Lions Panthera leo Prefer Killing Certain Cattle Bos taurus Types

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
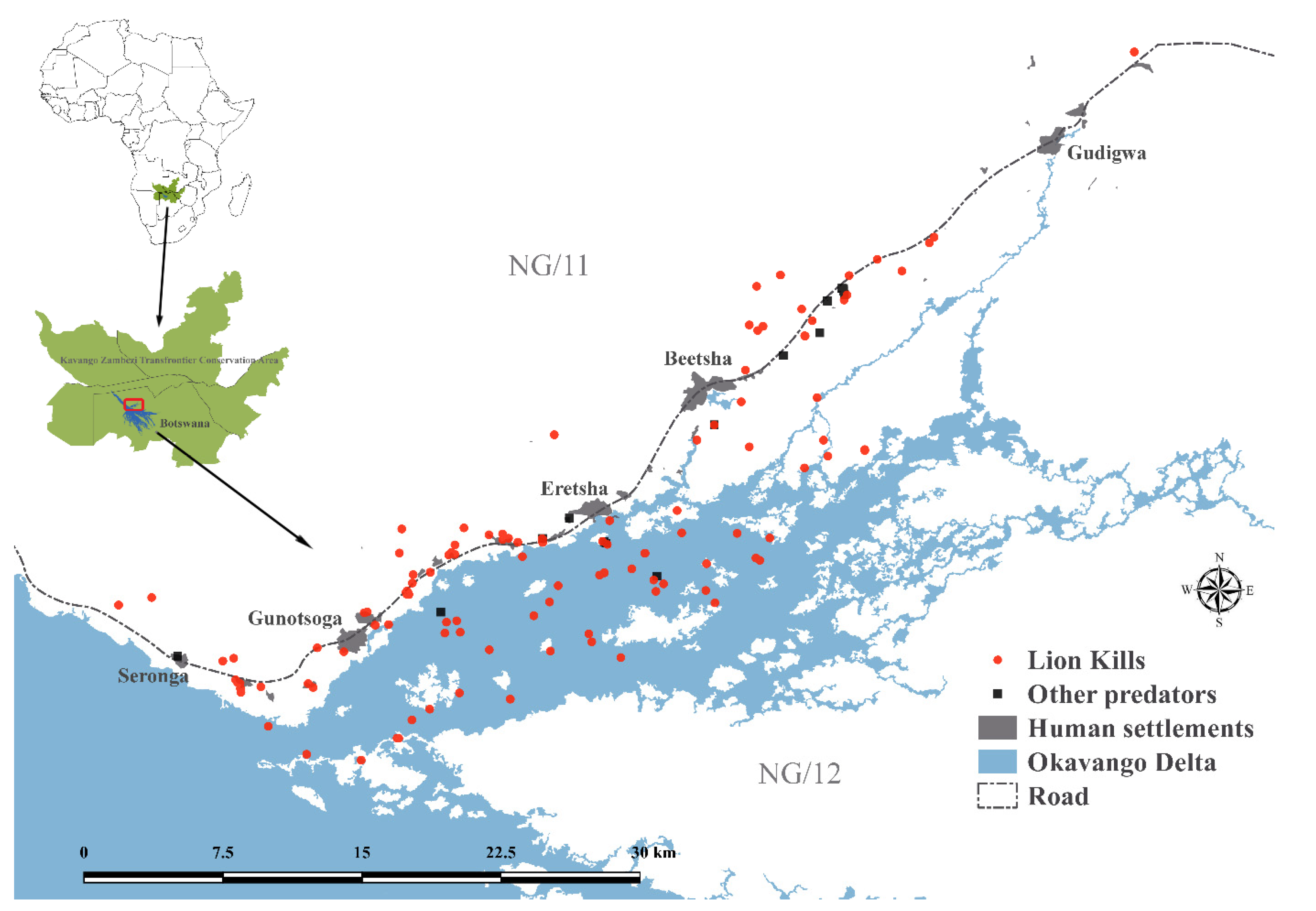
2.1. Study Site
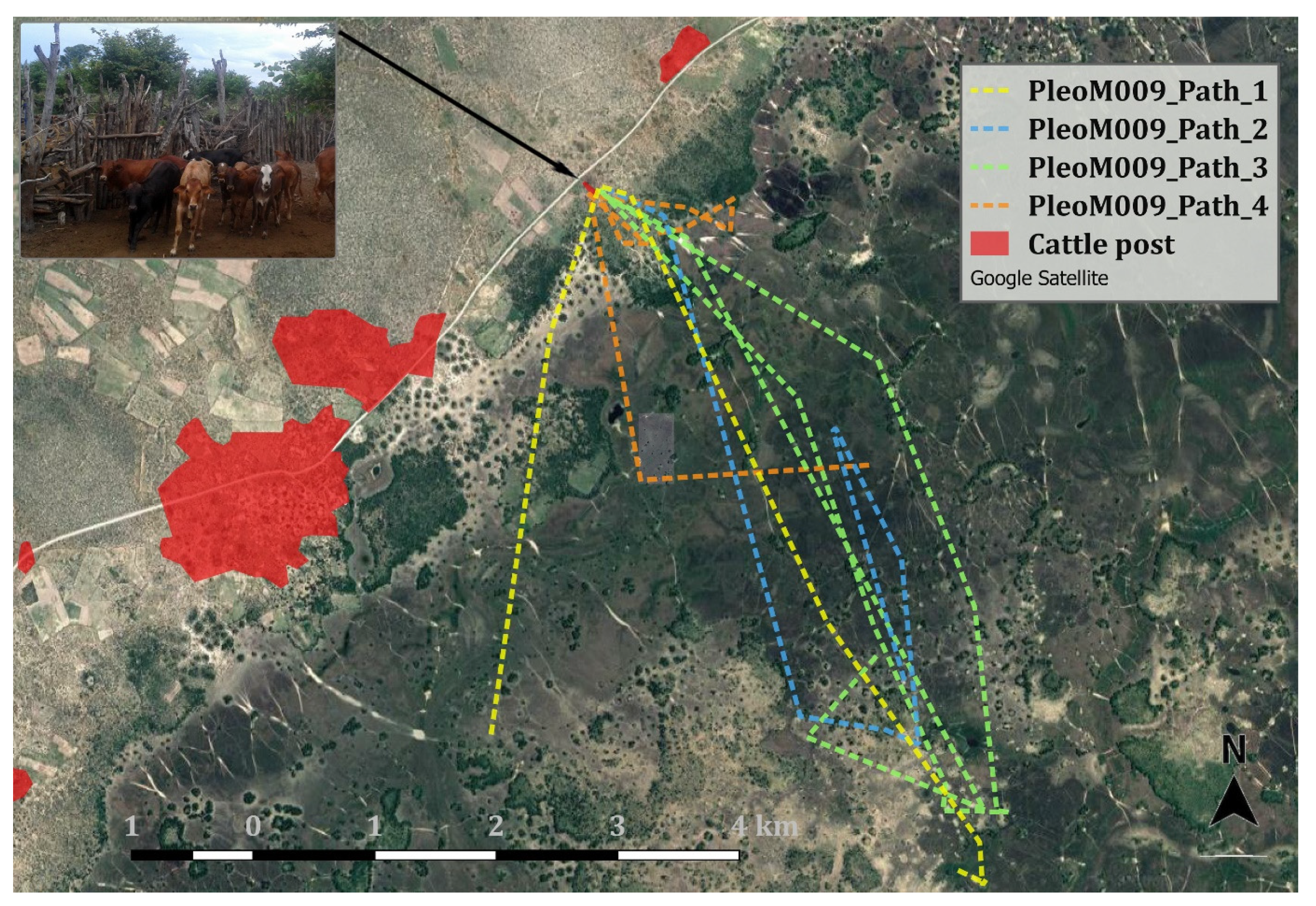
2.2. Survey Methods
2.3. Statistics
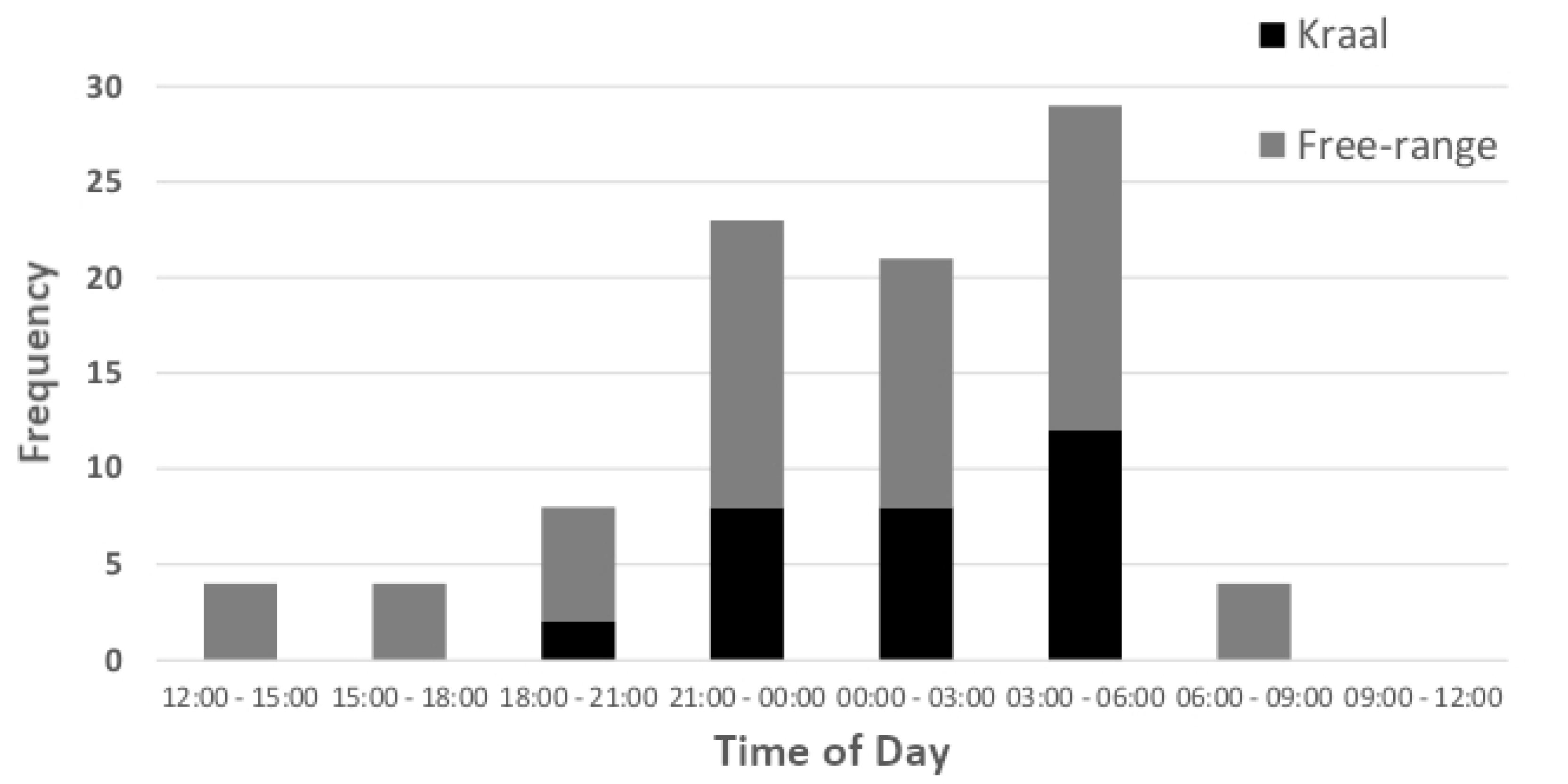
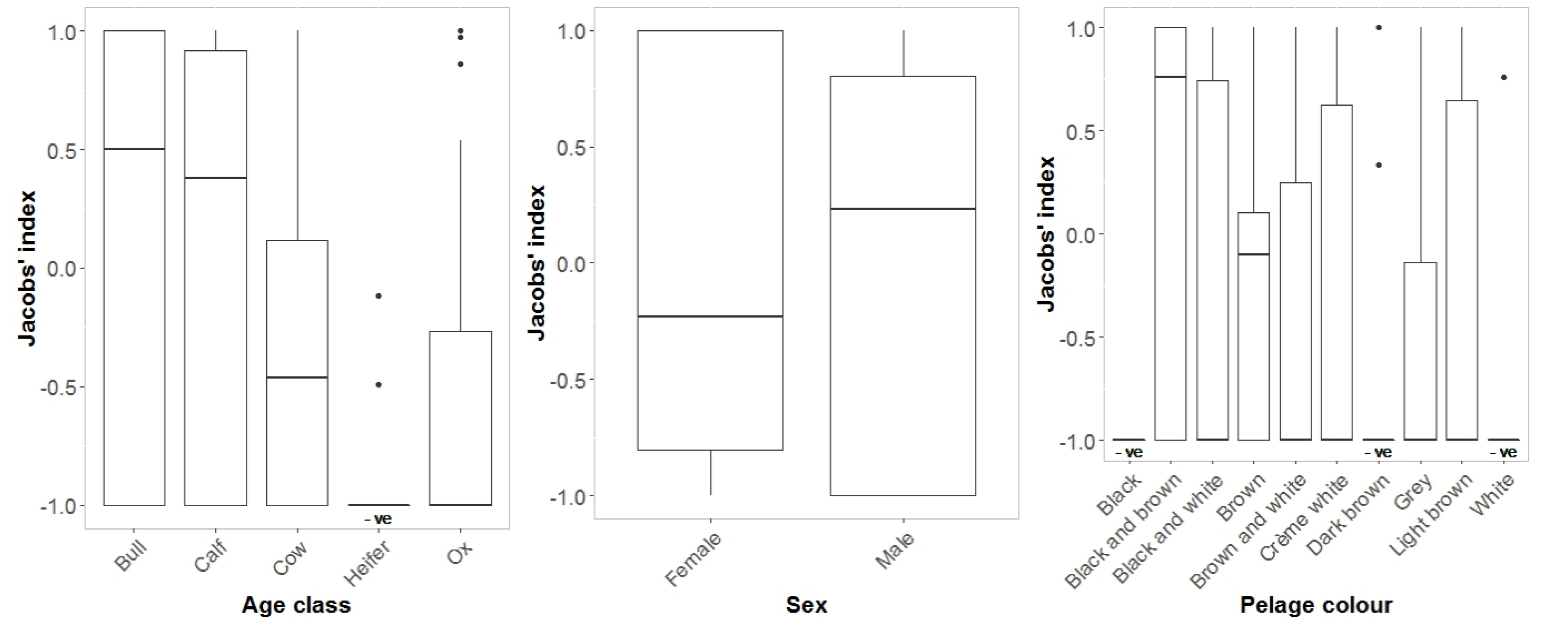
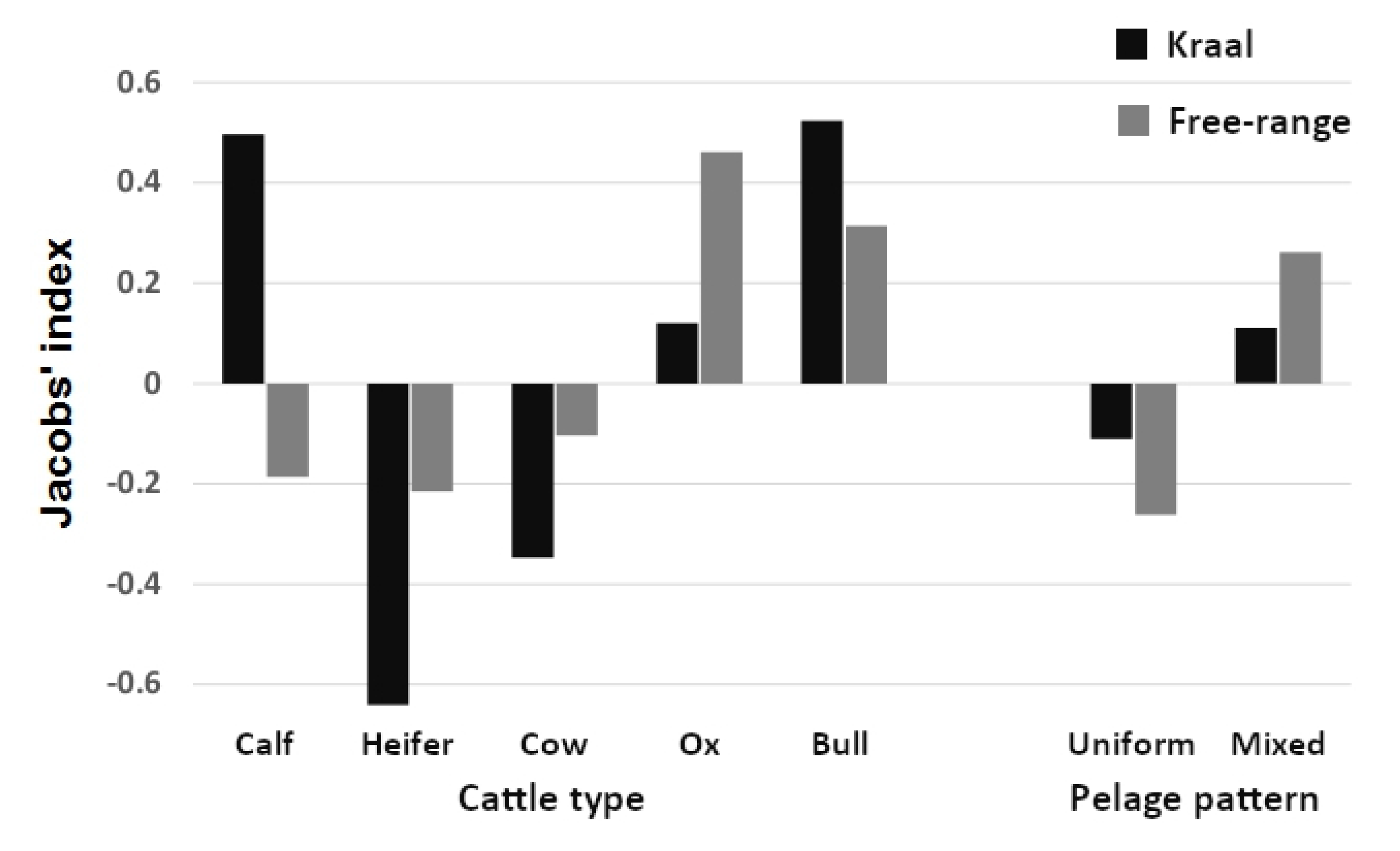
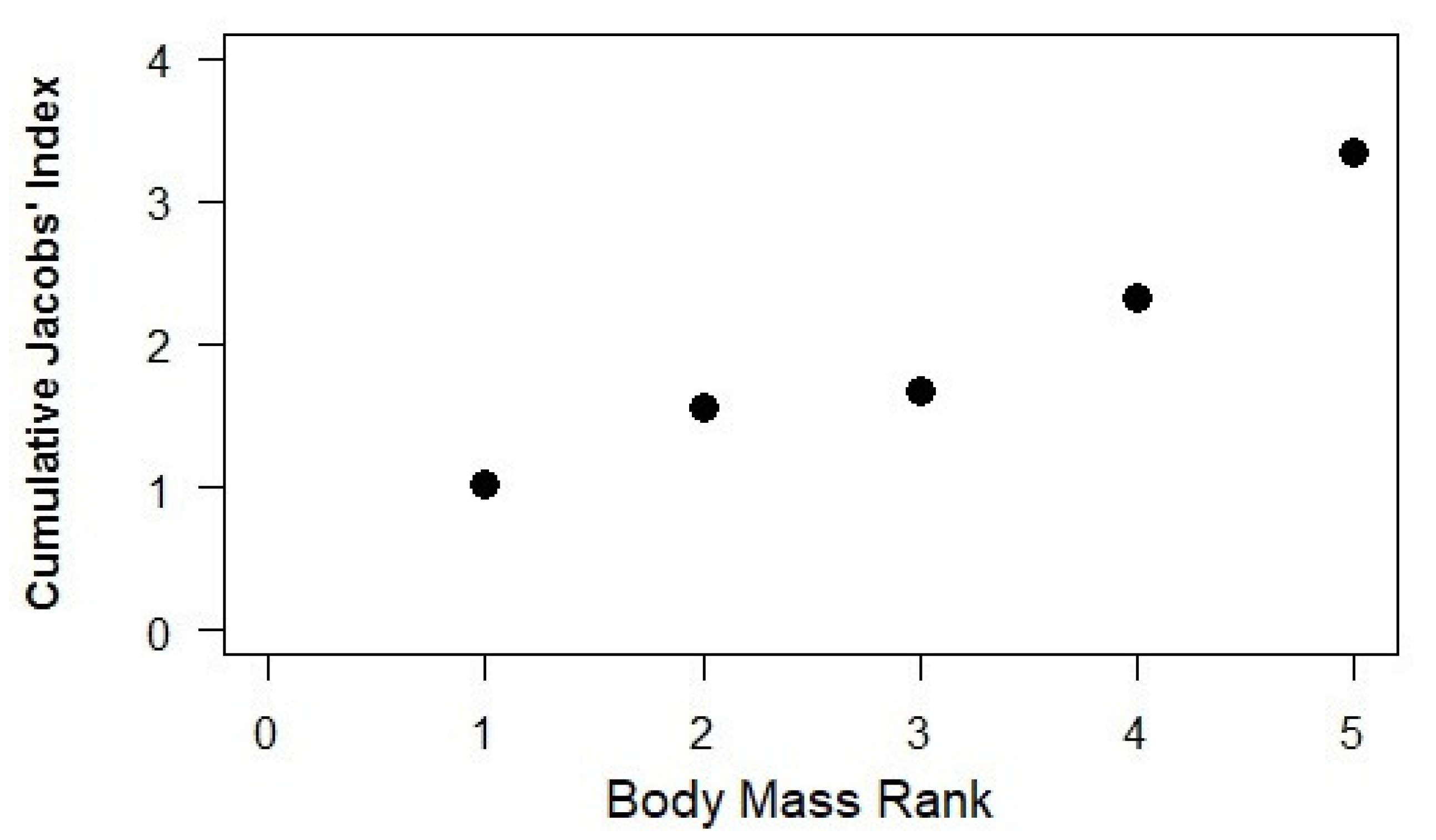
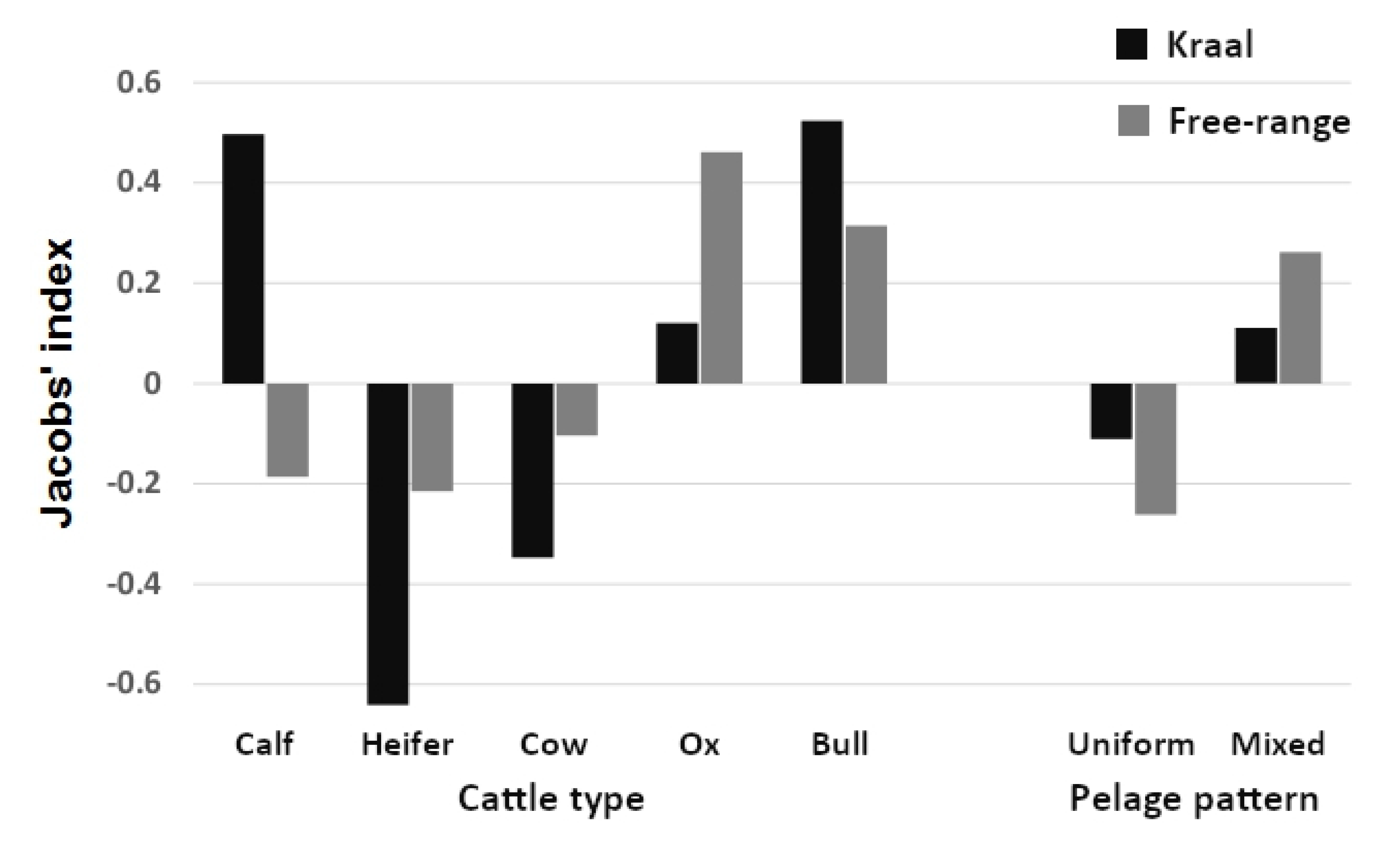
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hayward, M.W.; Kerley, G.I.H. Prey preferences of the lion (Panthera leo). J. Zool. 2005, 267, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Clements, H.S.; Tambling, C.J.; Hayward, M.W.; Kerley, G.I.H. An objective approach to determining the weight ranges of prey preferred by and accessible to the five large African carnivores. PLoS ONE 2014, 9, e101054. [Google Scholar]
- Davidson, Z.; Valeix, M.; Van Kesteren, F.; Loveridge, A.J.; Hunt, J.E.; Murindagomo, F. Seasonal Diet and Prey Preference of the African Lion in a Waterhole-Driven Semi-Arid Savanna. PLoS ONE 2013, 8, e55182. [Google Scholar]
- Hayward, M.W.; Hofmeyr, M.; O’brien, J.; Kerley, G.I.H. Testing predictions of the prey of the lion (Panthera leo) derived from modelled prey preferences. J. Wildl. Manag. 2007, 71, 1567–1575. [Google Scholar] [CrossRef]
- Sinclair, A.R.E. The African Buffalo; University of Chicago Press: Chicago, IL, USA, 1977. [Google Scholar]
- Weise, F.J.; Fynn, R.W.S.; Stein, A.B.; Tomeletso, M.; Somers, M.J.; Periquet, S. Seasonal selection of key resources by cattle in a mixed savanna-wetland ecosystem increases the potential for conflict with lions. Biol. Conserv. 2019, 237, 253–266. [Google Scholar] [CrossRef]
- Valeix, M.; Hemson, G.; Loveridge, A.J.; Mills, G.; Macdonald, D.W. Behavioural adjustments of a large carnivore to access secondary prey in a human-dominated landscape. J. Appl. Ecol. 2012, 49, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Meena, V.; Jhala, Y.V.; Chellam, R.; Pathak, B. Implications of diet composition of Asiatic lions for their conservation. J. Zool. 2011, 284, 60–67. [Google Scholar] [CrossRef]
- Bauer, H.; De Iongh, H.H. Lion (Panthera leo) home ranges and livestock conflicts in Waza National Park, Cameroon. Afr. J. Ecol. 2005, 43, 208–214. [Google Scholar] [CrossRef]
- Funston, P.J. Kalahari Transfrontier Lion Project. Available online: http://www.satibtrust.com/projects/kgalagadi-lion-project/ (accessed on 11 December 2018).
- Hazzah, L.; Mulder, M.B.; Frank, L. Lions and warriors: Social factors underlying declining African lion populations and the effect of incentive-based management in Kenya. Biol. Conserv. 2009, 142, 2428–2437. [Google Scholar] [CrossRef]
- Montgomery, R.A.; Hoffmann, C.F.; Tans, E.D.; Kissui, B. Discordant scales and the potential pitfalls for human-carnivore conflict mitigation. Biol. Conserv. 2018, 224, 170–177. [Google Scholar] [CrossRef]
- Weise, F.J.; Hayward, M.W.; Cassillas Aguirre, R.; Tomeletso, M.; Gadimang, P.; Somers, M.J.; Stein, A.B. Size, shape and maintenance matter: A critical appraisal of a global carnivore conflict mitigation strategy—Livestock protection kraals in northern Botswana. Biol. Conserv. 2018, 225, 88–97. [Google Scholar]
- Van Vuure, T. History, morphology and ecology of the aurochs (Bos primigenius). Lutra 2002, 45, 1–16. [Google Scholar]
- Thornton, P.K. Livestock production: Recent trends, future prospects. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furstenburg, D. Focus on the African buffalo (Syncerus caffer). S A Hunter 2010, 05040, 46–49. [Google Scholar]
- Kyselý, R. Aurochs and potential crossbreeding with domestic cattle in Central Europe in the Eneolithic period. A metric analysis of bones from the archaeological site of Kutná Hora-Denemark (Czech Republic). Anthropozoologica 2008, 43, 7–37. [Google Scholar]
- Masilo, B.S.; Podisi, B. Field Guide to Livestock Breeds in Botswana; Department of Agricultural Research, Ministry of Agriculture: Gaborone, Botswana, 2001. [Google Scholar]
- Raphaka, K.; Dzama, K. Sex of calf and age of dam adjustment factors for birth and weaning weight in Tswana and Composite beef cattle breeds in Botswana. S. Afr. J. Anim. Sci. 2009, 39, 296–300. [Google Scholar]
- Wikipedia: Auerochse. Available online: https://de.wikipedia.org/wiki/Auerochse (accessed on 11 December 2018).
- Murray-Hudson, M. Floodplain Vegetation Responses to Flood Regime in the Seasonal Okavango Delta, Botswana. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2009. [Google Scholar]
- Sianga, K.; Fynn, R. The vegetation and wildlife habitats of the Savuti-Mababe-Linyanti ecosystem, northern Botswana. Koedoe 2017, 59, a1406. [Google Scholar] [CrossRef] [Green Version]
- Fynn, R.W.S.; Murray-Hudson, M.; Dhliwayo, M.; Scholte, P. African wetlands and their seasonal use by wild and domestic herbivores. Wetl. Ecol. Manag. 2015, 23, 559–581. [Google Scholar] [CrossRef]
- Bauer, H.; Chapron, G.; Nowell, K.; Henschel, P.; Funston, P.; Hunter, L.T.B.; Macdonald, D.W.; Packer, C. Lion (Panthera leo) populations are declining rapidly across Africa, except in intensively managed areas. PNAS 2015, 112, 14894–14899. [Google Scholar] [CrossRef] [Green Version]
- Riggio, J.; Jacobson, A.; Dollar, L.; Bauer, H.; Becker, M.; Dickman, A.; Funston, P.; Groom, R.; Henschel, P.; de Longh, H.; et al. The size of savannah Africa: A lion’s (Panthera leo) view. Biodivers. Conserv. 2013, 22, 17–35. [Google Scholar] [CrossRef] [Green Version]
- Department of Wildlife and National Parks. Compensation Guidelines for Damages Caused by Elephants and Lion; Department of Wildlife and National Parks, Ministry of Environment, Wildlife and Tourism of the Government of Botswana: Gaborone, Botswana, 2013. [Google Scholar]
- Weise, F.J.; Hauptmeier, H.; Stratford, K.J.; Hayward, M.W.; Aal, K.; Heuer, M.; Tomeletso, M.; Wulf, V.; Somers, M.J.; Stein, A.B. Lions at the Gates: Trans-disciplinary Design of an Early Warning System to Improve Human-Lion Coexistence. Front. Ecol. Evol. 2019, 6, 242. [Google Scholar] [CrossRef] [Green Version]
- Lethola, L.L.; Buck, N.G.; Light, D.E. Beef cattle breeding in Botswana. Botsw. Notes Rec. 1983, 15, 39–47. [Google Scholar]
- Rahway, N.J. Genus Bos: Cattle Breeds of the World. MSO-AGVET. Merck Co., Inc. Available online: http://afs.okstate.edu/breeds/cattle/tswana/ (accessed on 11 December 2018).
- Trail, J.C.M.; Buck, N.G.; Light, D.; Rennie, T.W.; Rutherford, A.; Miller, M.; Pratchett, D.; Capper, B.S. Productivity of Africander, Tswana, Tuli and crossbred beef cattle in Botswana. Anim. Sci. 1977, 24, 57–62. [Google Scholar] [CrossRef]
- Stander, P.E.; Ghau, I.I.; Tsisaba, D.; Oma, I.I.; Vi, |.|. Tracking and the interpretation of spoor: A scientifically sound method in ecology. J. Zool. 1997, 242, 329–341. [Google Scholar] [CrossRef]
- Jacobs, J. Quantitative measurement of food selection. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Hayward, M.W.; Porter, L.; Lanszki, J.; Kamler, J.F.; Beck, J.M.; Kerley, G.I.H.; Macdonald, D.W.; Montgomery, R.A.; Parker, D.M.; Scott, D.M.; et al. Factors affecting the prey preferences of jackals (Canidae). Mamm. Biol. 2017, 85, 70–82. [Google Scholar] [CrossRef]
- Muggeo, V. Segmented: An R package to fit regression models with broken-line relationships. R News 2008, 8, 20–25. [Google Scholar]
- Kruuk, H. Surplus killing by carnivores. J. Zool. 1972, 166, 233–244. [Google Scholar] [CrossRef]
- Suraci, J.; Frank, L.G.; Oriol Cotterill, A.; Ekwanga, S.; Williams, T.; Wilmers, C. Behavior-specific habitat selection by African lions may promote their persistence in a human-dominated landscape. Ecology 2019, 100, e02644. [Google Scholar] [CrossRef]
- Stander, P.E. A suggested management strategy for stock-raiding lions in Namibia. S. Afr. J. Wildl. Res. 1990, 20, 37–42. [Google Scholar]
- Oriol-Cotterill, A.; Macdonald, D.W.; Valeix, M.; Ekwanga, S.; Frank, L. Spatiotemporal patterns of lion space use in a human-dominated landscape. Anim. Behav. 2015, 101, 27–39. [Google Scholar] [CrossRef]
- Hemson, G.; Maclennan, S.; Mills, G.; Johnson, P.; Macdonald, D.W. Community, lions, livestock and money: A spatial and social analysis of attitudes to wildlife and the conservation value of tourism in a human-carnivore conflict in Botswana. Biol. Conserv. 2009, 142, 2718–2725. [Google Scholar] [CrossRef]
- Loveridge, A.J.; Kuiper, T.; Parry, R.H.; Sibanda, L.; Hunt, J.H.; Stapelkamp, B.; Sebele, L.; Macdonald, D.W. Bells, bomas and beefsteak: Complex patterns of human-predator conflict at the wildlife-agropastoral interface in Zimbabwe. PeerJ 2017, 5, e2898. [Google Scholar] [CrossRef] [PubMed]
- Tumenta, P.N.; Visser, H.D.; Van Rijssel, J.; Müller, L.; De Iongh, H.H.; Funston, P.J.; Udo de Haes, H.A. Lion predation on livestock and native wildlife in Waza National Park, northern Cameroon. Mammalia 2013, 77, 247–251. [Google Scholar] [CrossRef]
- Green, B.; Anderson, J.; Whateley, A. Water and sodium turnover and estimated food consumption in free-living lions (Panthera leo) and spotted hyaenas (Crocuta crocuta). J. Mammal. 1984, 65, 593–599. [Google Scholar] [CrossRef]
- Hayward, M.W.; Hayward, G.J.; Druce, D.; Kerley, G.I.H. Do fences constrain predator movements on an evolutionary scale? Home range, food intake and movement patterns of large predators reintroduced to Addo Elephant National Park, South Africa. Biodiv. Conserv. 2009, 18, 887–899. [Google Scholar] [CrossRef]
- Stander, P.E. Demography of lions in the Etosha National Park, Namibia. Madoqua 1991, 18, 1–9. [Google Scholar]
- Roberts, S.C. The evolution of hornedness in female ruminants. Behaviour 1996, 133, 399–442. [Google Scholar] [CrossRef]
- Caro, T.; Stankowich, T. The function of contrasting pelage markings in artiodactyls. Behav. Ecol. 2010, 21, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Melin, A.D.; Kline, D.W.; Hiramatsu, C.; Caro, T. Zebra stripes through the eyes of their predators, zebras, and humans. PLoS ONE 2016, 11, e0145679. [Google Scholar] [CrossRef] [Green Version]
- Ahnelt, P.K.; Schubert, C.; Kübber-Heiss, A.; Schiviz, A.; Anger, E. Independent variation of retinal S and M cone photoreceptor topographies: A survey of four families of mammals. Vis. Neurosci. 2006, 23, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Whitteridge, D. Ophthalmoscopic studies on the eyes of mammals. Philos. Trans. R. Soc. Lond. B. Sci. 1968, 254, 207–220. [Google Scholar]
- Schaller, G.B. The Serengeti Lion. A Study of Predator-Prey Relations; University of Chicago Press: Chicago, IL, USA, 1972. [Google Scholar]
- Hamilton, W.D. Geometry of the selfish herd. J. Theor. Biol. 1971, 31, 295–311. [Google Scholar] [CrossRef]
- Prins, H.H.T.; Iason, G.R. Dangerous lions and nonchalant buffalo. Behaviour 1989, 108, 262–292. [Google Scholar]
- Sianga, K.; Fynn, R.; Bonyongo, M.C. Seasonal habitat selection by African buffalo Syncerus caffer in the Savuti-Mababe–Linyanti ecosystem of northern Botswana. Koedoe 2017, 59, a1382. [Google Scholar] [CrossRef]
- Ogada, M.O.; Woodroffe, R.; Oguge, N.O.; Frank, L.G. Limiting Depredation by African Carnivores: The Role of Livestock Husbandry. Conserv. Biol. 2003, 6, 1–10. [Google Scholar] [CrossRef]
- Woodroffe, R.; Frank, L.G.; Lindsey, P.A.; Ole Ranah, S.M.; Romanach, S. Livestock husbandry as a tool for carnivore conservation in Africa’s community rangelands: A case-control study. Biodivers. Conserv. 2007, 16, 1245–1260. [Google Scholar] [CrossRef]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal evolution during domestication: The domesticated fox as a model. Bioessays 2009, 31, 349–360. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weise, F.J.; Tomeletso, M.; Stein, A.B.; Somers, M.J.; Hayward, M.W. Lions Panthera leo Prefer Killing Certain Cattle Bos taurus Types. Animals 2020, 10, 692. https://doi.org/10.3390/ani10040692
Weise FJ, Tomeletso M, Stein AB, Somers MJ, Hayward MW. Lions Panthera leo Prefer Killing Certain Cattle Bos taurus Types. Animals. 2020; 10(4):692. https://doi.org/10.3390/ani10040692
Chicago/Turabian StyleWeise, Florian J., Mathata Tomeletso, Andrew B. Stein, Michael J. Somers, and Matt W. Hayward. 2020. "Lions Panthera leo Prefer Killing Certain Cattle Bos taurus Types" Animals 10, no. 4: 692. https://doi.org/10.3390/ani10040692