.L.; van Olphen, A.L.; Hansen, T.R. Acute non-cytopathic bovine viral diarrhea virus infection induces pronounced type I interferon response in pregnant cows and fetuses. Virus Res. 2008, 132, 49–58. [Google Scholar] [CrossRef]
- Richer, L.; Marois, P.; Lamontagne, L. Association of bovine viral diarrhea virus with multiple viral infections in bovine respiratory disease outbreaks. Can. Vet. J. 1988, 29, 713–717. [Google Scholar]
- Campbell, J.R. Effect of bovine viral diarrhea virus in the feedlot. Vet. Clin. North Am. Food Anim. Pract. 2004, 20, 39–50. [Google Scholar] [CrossRef]
- Zanella, R.; Casas, E.; Snowder, G.; Neibergs, H.L. Fine mapping of loci on BTA2 and BTA26 associated with bovine viral diarrhea persistent infection and linked with bovine respiratory disease in cattle. Front. Genet. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Newcomer, B.W.; Chamorro, M.F.; Walz, P.H. Vaccination of cattle against bovine viral diarrhea virus. Vet. Microbiol. 2017, 206, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Rashid, A.; Rasheed, K.; Akhtar, M. Factors influencing vaccine efficacy - a general review. J. Anim.Plant. Sci. 2009, 19, 22–25. [Google Scholar]
- Niewiesk, S. Maternal antibodies: Clinical significance, mechanism of interference with immune responses, and possible vaccination strategies. Front. Immunol. 2014, 5, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Prasad, M.; Mishra, B.; Manjunath, S.; Sahu, A.R.; Bhuvana Priya, G.; Wani, S.A.; Sahoo, A.P.; Kumar, A.; Balodi, S.; et al. Transcriptome analysis reveals common differential and global gene expression profiles in bluetongue virus serotype 16 (BTV-16) infected peripheral blood mononuclear cells (PBMCs) in sheep and goats. Genomics Data 2017, 11, 62–72. [Google Scholar] [CrossRef]
- Barreto, D.M.; Barros, G.S.; Santos, L.A.B.O.; Soares, R.C.; Batista, M.V.A. Comparative transcriptomic analysis of bovine papillomatosis. BMC Genomics 2018, 19, 949. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Zhao, F.R.; Xie, Y.L.; Liu, Z.Z.; Shao, J.J.; Li, S.F.; Zhang, Y.G.; Chang, H.Y. Transcriptomic analysis of porcine PBMCs in response to FMDV infection. Acta Trop. 2017, 173, 69–75. [Google Scholar] [CrossRef]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. Transcriptome analysis based on RNA-Seq in understanding pathogenic mechanisms of diseases and the immune system of fish: A comprehensive review. Int. J. Mol. Sci. 2018, 19, 245. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Liu, Y.; Liang, L.; Cui, S.; Zhang, Y. RNA-Seq based transcriptome analysis during bovine viral diarrhoea virus (BVDV) infection. BMC Genomics 2019, 20, 774. [Google Scholar] [CrossRef] [Green Version]
- Houe, H. Economic impact of BVDV infection in dairies. Biologicals 2003, 31, 137–143. [Google Scholar] [CrossRef]
- Carman, S.; Van Dreumel, T.; Ridpath, J.; Hazlett, M.; Alves, D.; Dubovi, E.; Tremblay, R.; Bolin, S.; Godkin, A.; Anderson, N. Severe acute bovine viral diarrhea in Ontario, 1993–1995. J. Vet. Diagn. Invest. 1998, 10, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, A.L.E. Bovine viral Diarrhoea virus infections and its control. A review. Vet. Q. 2003, 25, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J.F. Immunology of BVDV vaccines. Biologicals 2013, 41, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Mutz, K.O.; Heilkenbrinker, A.; Lönne, M.; Walter, J.G.; Stahl, F. Transcriptome analysis using next-generation sequencing. Curr. Opin. Biotechnol. 2013, 24, 22–30. [Google Scholar] [CrossRef]
- Hrdlickova, R.; Toloue, M.; Tian, B. RNA-Seq methods for transcriptome analysis. Wiley Interdiscip. Rev. RNA 2017, 8, e1364. [Google Scholar] [CrossRef] [Green Version]
- Garber, M.; Grabherr, M.G.; Guttman, M.; Trapnell, C. Computational methods for transcriptome annotation and quantification using RNA-seq. Nat. Methods 2011, 8, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Buonaguro, L.; Pulendran, B. Immunogenomics and systems biology of vaccines. Immunol. Rev. 2011, 239, 197–208. [Google Scholar] [CrossRef]
- Kaur, B.P.; Secord, E. Innate Immunity. Pediatr. Clin. North Am. 2019, 66, 905–911. [Google Scholar] [CrossRef]
- Bonilla, F.A.; Oettgen, H.C. Adaptive immunity. J. Allergy Clin. Immunol. 2010, 125, S33–S40. [Google Scholar] [CrossRef]
- Bhushan, B.; Patra, B.N.; Das, P.J.; Dutt, T.; Kumar, P.; Sharma, A.; Umang; Dandapat, S.; Ahlawat, S.P.S. Polymorphism of exon 2-3 of bovine major histocompatibility complex class I BoLa-A gene. Genet. Molec. Biol. 2007. [Google Scholar] [CrossRef] [Green Version]
- Untalan, P.M.; Pruett, J.H.; Steelman, C.D. Association of the bovine leukocyte antigen major histocompatibility complex class II DRB3*4401 allele with host resistance to the Lone Star tick, Amblyomma americanum. Vet. Parasitol. 2007, 145, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.F.; Harvey, J.T.; Boothby, J.T. Characterization of a harbor seal class I major histocompatability complex cDNA clone. Immunogenetics 1998, 48, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A.; Travers, P.; Walport, M.; Al, E. Principles of innate and adaptive immunity. Immunobiology: The Immune System in Health and Disease, 5th ed.; Janeway, C., Travers, P., Walport, M., Shlomchik, M., Eds.; Taylor & Francis, Inc.: Abingdon, UK, 2001. [Google Scholar]
- Gutiérrez, S.E.; Esteban, E.N.; Lützelschwab, C.M.; Juliarena, M.A. Major Histocompatibility Complex-Associated Resistance to Infectious Diseases: The Case of Bovine Leukemia Virus Infection. In Trends and Advances in Veterinary Genetics; Abubakar, M., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Weigel, K.A.; Freeman, A.E.; Kehrli, M.E.; Stear, M.J.; Kelley, D.H. Association of Class I Bovine Lymphocyte Antigen Complex Alleles with Health and Production Traits in Dairy Cattle. J. Dairy Sci. 1990, 73, 2538–2546. [Google Scholar] [CrossRef]
- Wawrocki, S.; Druszczynska, M.; Kowalewicz-Kulbat, M.; Rudnicka, W. Interleukin 18 (IL-18) as a target for immune intervention. Acta Biochim. Pol. 2016, 63, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Okamura, H.; Wada, M.; Nagata, K.; Tamura, T. Endotoxin-induced serum factor that stimulates gamma interferon production. Infect. Immun. 1989, 57, 590–595. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.A. Interleukin 1 and interleukin 18 as mediators of inflammation and the aging process. Am. J. Clin. Nutr. 2006, 83, 447S–455S. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, E.M.; Krimpenfort, P.; Berns, A.; Verma, I.M. Immunological defects in mice with a targeted disruption in Bcl-3. Genes Dev. 1997, 11, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Fredericksen, F.; Carrasco, G.; Villalba, M.; Olavarría, V.H. Cytopathic BVDV-1 strain induces immune marker production in bovine cells through the NF-κB signaling pathway. Mol. Immunol. 2015, 68, 213–222. [Google Scholar] [CrossRef]
- Carmody, R.J.; Ruan, Q.; Palmer, S.; Hilliard, B.; Chen, Y.H. Negative regulation of toll-like receptor signaling by NF-κB p50 ubiquitination blockade. Science 2007, 317, 675–678. [Google Scholar] [CrossRef] [Green Version]
- Turrin, N.P.; Plata-Salamán, C.R. Cytokine-cytokine interactions and the brain. Brain Res. Bull. 2000, 51, 3–8. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorokin, L. The impact of the extracellular matrix on inflammation. Nat. Rev. Immunol. 2010, 10, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Giancotti, F.G.; Ruoslahti, E. Integrin Signaling. Science 1999, 285, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Kroll, M.H.; Harris, T.S.; Moake, J.L.; Handin, R.I.; Schafer, A.I. Von Willebrand factor binding to platelet Gplb initiates signals for platelet activation. J. Clin. Invest. 1991, 88, 1568–1573. [Google Scholar] [CrossRef]
- Englund, G.D.; Bodnar, R.J.; Li, Z.; Ruggeri, Z.M.; Du, X. Regulation of von Willebrand Factor Binding to the Platelet Glycoprotein Ib-IX by a Membrane Skeleton-dependent Inside-out Signal. J. Biol. Chem. 2001, 276, 16952–16959. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.Z.; Mo, X.; Afshar-Kharghan, V.; Srinivasan, S.; López, J.A.; Li, R. Glycoprotein Ibα forms disulfide bonds with 2 glycoprotein Ibβ subunits in the resting platelet. Blood 2007, 109, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Klein, G. The extracellular matrix of the hematopoietic microenvironment. Experientia 1995, 51, 914–926. [Google Scholar] [CrossRef]
- Assinger, A. Platelets and infection - An emerging role of platelets in viral infection. Front. Immunol. 2014, 5, 649. [Google Scholar] [CrossRef] [Green Version]
- Procko, E.; Gaudet, R. Antigen processing and presentation: TAPping into ABC transporters. Curr. Opin. Immunol. 2009, 21, 84–91. [Google Scholar] [CrossRef]
- Seyffer, F.; Tampé, R. ABC transporters in adaptive immunity. BBA-Gen Subjects 2015, 1850, 449–460. [Google Scholar] [CrossRef]
- Hinz, A.; Tampé, R. ABC transporters and immunity: Mechanism of self-defense. Biochemistry 2012, 51, 4981–4989. [Google Scholar] [CrossRef] [PubMed]
- Tajima, M.; Yazawa, T.; Hagiwara, K.; Kurosawa, T.; Takahashi, K. Diabetes Mellitus in Cattle Infected with Bovine Viral Diarrhea Mucosal Disease Virus. J. Vet. Med. 1992, 39, 616–620. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Total Raw Reads | Total Clean Reads | GC (%) | Q30 (%) | No. of Processed Reads | No. of Mapped Reads | No. of Unmapped Reads |
|---|---|---|---|---|---|---|---|
| 13064 | 1.13E + 10 | 1.12E + 08 | 45.52 | 95.28 | 1.11E + 08 | 1.08E + 08 | 2.32E + 06 |
| 13083 | 9.59E + 09 | 9.50E + 07 | 45.73 | 95.27 | 9.40E + 07 | 9.12E + 07 | 2.74E + 06 |
| 13090 | 9.77E + 09 | 9.67E + 07 | 45.22 | 94.65 | 9.55E + 07 | 9.38E + 07 | 1.70E + 06 |
| 14010 | 8.52E + 09 | 8.44E + 07 | 45.00 | 95.31 | 8.36E + 07 | 8.17E + 07 | 1.82E + 06 |
| 14017 | 9.38E + 09 | 9.29E + 07 | 45.99 | 95.30 | 9.19E + 07 | 8.93E + 07 | 2.55E + 06 |
| 14107 * | 9.67E + 09 | 9.58E + 07 | 45.03 | 95.22 | 9.47E + 07 | 9.27E + 07 | 2.02E + 06 |
| 15060 * | 8.31E + 09 | 8.22E + 07 | 45.54 | 95.08 | 8.13E + 07 | 7.96E + 07 | 1.62E + 06 |
| 15071 * | 9.01E + 09 | 8.92E + 07 | 45.87 | 95.23 | 8.83E + 07 | 8.18E + 07 | 6.37E + 06 |
| 15083 * | 8.37E + 09 | 8.29E + 07 | 44.48 | 95.19 | 8.20E + 07 | 8.02E + 07 | 1.77E + 06 |
| 15094 * | 7.42E + 09 | 7.35E + 07 | 44.86 | 94.98 | 7.26E + 07 | 7.07E + 07 | 1.92E + 06 |
| Pathways | ID | DEGs No. | p-Value | Up-Regulated Genes | Down-Regulated Genes | |
|---|---|---|---|---|---|---|
| 1 | Platelet activation | bta04611 | 5 | 1.34E-02 | GP5, P2RX1, GP1BA, GP9 | MAPK12 |
| 2 | Cytokine-cytokine receptor interaction | bta04060 | 6 | 2.07E-02 | IL18, IL1RAP | CCR8, CCL3, IL20RA,TGFB2 |
| 3 | ECM-receptor interaction | bta04512 | 4 | 2.52E-02 | GP5, GP1BA, GP9 | ITGB4 |
| 4 | Hematopoietic cell lineage | bta04640 | 4 | 2.91E-02 | GP5, GP1BA, CD24, GP9 | - |
| 5 | ABC transporters | bta02010 | 3 | 3.73E-02 | ABCB11 | LOC100296627, CFTR |
| 6 | Type I diabetes mellitus | bta04940 | 2 | 5.48E-02 | BOLA, PTPRN2 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, B.I.; Santiago, K.G.; Lee, D.; Ha, S.; Seo, K. RNA Sequencing (RNA-Seq) Based Transcriptome Analysis in Immune Response of Holstein Cattle to Killed Vaccine against Bovine Viral Diarrhea Virus Type I. Animals 2020, 10, 344. https://doi.org/10.3390/ani10020344
Lopez BI, Santiago KG, Lee D, Ha S, Seo K. RNA Sequencing (RNA-Seq) Based Transcriptome Analysis in Immune Response of Holstein Cattle to Killed Vaccine against Bovine Viral Diarrhea Virus Type I. Animals. 2020; 10(2):344. https://doi.org/10.3390/ani10020344
Chicago/Turabian StyleLopez, Bryan Irvine, Kier Gumangan Santiago, Donghui Lee, Seungmin Ha, and Kangseok Seo. 2020. "RNA Sequencing (RNA-Seq) Based Transcriptome Analysis in Immune Response of Holstein Cattle to Killed Vaccine against Bovine Viral Diarrhea Virus Type I" Animals 10, no. 2: 344. https://doi.org/10.3390/ani10020344
APA StyleLopez, B. I., Santiago, K. G., Lee, D., Ha, S., & Seo, K. (2020). RNA Sequencing (RNA-Seq) Based Transcriptome Analysis in Immune Response of Holstein Cattle to Killed Vaccine against Bovine Viral Diarrhea Virus Type I. Animals, 10(2), 344. https://doi.org/10.3390/ani10020344