Milk Composition of Asian Elephants (Elephas maximus) in a Natural Environment in Myanmar during Late Lactation

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Population, Animals and Sample Collection
2.1.1. Milk
2.1.2. Forage
2.2. Laboratory Methods
2.2.1. Milk
2.2.2. Forage
2.3. Statistical Analyses
2.3.1. Milk
2.3.2. Forage
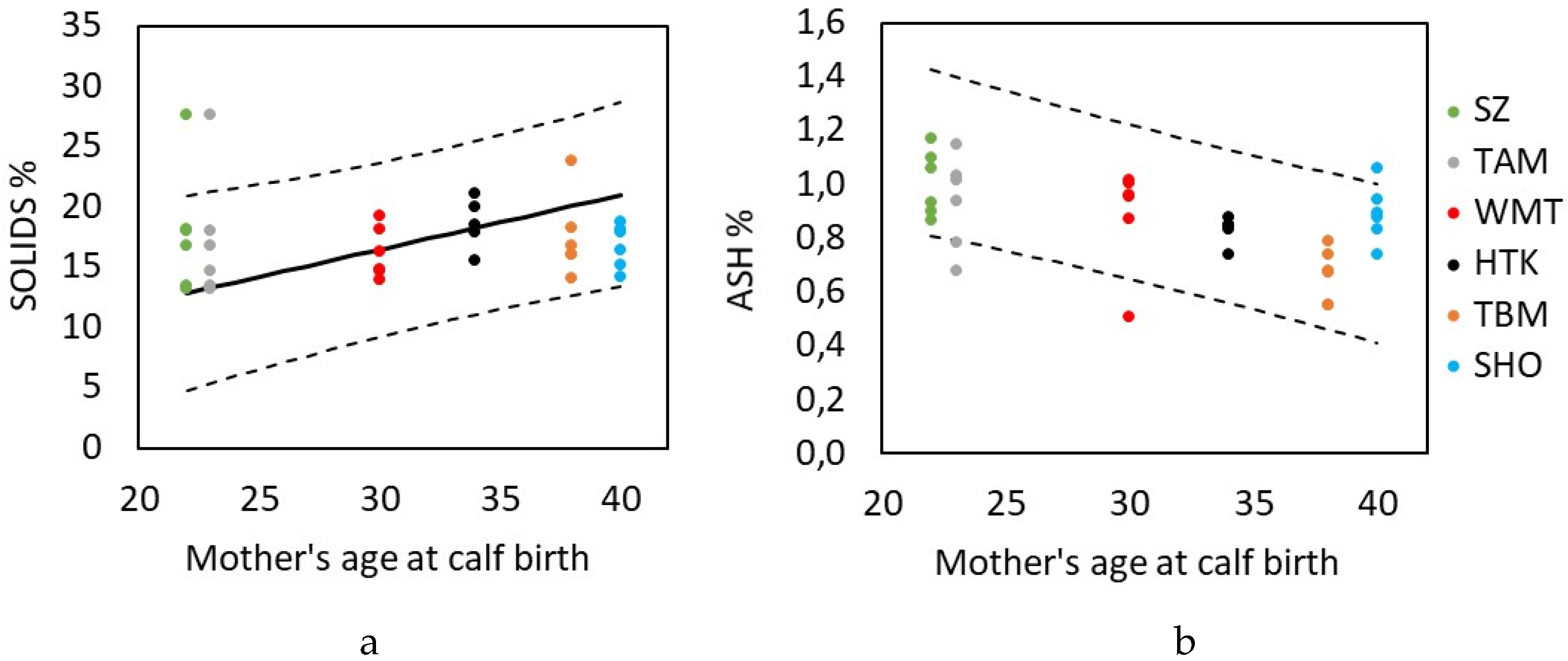
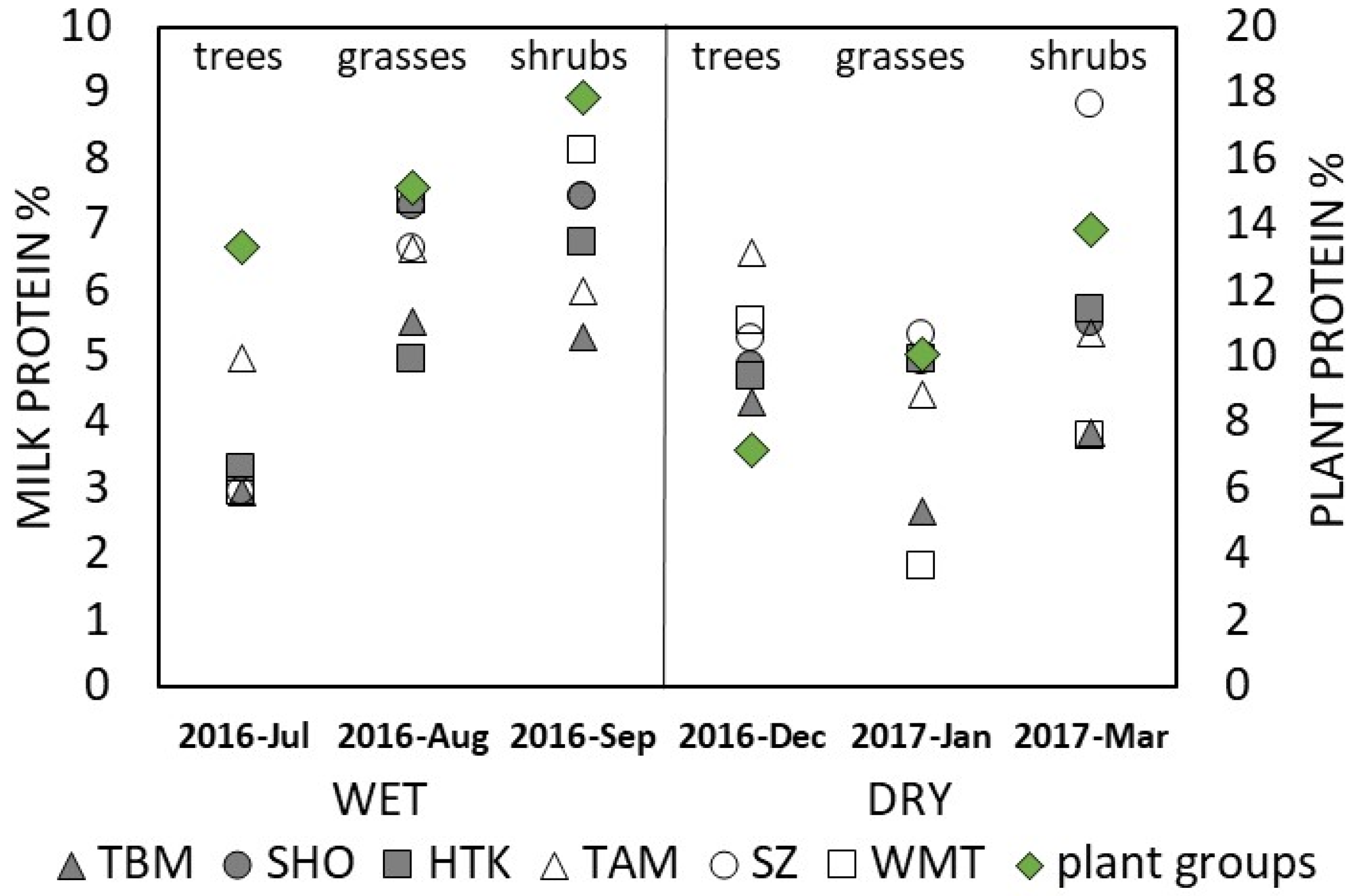
3. Results
3.1. Milk
3.2. Forage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Santiapillai, C. The Asian elephant conservation: A global strategy. Gajah 2010, 18, 21–39. [Google Scholar]
- Sukumar, R. The Living Elephants: Evolutionary Ecology, Behaviour, and Conservation; Oxford University Press: Oxford, UK, 2003; p. 477. [Google Scholar]
- Kurt, F.; Mar, K.U.; Garai, M. Giants in chains: History, biology and preservation of Asian elephants in Asia. In Elephants and Ethics: Toward a Morality of Coexistence; Wemmer, C., Christen, C.A., Eds.; The John Hopkins University Press: Baltimore, MD, USA, 2008; pp. 327–345. [Google Scholar]
- Menon, V.; Tiwari, S.K. Population status of Asian elephants Elephas maximus and key threats. Int. Zoo Yearb. 2019, 53, 17–30. [Google Scholar] [CrossRef]
- Sukumar, R. A brief review of the status, distribution and biology of wild Asian elephants Elephas maximus. Int. Zoo Yearb. 2006, 40, 1–8. [Google Scholar] [CrossRef]
- Hedges, S.; Leimgruber, P.; Lynam, A.; Mar, K.U.; Riddle, H.; Thaw, U.W.N.; Tyson, M. Myanmar Elephant Conservation Action Plan (MECAP): 2018–2027; Forest Department, Ministry of Natural Resources and Environmental Conservation: Nay Pyi Taw, Myanmar, 2018; p. 88. [Google Scholar]
- Jackson, J.; Childs, D.Z.; Mar, K.U.; Htut, W.; Lummaa, V. Long-term trends in wild-capture and population dynamics point to an uncertain future for captive elephants. Proc. R. Soc. B Biol. Sci. 2019, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.A.M.; Dierenfeld, E.S.; Mar, K.U.; Lahdenperä, M.; Lummaa, V.; Aung, A. A simple, practical method for measurement of fat in milk, applied to mid- to late-lactating working elephants in Myanmar. Nov. Tech. Nutr. Food Sci. 2018, 2. [Google Scholar] [CrossRef]
- Clubb, R.; Rowcliffe, M.; Lee, P.; Mar, K.U.; Moss, C.; Mason, G.J. Compromised survivorship in zoo elephants. Science 2008, 322, 1649. [Google Scholar] [CrossRef] [Green Version]
- Mar, K.U.; Lahdenperä, M.; Lummaa, V. Causes and correlates of calf mortality in captive Asian elephants (Elephas maximus). PLoS ONE 2012, 7, e32335. [Google Scholar] [CrossRef]
- Moss, C.J.; Croze, H.; Lee, P.C. The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; The University of Chicago Press: Chicago, IL, USA, 2011. [Google Scholar]
- Lee, P.C.; Moss, C.J. Early maternal investment in male and female African elephant calves. Behav. Ecol. Sociobiol. 1986, 18, 353–361. [Google Scholar] [CrossRef]
- Lindsay, W.K. Feeding Ecology and Population Demography of African Elephants in Amboseli, Kenya. Ph.D. Thesis, University of Cambridge, Cambridge, UK, 1994. [Google Scholar]
- Clubb, R.; Mason, G. A Review of the Welfare of Zoo Elephants in Europe; RSPCA: Horsham, UK, 2002; p. 303. [Google Scholar]
- Boonprasert, K.; Punyapornwithaya, V.; Tankaew, P.; Angkawanish, T.; Sriphiboon, S.; Titharam, C.; Brown, J.L.; Somgird, C. Survival analysis of confirmed elephant endotheliotropic herpes virus cases in Thailand from 2006–2018. PLoS ONE 2019, 14, e0219288. [Google Scholar] [CrossRef]
- Crawley, J.A.H.; Lahdenperä, M.; Oo, Z.M.; Htut, W.; Nandar, H.; Lummaa, V. Taming age mortality in semi-captive Asian elephants. Sci. Rep. 2020, 10, 1889. [Google Scholar] [CrossRef]
- Lahdenperä, M.; Mar, K.U.; Lummaa, V. Nearby grandmothers enhances calf survival and reproduction in Asian elephants. Sci. Rep. 2016, 6, 27213. [Google Scholar] [CrossRef] [PubMed]
- Lahdenperä, M.; Mar, K.U.; Lummaa, V. Short-term and delayed effects of mother death on calf mortality in Asian elephants. Behav. Ecol. 2016, 27, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochs, A.; Hildebrandt, T.B.; Hentschke, J.; Lange, A. Birth and hand rearing of an Asian elephant (Elephas maximus) at Berlin Zoo-Veterinary Experiences. Verh Ber Erkrg Zootiere 2001, 40, 147–156. [Google Scholar]
- Osthoff, G. Elephant milk. In Elephants: Ecology, Behavior and Conservation; Aranovich, M., Dufresne, O., Eds.; Nova Science: Hauppauge, NY, USA, 2012; pp. 97–116. [Google Scholar]
- McCullagh, K.G.; Widdowson, E.M. The milk of the African elephant. Br. J. Nutr. 1970, 24, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Osthoff, G.; De Wit, M.; Hugo, A.; Kamara, B.I. Milk composition of three free-ranging African elephant (Loxodonta africana africana) cows during mid lactation. Comp. Biochem. Physiol. B 2007, 148, 1–5. [Google Scholar] [CrossRef]
- Kunz, C.; Rudloff, S.; Schad, W.; Braun, D. Lactose-derived oligosaccharides in the milk of elephants: Comparison with human milk. Br. J. Nutr. 1999, 82, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Takatsu, Z.; Tsuda, M.; Yamada, A.; Matsumoto, H.; Takai, A.; Takeda, Y.; Takase, M. Elephant’s breast milk contains large amounts of glucosamine. J. Vet. Med. Sci. 2017, 79, 524–533. [Google Scholar] [CrossRef]
- Peters, J.M.; Maire, R.; Hawthorne, B.E.; Storvick, C.A. Composition and nutrient content of elephant (Elephas maximus) milk. J. Mammal. 1972, 53, 717–724. [Google Scholar] [CrossRef]
- Mainka, S.A.; Cooper, R.M.; Black, S.R.; Dierenfeld, E.S. Asian elephant (Elephas maximus) milk composition during the first 280 days of lactation. Zoo Biol. 1994, 13, 389–393. [Google Scholar] [CrossRef]
- Parrott, J.J. Analysis of African elephant mature milk in early lactation and formulation of an elephant calf milk replacer. In Annual Conference Proceedings; American Association of Zoo Veterinarians: Puerto Vallarta, Mexico, 1996; pp. 102–111. [Google Scholar]
- Abbondanza, F.N.; Power, M.L.; Dickson, M.A.; Brown, J.; Oftedal, O.T. Variation in the composition of milk of Asian elephants (Elephas maximus) throughout lactation. Zoo Biol. 2013, 32, 291–298. [Google Scholar] [CrossRef]
- Noori, M.; Ranjbar, S.; Nazari, R.; Jafari, M. The relationship between feed composition and milk qualification in some Iranian grazieries. Int. J. Food Sci. Nutr. Eng. 2014, 4, 55–65. [Google Scholar]
- Tripathi, M.K. Effect of nutrition on production, composition, fatty acids and nutraceutical properties of milk. J. Adv. Dairy Res. 2014, 2, 1–11. [Google Scholar]
- Tyasi, T.L.; Gxasheka, M.; Tlabela, C.P. Assessing the effect of nutrition on milk composition of dairy cows: A review. Int. J. Curr. Sci. 2015, 17, 56–63. [Google Scholar]
- Sutton, J.D. Altering milk composition by feeding. J. Dairy Sci. 1989, 72, 2801–2814. [Google Scholar] [CrossRef]
- Jelen, P.; Lutz, S. Functional milk and dairy products. In Functional Foods: Biochemical and Processing Aspects; Mazza, G., Ed.; Technomic Publishing Company: Lancaster, PA, USA, 1998; pp. 357–380. [Google Scholar]
- Hoffman, R.M.; Kronfeld, D.S.; Herbein, J.H.; Swecker, W.S.; Cooper, W.L.; Harris, P.A. Dietary carbohydrates and fat influence milk composition and fatty acid profile of mare’s milk. J. Nutr. 1998, 128, 2708S–2711S. [Google Scholar] [CrossRef]
- Hinde, K.; Carpenter, A.J.; Clay, J.S.; Bradford, B.J. Holsteins favor heifers, not bulls: Biased milk production programmed during pregnancy as a function of fetal sex. PLoS ONE 2014, 9, e86169. [Google Scholar] [CrossRef]
- Mumby, H.S.; Courtiol, A.; Mar, K.U.; Lummaa, V. Climatic variation and age-specific survival in Asian elephants from Myanmar. Ecology 2013, 94, 1131–1141. [Google Scholar] [CrossRef]
- Mumby, H.S.; Mar, K.U.; Thitaram, C.; Courtiol, A.; Towiboon, P.; Min-Oo, Z.; Htut-Aung, Y.; Brown, J.L.; Lummaa, V. Stress and body condition are associated with climate and demography in Asian elephants. Conserv. Physiol. 2015, 3, cov030. [Google Scholar] [CrossRef] [Green Version]
- Reichert, S.; Berger, V.; Jackson, J.; Chapman, S.N.; Htut, W.; Mar, K.U.; Lummaa, V. Maternal age at birth shapes offspring life-history trajectory across generations in long-lived Asian elephants. J. Anim. Ecol. 2019. [Google Scholar] [CrossRef]
- Lahdenperä, M.; Mar, K.U.; Courtiol, A.; Lummaa, V. Differences in age-specific mortality between wild-caught and captive-born Asian elephants. Nat. Commun. 2018, 9, 3023. [Google Scholar] [CrossRef]
- Lahdenperä, M.; Jackson, J.; Htut, W.; Lummaa, V. Capture from the wild has long-term costs on reproductive success in Asian elephants. Proc. R. Soc. B 2019, 286. [Google Scholar] [CrossRef]
- Chapman, S.N.; Mumby, H.S.; Crawley, J.A.H.; Mar, K.U.; Htut, W.; Soe, A.T.; Aung, H.H.; Lummaa, V. How big is it really? Assessing the efficacy of indirect estimates of body size in Asian elephants. PLoS ONE 2016, 11, e0150533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaw, K. Utilization of elephants in timber harvesting in Myanmar. Gajah 1997, 17, 9–22. [Google Scholar]
- Lynch, E.; Lummaa, V.; Htut, W.; Lahdenperä, M. Evolutionary significance of maternal kinship in a long-lived mammal. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumby, H.S.; Courtiol, A.; Mar, K.U.; Lummaa, V. Birth seasonality and calf mortality in a large population of Asian elephants. Ecol. Evol. 2013, 11, 3794–3803. [Google Scholar] [CrossRef] [PubMed]
- Dierenfeld, E.S.; Dolensek, E.P. Circulating levels of vitamin E in captive Asian elephants (Elephas maximus). Zoo Biol. 1988, 7, 165–172. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Fleet, I.R.; Linzell, J.L. A rapid method of estimating fat in very small quantities of milk. J. Physiol. 1964, 175, 15–17. [Google Scholar]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analyses (Apparatus, Reagents, Procedures and Some Applications); Agricultural Research Service Handbook No. 379; USDA: Washington, DC, USA, 1970; p. 20.
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-Plus; Springer: New York, NY, USA, 2000; ISBN 0-387-98957-0. [Google Scholar]
- Sukumar, R. Ecology of the Asian elephant in southern India. I. Movement and habitat utilization patterns. J. Trop. Ecol. 1989, 5, 1–18. [Google Scholar] [CrossRef]
- Walker, G.P.; Dunshea, F.R.; Doyle, P.T. Effects of nutrition and management on the production and composition of milk fat and protein: A review. Aust. J. Agric. Res. 2004, 55, 1009–1028. [Google Scholar] [CrossRef]
- Glade, M.J. Dietary yeast culture supplementation of mares during late gestation and early lactation: Effects on milk production, milk composition, weight gain and linear growth of nursing foals. J. Equine Veter Sci. 1991, 11, 89–95. [Google Scholar] [CrossRef]
- Ullrey, D.E.; Crissey, S.D.; Hintz, H.F. Elephants: Nutrition and Dietary Husbandry; Nutrition Advisory Group: East Lansing, MI, USA, 1997; pp. 1–20. Available online: www.nagonline.net (accessed on 15 January 2020).
- Reitkirk, F.E.; Hiddingh, H.; Van Dijk, S. Hand-rearing an Asian elephant (Elephas maximus) at the Noorder Zoo, Emmen. Int. Zoo Yearb. 1993, 32, 244–252. [Google Scholar] [CrossRef]
- Grummer, R.R. Effect of feed on the composition of milk fat. J. Dairy Sci. 1991, 74, 3244–3257. [Google Scholar] [CrossRef]
- Sawal, R.K.; Kurar, C.K. Milk yield and its fat content as affected by dietary factors: A review. Asian Australas. J. Anim. Sci. 1997, 11, 217–233. [Google Scholar] [CrossRef]
- Wegner, T.N.; Stull, J.W. Relation between mastitis test score, mineral composition of milk, and blood electrolyte profiles in Holstein cows. J. Dairy Sci. 1978, 61, 1755–1759. [Google Scholar] [CrossRef]
- Landee-Castillejos, T.; Garcia, A.; Lopez-Serrano, F.R.; Gallego, L. Maternal quality and differences in milk production and composition for male and female Iberian red deer calves (Cervus elaphus hispanicus). Behav. Ecol. Sociobiol. 2005, 57, 267–274. [Google Scholar] [CrossRef]
- Hinde, K. First time macaque mothers bias milk composition in favor of sons. Curr. Biol. 2007, 17, 958–959. [Google Scholar] [CrossRef] [Green Version]
- Hinde, K. Richer milk for sons but more milk for daughters: Sex-biased investment during lactation varies with maternal life history in rhesus macaques. Am. J. Hum. Biol. 2009, 21, 512–519. [Google Scholar] [CrossRef]
- Robert, K.A.; Braun, S. Milk composition during lactation suggests a mechanism for male biased allocation of maternal resources in the tammar wallaby (Macropus eugenii). PLoS ONE 2012, 7, e51099. [Google Scholar] [CrossRef]
- Jensen, R.G. (Ed.) Handbook of Milk Composition; Academic Press: New York, NY, USA, 1995; p. 117. [Google Scholar]
- Bakheit, S.A.; Majid, A.M.A.; Nikhala, A.M.M.A. Camels (Camelus dromedarius) under pastoral systems in North Kordofan, Sudan: Seasonal and parity effects upon milk composition. J. Camelid Sci. 2008, 1, 32–36. [Google Scholar]
- Carnicella, D.; Dario, M.; Ayres, M.C.C.; Laudadio, V.; Dario, C. The effect of diet, parity, year and number of kids on milk yield and milk composition in Maltese goat. Small Rumin. Res. 2008, 77, 71–74. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Q.; Yi, M.; Pang, Z.H.; Xiong, B.H. Effects of seasonal change and parity on raw milk composition and related indices in Chinese Holstein cows in northern China. J. Dairy Sci. 2013, 96, 6863–6869. [Google Scholar] [CrossRef] [PubMed]
- Sevi, A.; Taibi, L.; Albenzio, M.; Muscio, A.; Annicchiarico, G. Effect of parity on milk yield, composition, somatic cell count, renneting parameters, and bacteria counts of Comisana ewes. Small Rumin. Res. 2000, 37, 99–107. [Google Scholar] [CrossRef]
- Doreau, M.; Boulot, S.; Martin-Rosset, W. Effect of parity and physiological state on intake, milk production, and blood parameters in lactating mares differing in body size. Anim. Sci. 1999, 53, 111–118. [Google Scholar] [CrossRef]
- Himmelsbach, W.; González Tagle, M.A.; Fueldner, K.; Hoefle, H.H.; Htun, W. Food plants of captive elephants in the Okkan Reserved Forest, Myanmar (Burma), Southeast Asia. Ecotropica 2006, 12, 15–26. [Google Scholar]
- Campos-Arceiz, A.; Lin, T.Z.; Htun, W.; Takatsuki, S.; Leimgruber, P. Working with mahouts to explore the diet of work elephants in Myanmar (Burma). Ecol. Res. 2008, 23, 1057–1064. [Google Scholar] [CrossRef]
- Knowles, S.O.; Grace, N.D.; Knight, T.W.; McNabb, W.C.; Lee, J. Reasons and means for manipulating the micronutrient composition of milk from grazing dairy cattle. Anim. Feed. Sci. Technol. 2006, 131, 165–167. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Elephant | Age at Birth (Years) | Origin | Parity No. | Body Weight (kg) | Date of Calf Birth | Sex of Calf | Months of Lactation Sampled |
|---|---|---|---|---|---|---|---|
| SZ | 22 | C | 2 | 2404 (B) | 1.7.2014 | M | 24–32 |
| TAM | 23 | C | 4 | 2582 (B) | 15.1.2014 | M | 30–38 |
| WMT | 30 | W | 2 | 2706 (B) | 22.9.2014 | M | 22–30 |
| HTK | 34 | C | 2 | 2407 (S) | 26.2.2015 | M | 17–25 |
| TBM | 38 | C | 5 | 2964 (B) | 22.2.2015 | F | 17–25 |
| SHO | 40 | W | 4 | 2358 (S) | 1.1.2015 | F | 18–26 |
| Local Name | Botanical Name | Type of Plant | Common Name | Part Eaten |
|---|---|---|---|---|
| Tin-War | Cephalostachum pergracile | Grass(Tree) | Bamboo | Leaf |
| Kaung-Si-Nwel | UNKNOWN | Shrub | Entire plant | |
| Khway-Ei-Poke-Nwel | Sarcococca pruniformis | Shrub | Fleshy berry | Entire plant |
| Wabo-War | Dendrocalamus brandisii | Grass(Tree) | Bamboo | Leaf |
| Kha-Oung | Ficus hispida | Tree | Hairy Fig | Leaf |
| Pang-Zauk-Htoe | UNKNOWN | Shrub | Entire plant | |
| Tama-saing | Thysanolaena maxima | Grass | Tiger Grass | Leaf |
| Ka-Pyin-Nwel | Quercus fenestrata | Tree | Oak Tree | Leaf |
| Dant-Kywe | Cassia sophera | Shrub | Senna Sophera | Leaf |
| Wa-Phyu-War | Dendrocalamus membranaceus | Grass(Tree) | Bamboo | Leaf |
| Constituent | Mean ± SD | Range |
|---|---|---|
| Water, % | 82.44 ± 3.65 | (72.28–86.74) |
| Total Solids, % | 17.56 ± 3.65 | (13.26–27.72) |
| Total Fat, % | 15.10 ± 3.87 1 | (7.59–23.48) |
| Liquid fat layer, % of total fat | 79.19 ± 12.44 | (38.77–95.75) |
| Cream fat layer, % of total fat | 20.81 ± 12.44 | (4.25–61.23) |
| Crude Protein, % | 5.23 ± 1.66 | (1.82–8.81) |
| Ash, % | 0.87 ± 0.16 | (0.51–1.17) |
| Vitamin E (µg/mL) | 0.18 ± 0.014 | (0.16–0.21) |
| Species | CP, % | CP, % | NDF, % | NDF, % | ADF, % | ADF, % | Ash, % | Ash, % |
|---|---|---|---|---|---|---|---|---|
| Season | Dry | Wet | Dry | Wet | Dry | Wet | Dry | Wet |
| Wabo-war Dendrocalamus brandisii | 12.70 | 16.72 | 66.92 | 67.60 | 41.21 | 37.88 | 11.55 | 10.60 |
| Tin-war Cephalostachum pergracile | 8.82 | 11.26 | 59.49 | 67.18 | 46.55 | 39.45 | 21.65 | 16.80 |
| Wa-phyu-war Dendrocalamus membranaceus | 10.95 | 19.39 | 58.56 | 68.76 | 40.31 | 30.36 | 16.76 | 10.16 |
| Tama-saing Thysanolaena maxima | 7.75 | 13.08 | 74.67 | 72.08 | 45.80 | 38.60 | 10.36 | 11.15 |
| Khway-ei-poke-nwel Sarcococca pruniformis | 8.61 | 10.61 | 46.92 | 52.69 | 38.47 | 45.86 | 10.50 | 14.23 |
| Ka-pyin-nwel Quercus fenestrate | 6.03 | 13.18 | 61.35 | 32.27 | 51.21 | 26.60 | 6.14 | 8.67 |
| Dant-kywe Cassia sophera | 18.59 | 22.39 | 37.59 | 28.60 | 34.60 | 19.51 | 15.32 | 13.79 |
| Kaung-si-nwel | 12.06 | 25.34 | 38.50 | 43.03 | 33.48 | 42.36 | 15.67 | 17.09 |
| Pan-zauk-htoe | 16.21 | 13.03 | 34.79 | 40.29 | 34.94 | 35.65 | 19.90 | 23.26 |
| Kha-oung Ficus hispida | 8.35 | 13.54 | 40.74 | 40.80 | 37.75 | 32.43 | 21.53 | 20.05 |
| Mean | ** 11.01 | 15.86 | 51.95 | 51.33 | 40.43 | 34.87 | 14.94 | 14.58 |
| ±SD | 3.97 | 4.97 | 13.99 | 16.44 | 5.84 | 7.83 | 5.23 | 4.71 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dierenfeld, E.S.; Han, Y.A.M.; Mar, K.U.; Aung, A.; Soe, A.T.; Lummaa, V.; Lahdenperä, M. Milk Composition of Asian Elephants (Elephas maximus) in a Natural Environment in Myanmar during Late Lactation. Animals 2020, 10, 725. https://doi.org/10.3390/ani10040725
Dierenfeld ES, Han YAM, Mar KU, Aung A, Soe AT, Lummaa V, Lahdenperä M. Milk Composition of Asian Elephants (Elephas maximus) in a Natural Environment in Myanmar during Late Lactation. Animals. 2020; 10(4):725. https://doi.org/10.3390/ani10040725
Chicago/Turabian StyleDierenfeld, Ellen S., Yadana A. M. Han, Khyne U. Mar, Aung Aung, Aung Thura Soe, Virpi Lummaa, and Mirkka Lahdenperä. 2020. "Milk Composition of Asian Elephants (Elephas maximus) in a Natural Environment in Myanmar during Late Lactation" Animals 10, no. 4: 725. https://doi.org/10.3390/ani10040725
APA StyleDierenfeld, E. S., Han, Y. A. M., Mar, K. U., Aung, A., Soe, A. T., Lummaa, V., & Lahdenperä, M. (2020). Milk Composition of Asian Elephants (Elephas maximus) in a Natural Environment in Myanmar during Late Lactation. Animals, 10(4), 725. https://doi.org/10.3390/ani10040725