Ontogeny of Expression and Activity of Digestive Enzymes and Establishment of gh/igf1 Axis in the Omnivorous Fish Chelon labrosus

 , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Rearing and Sampling Protocol
2.2. Cloning and Gene Expression Analyses
2.2.1. Molecular Cloning
2.2.2. Gene Quantification Using Quantitative Real-Time PCR
2.3. Enzyme Activity Analyses
2.4. Statistical Analysis
3. Results
3.1. Growth
3.2. Molecular Sequencing
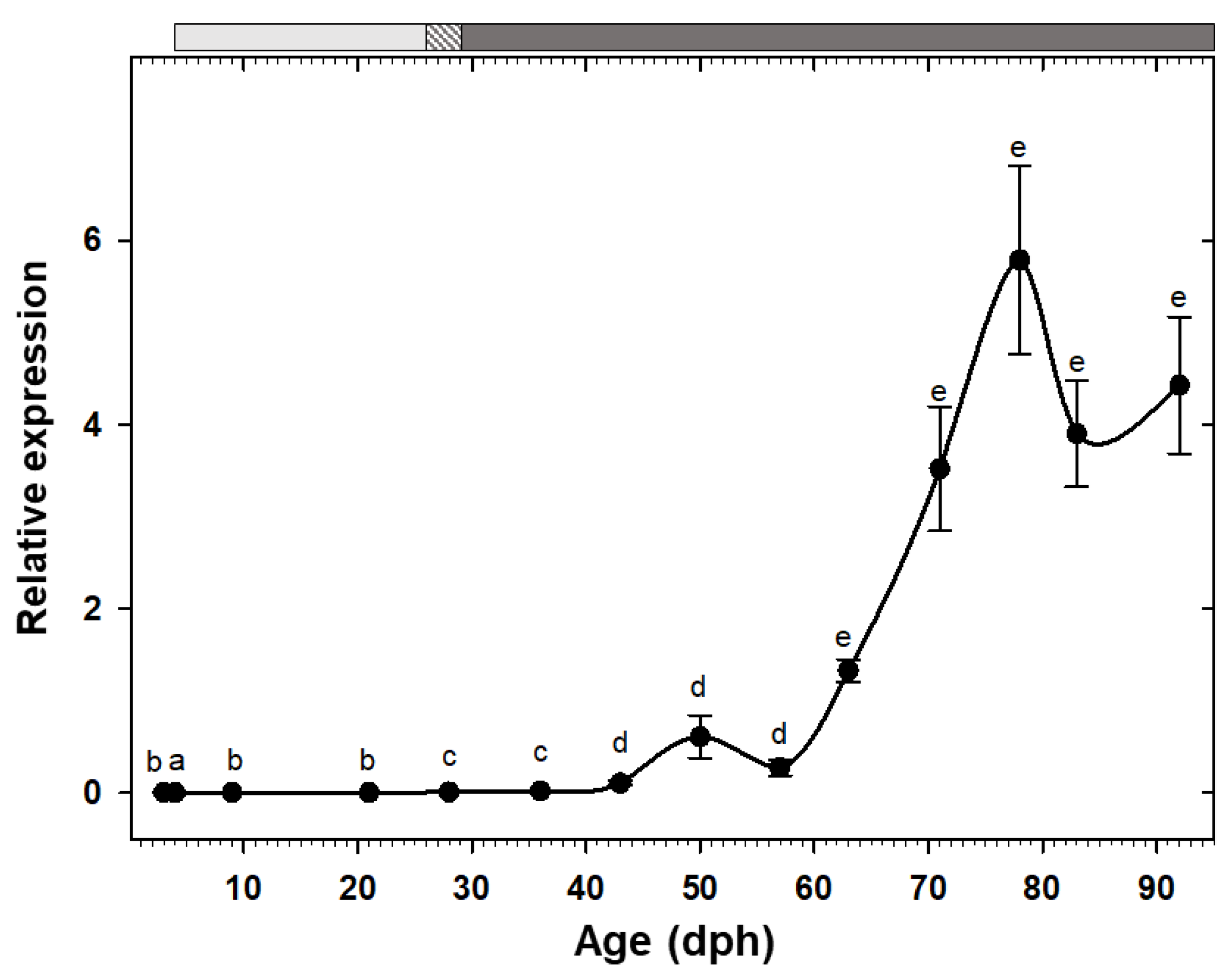
3.3. Gene Expression
3.4. Enzyme Activity Analyses
4. Discussion
4.1. Growth
4.2. Sequences
4.3. Ontogeny of Digestive Function
4.3.1. Protein Digestion
4.3.2. Lipid Digestion
4.3.3. Carbohydrate Digestion
4.4. Ontogeny of gh/igf1 Axis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| actb | actin beta |
| amy2a | pancreatic alpha amylase |
| atp4a | gastric H+/K+-ATPase |
| cel | carboxyl ester lipase precursor |
| ctr | chymotrypsinogen precursor |
| gh | growth hormone |
| igf1 | insulin-like growth factor 1 |
| pga2 | pepsinogen 2 precursor |
| pla2g1b | pancreatic phospholipase A2 |
| try2 | trypsinogen 2 precursor |
| aa | amino acid |
| Ct | cycle threshold |
| dph | days post hatch |
| Gh | growth hormone |
| GIT | gastrointestinal tract |
| Igf1 | insulin-like growth factor 1 |
| ORF | open reading frame |
| qPCR | quantitative real-time PCR |
| SEM | standard error of the mean |
References
- Ben Khemis, I.; Zouiten, D.; Besbes, R.; Kamoun, F. Larval rearing and weaning of thick lipped grey mullet (Chelon labrosus) in mesocosm with semiextensive technology. Aquaculture 2006, 259, 190–201. [Google Scholar] [CrossRef]
- Ben Khemis, I.; Gisbert, E.; Alcaraz, C.; Zouiten, D.; Besbes, R.; Zouiten, A.; Masmoudi, A.S.; Cahu, C. Allometric growth patterns and development in larvae and juveniles of thick-lipped grey mullet Chelon labrosus reared in mesocosm conditions. Aquac. Res. 2013, 44, 1872–1888. [Google Scholar] [CrossRef] [Green Version]
- Zouiten, D.; Ben Khemis, I.; Besbes, R.; Cahu, C. Ontogeny of the digestive tract of thick lipped grey mullet (Chelon labrosus) larvae in “mesocosms”. Aquaculture 2008, 279, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Delgado, C.; Drake, P.; Arias, A.; García, D. Peces de Doñana y su Entorno; Organismo Autónomo Parques Nacionales—Ministerio de Medio Ambiente: Madrid, Spain, 2000. [Google Scholar]
- Srichanun, M.; Tantikitti, C.; Utarabhand, P.; Kortner, T.M. Gene expression and activity of digestive enzymes during the larval development of Asian seabass (Lates calcarifer). Comp. Biochem. Physiol. B 2013, 165, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yúfera, M.; Martínez-Rodríguez, G.; Moyano, F.J. The Digestive Function in Developing Fish Larvae and Fry. From Molecular Gene Expression to Enzymatic Activity. In Emerging Issues in Fish Larvae Research; Yúfera, M., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 51–86. [Google Scholar]
- Parma, L.; Bonaldo, A.; Massi, P.; Yúfera, M.; Martínez-Rodríguez, G.; Gatta, P.P. Different early weaning protocols in common sole (Solea solea L.) larvae: Implications on the performances and molecular ontogeny of digestive enzyme precursors. Aquaculture 2013, 414–415, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Galaviz, M.A.; García-Ortega, A.; Gisbert, E.; López, L.M.; Gasca, A.G. Expression and activity of trypsin and pepsin during larval development of the spotted rose snapper Lutjanus guttatus. Comp. Biochem. Physiol. B 2012, 161, 9–16. [Google Scholar] [CrossRef]
- Khoa, T.; Waqalevu, V.; Honda, A.; Shiozaki, K.; Kotani, T. Early ontogenetic development, digestive enzymatic activity and gene expression in red sea bream (Pagrus major). Aquaculture 2019, 512, 734283. [Google Scholar] [CrossRef]
- Sarasquete, C.; Ortiz-Delgado, J.B.; Martos-Sitcha, J.A.; de las Heras, V.; Yúfera, M.; Martínez-Rodríguez, G. Ontogeny and functional histochemistry of the digestive and visual systems and other organs during the larval development of the thick-lipped grey mullet, Chelon labrosus. Sci. Mar. 2014, 78, 473–491. [Google Scholar] [CrossRef] [Green Version]
- Picha, M.E.; Yurano, M.J.; Beckman, B.R.; Borski, R.J. Endocrine biomarkers of growth and applications to aquaculture: A minireview of growth hormone, insulin-like growth factor (IGF)-I, and IGF-binding proteins as potential growth indicators in fish. N. Am. J. Aquacult. 2008, 70, 196–211. [Google Scholar] [CrossRef]
- Deane, E.E.; Kelly, S.P.; Collins, P.M.; Woo, N.Y. Larval development of silver sea bream (Sparus sarba): Ontogeny of RNA–DNA ratio, GH, IGF-I, and Na+–K+-ATPase. Mar. Biotechnol. 2003, 5, 79–91. [Google Scholar] [CrossRef]
- Miandare, H.K.; Farahmand, H.; Akbarzadeh, A.; Ramezanpour, S.; Kaiya, H.; Miyazato, M.; Rytkönen, K.T.; Nikinmaa, M. Developmental transcription of genes putatively associated with growth in two sturgeon species of different growth rate. Gen. Comp. Endocrinol. 2013, 182, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.Y.; Greene, M.; Chen, T.T. Early embryonic expression of the growth hormone family protein genes in the developing rainbow trout, Oncorhynchus mykiss. Mol. Reprod. Dev. 1999, 53, 127–134. [Google Scholar] [CrossRef]
- Funkenstein, B.; Almuly, R.; Chan, S.J. Localization of IGF-I and IGF-I Receptor mRNA in Sparus aurata Larvae. Gen. Comp. Endocrinol. 1997, 107, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Perrot, V.; Moiseeva, E.B.; Gozes, Y.; Chan, S.J.; Ingleton, P.; Funkenstein, B. Ontogeny of the insulin-like growth factor system (IGF-I, IGF-II, and IGF-1R) in gilthead seabream (Sparus aurata): Expression and cellular localization. Gen. Comp. Endocrinol. 1999, 116, 445–460. [Google Scholar] [CrossRef]
- Gilannejad, N.; Paykan Heyrati, F.; Dorafshan, S.; Martos-Sitcha, J.; Yúfera, M.; Martínez-Rodríguez, G. Molecular basis of the digestive functionality in developing Persian sturgeon (Acipenser persicus) larvae: Additional clues for its phylogenetic status. J. Comp. Physiol. B 2019, 189, 367–383. [Google Scholar] [CrossRef]
- Pujante, I.M.; Martos-Sitcha, J.A.; Moyano, F.J.; Ruiz-Jarabo, I.; Martínez-Rodríguez, G.; Mancera, J.M. Starving/re-feeding processes induce metabolic modifications in thick-lipped greymullet (Chelon labrosus, Risso 1827). Comp. Biochem. Physiol. B 2015, 180, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pujante, I.M.; Moyano, F.J.; Martos-Sitcha, J.A.; Mancera, J.M.; Martínez-Rodríguez, G. Effect of different salinities on gene expression and activity of digestive enzymes in the thick-lipped grey mullet (Chelon labrosus). Fish Physiol. Biochem. 2018, 44, 349–373. [Google Scholar] [CrossRef]
- Rotllant, G.; Moyano, F.; Andrés, M.; Díaz, M.; Estévez, A.; Gisbert, E. Evaluation of fluorogenic substrates in the assessment of digestive enzymes in a decapod crustacean Maja brachydactyla larvae. Aquaculture 2008, 282, 90–96. [Google Scholar] [CrossRef]
- Finn, R.; Attwood, T.; Babbitt, P.; Bateman, A.; Bork, P.; Bridge, A.; Chang, H.Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017—beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef]
- Rønnestad, I.; Yúfera, M.; Ueberschär, B.; Ribeiro, L.; Sæle, Ø.; Boglione, C. Feeding behaviour and digestive physiology in larval fish: Current knowledge, and gaps and bottlenecks in research. Rev. Aquacult. 2013, 5 (Suppl. 1), S59–S98. [Google Scholar] [CrossRef]
- Zambonino-Infante, J.L.; Cahu, C.L. Ontogeny of the gastrointestinal tract of marine fish larvae. Comp. Biochem. Physiol. C 2001, 130, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Yúfera, M.; Darias, M.J. Changes in the gastrointestinal pH from larvae to adult in Senegal sole (Solea senegalensis). Aquaculture 2007, 267, 94–99. [Google Scholar] [CrossRef]
- Darias, M.J.; Zambonino-Infante, J.; Hugot, K.; Cahu, C.; Mazurais, D. Gene expression patterns during the larval development of European seabass (Dicentrarchus labrax) by microarray analysis. Mar. Biotechnol. 2008, 10, 416–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortner, T.M.; Overrein, I.; Øie, G.; Kjørsvik, E.; Bardal, T.; Wold, P.A.; Arukwe, A. Molecular ontogenesis of digestive capability and associated endocrine control in Atlantic cod (Gadus morhua) larvae. Comp. Biochem. Physiol. A 2011, 160, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Moguel-Hernández, I.; Peña, R.; Andree, K.; Tovar-Ramirez, D.; Bonacic, K.; Dumas, S.; Gisbert, E. Ontogeny changes and weaning effects in gene expression patterns of digestive enzymes and regulatory digestive factors in spotted rose snapper (Lutjanus guttatus) larvae. Fish Physiol. Biochem. 2016, 42, 1319–1334. [Google Scholar] [CrossRef]
- Péres, A.; Zambonino-Infante, J.; Cahu, C. Dietary regulation of activities and mRNA levels of trypsin and amylase in sea bass (Dicentrarchus labrax) larvae. Fish Physiol. Biochem. 1998, 19, 145–152. [Google Scholar] [CrossRef]
- Pujante, I.M.; Díaz-López, M.; Mancera, J.M.; Moyano, F.J. Characterization of digestive enzymes protease and alpha-amylase activities in the thick-lipped grey mullet (Chelon labrosus, Risso 1827). Aquac. Res. 2017, 48, 367–376. [Google Scholar] [CrossRef]
- Douglas, S.E.; Gawlicka, A.; Mandla, S.; Gallant, J.W. Ontogeny of the stomach in winter flounder: Characterization and expression of the pepsinogen and proton pump genes and determination of pepsin activity. J. Fish Biol. 1999, 55, 897–915. [Google Scholar] [CrossRef]
- Feng, S.Z.; Li, W.S.; Lin, H.R. Characterization and expression of the pepsinogen C gene and determination of pepsinlike enzyme activity from orange-spotted grouper (Epinephelus coioides). Comp. Biochem. Physiol. 2008, 55, 897–915. [Google Scholar]
- Wu, X.; Washio, Y.; Aritaki, M.; Fujinami, Y.; Shimizu, D.; Hashimoto, H.; Iwasaki, T.; Uji, S.; Suzuki, T. Staging of initial pepsinogen and chitinase expression and complete gastric gland development within the larval stomach of Japanese flounder, spotted halibut, seven-band grouper and greater amberjack. Aquaculture 2011, 314, 165–172. [Google Scholar] [CrossRef]
- Koven, W.; Gisbert, E.; Nixon, O.; Solovyev, M.; Gaon, A.; Allon, G.; Meiri-Ashkenazi, I.; Tandler, A.; Rosenfeld, H. The effect of algal turbidity on larval performance and the ontogeny of digestive enzymes in the grey mullet (Mugil cephalus). Comp. Biochem. Physiol. A 2019, 228, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazemroaya, S.; Nematollahi, M.A.; Yazdanparast, R.; Farahmand, H.; Rezaie, A.; Najafabadi, M.Z. Pepsinogen expression during larval development of a Persian Gulf Sparid, Sobaity. Aquaculture 2020, 523, 735131. [Google Scholar] [CrossRef]
- Gilannejad, N. Mechanisms Involved in the Efficiency of Digestive Process in Farmed Marine Fishes. Ph.D. Thesis, University of Cádiz, Cadiz, Spain, 2018. [Google Scholar]
- Teles, A.; Salas-Leiva, J.; Alvarez-González, C.; Tovar-Ramírez, D. Changes in digestive enzyme activities during early ontogeny of Seriola rivoliana. Fish Physiol. Biochem. 2019, 45, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.W.; Folkvord, A.; Grøtan, E.; Sæle, Ø. Genetic ontogeny of pancreatic enzymes in Labrus bergylta larvae and the effect of feed type on enzyme activities and gene expression. Comp. Biochem. Physiol. B 2013, 164, 176–184. [Google Scholar] [CrossRef]
- Hoehne-Reitan, K.; Kjorsvik, E.; Gjellesvik, D.R. Development of bile salt-dependent lipase in larval turbot. J. Fish Biol. 2001, 58, 737–745. [Google Scholar] [CrossRef]
- Perez-Casanova, J.C.; Murray, H.M.; Gallant, J.W.; Ross, N.W.; Douglas, S.E.; Johnson, S.C. Development of the digestive capacity in larvae of haddock (Melanogrammus aeglefinus) and Atlantic cod (Gadus morhua). Aquaculture 2006, 251, 377–401. [Google Scholar] [CrossRef]
- Sæle, Ø.; Nordgreen, A.; Olsvik, P.A.; Hamre, K. Characterization and expression of digestive neutral lipases during ontogeny of Atlantic cod (Gadus morhua). Comp. Biochem. Physiol. A 2010, 157, 252–259. [Google Scholar]
- Mata-Sotres, J.A.; Martos-Sitcha, J.A.; Astola, A.; Yúfera, M.; Martínez-Rodríguez, G. Cloning and molecular ontogeny of digestive enzymes in fed and food-deprived developing gilthead seabream (Sparus aurata) larvae. Comp. Biochem. Physiol. B 2016, 191, 53–65. [Google Scholar] [CrossRef]
- Hoehne-Reitan, K.; Kjørsvik, E.; Reitan, K.I. Lipolytic activities in developing turbot larvae as influenced by diet. Aquacult. Int. 2003, 11, 477–489. [Google Scholar] [CrossRef]
- Morais, S.; Narciso, L.; Dores, E.; Pousao-Ferreira, P. Lipid enrichment for Senegalese sole (Solea senegalensis) larvae: Effect on larval growth, survival and fatty acid profile. Aquac. Int. 2004, 12, 281–298. [Google Scholar] [CrossRef]
- Zambonino-Infante, J.L.; Gisbert, E.; Sarasquete, C.; Navarro, I.; Gutierrez, J.; Cahu, C.L. Ontogeny and physiology of the digestive system of marine fish larvae. In Feeding and Digestive Functions of Fishes; Cyrino, J.E.P., Bureau, D.P., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA, 2008; pp. 281–348. [Google Scholar]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. BBA-Mol. Cell Biol. Lipids 2006, 1761, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Subramaniam, S.; Brown, H.A. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tocher, D.R.; Bendiksen, E.A.; Campbell, P.J.; Bell, J.G. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Falk-Petersen, I.B. Comparative organ differentiation during early life stages of marine fish. Fish Shellfish Immunol. 2005, 19, 397–412. [Google Scholar] [CrossRef]
- Sæle, Ø.; Nordgreen, A.; Olsvik, P.A.; Hamre, K. Characterisation and expression of secretory phospholipase A2 group IB during ontogeny of Atlantic cod (Gadus morhua). Br. J. Nutr. 2011, 105, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Sanz, A.; Llorente, J.I.; Furné, M.; Ostos-Garrido, M.V.; Carmona, R.; Domezain, A.; Hidalgo, M.C. Digestive enzymes during ontogeny of the sturgeon Acipenser naccarii: Intestine and pancreas development. J. Appl. Ichthyol. 2011, 27, 1139–1146. [Google Scholar] [CrossRef]
- Kim, K.H.; Horn, M.H.; Sosa, A.E.; German, D.P. Sequence and expression of an α-amylase gene in four related species of prickleback fishes (Teleostei: Stichaeidae): Ontogenetic, dietary, and species-level effects. J. Comp. Physiol. B 2014, 184, 221–234. [Google Scholar] [CrossRef]
- Darias, M.J.; Murray, H.M.; Gallant, J.W.; Astola, A.; Douglas, S.E.; Yúfera, M.; Martínez-Rodríguez, G. Characterization of a partial α-amylase clone from red porgy (Pagrus pagrus): Expression during larval development. Comp. Biochem. Physiol. B 2006, 143, 209–218. [Google Scholar] [CrossRef]
- Khoa, T.; Waqalevu, V.; Honda, A.; Shiozaki, K.; Kotani, T. Comparative study on early digestive enzyme activity and expression in red sea bream (Pagrus major) fed on live feed and micro-diet. Aquaculture 2020, 519, 734721. [Google Scholar] [CrossRef]
- Péres, A.; Cahu, C.L.; Zambonino-Infante, J.L.; Le Gall, M.M.; Quazuguel, P. Amylase and trypsin responses to intake of dietary carbohydrate and protein depend on the developmental stage in sea bass (Dicentrarchus labrax) larvae. Fish Physiol. Biochem. 1996, 15, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, M.M.; Kashinskaya, E.N.; Izvekova, G.I.; Gisbert, E.; Glupov, V.V. Feeding habits and ontogenic changes in digestive enzyme patterns in five freshwater teleosts. J. Fish Biol. 2016, 85, 1395–1412. [Google Scholar] [CrossRef] [PubMed]
- Li, W.S.; Chen, D.; Wong, A.O.L.; Lin, H.R. Molecular cloning, tissue distribution, and ontogeny of mRNA expression of growth hormone in orange-spotted grouper (Epinephelus coioides). Gen. Comp. Endocrinol. 2005, 144, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Ferraresso, S.; Bonaldo, A.; Parma, L.; Cinotti, S.; Massi, P.; Bargelloni, L.; Gatta, P.P. Exploring the larval transcriptome of the common sole (Solea solea L.). BMC Genom. 2013, 14, 315. [Google Scholar] [CrossRef] [Green Version]
- Ibarra-Castro, L.; Webb, K.A., Jr.; Holt, G.J. Molecular cloning, tissue distribution and ontogenetic expression of growth hormone in cobia, Rachycentron canadum. Rev. Biol. Mar. Oceanog. 2016, 51, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Froesch, E.R.; Schmid, C.; Schwander, J.; Zapf, J. Actions of insulin-like growth factors. Annu. Rev. Physiol. 1985, 47, 443–467. [Google Scholar] [CrossRef]
- Duan, C. The insulin-like growth factor system and its biological actions in fish. Am. Zool. 1997, 37, 491–503. [Google Scholar] [CrossRef] [Green Version]
- de las Heras, V.; Martos-Sitcha, J.A.; Yúfera, M.; Mancera, J.M.; Martínez-Rodríguez, G. Influence of stocking density on growth, metabolism and stress of thick-lipped grey mullet (Chelon labrosus) juveniles. Aquaculture 2015, 448, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Riley, L.G.; Richman, N.H.; Hirano, T.; Grau, E.G. Activation of the growth hormone/insulin-like growth factor axis by treatment with 17amethyltestosterone and seawater rearing in the tilapia, Oreochromis mossambicus. Gen. Comp. Endocrinol. 2002, 127, 285–292. [Google Scholar] [CrossRef]
- Busby, E.R.; Roch, G.J.; Sherwood, N.M. Endocrinology of zebrafish: A small fish with a large gene pool. In Fish Physiology; Perry, S.F., Ekker, M., Farrell, A.P., Brauner, C.J., Eds.; Academic Press/Elsevier: New York, NY, USA, 2010; Volume 29, pp. 173–247. [Google Scholar]
- Hildahl, J.; Power, D.M.; Björnsson, B.T.; Einarsdóttir, I.E. Involvement of growth hormone- insulin-like growth factor I system in cranial remodeling during halibut metamorphosis as indicated by tissue- and stage-specific receptor gene expression and the presence of growth hormone receptor protein. Cell Tissue Res. 2008, 332, 211–225. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Direction | Sequence (5′–3′) | Position 1 | Amplicon Size (bp) | Concentration (nM) | Efficiency | R2 | Amplification Range | Accession Number |
|---|---|---|---|---|---|---|---|---|---|
| Q-actb | Forward | TCTTCCAGCCTTCCTTCCTTG | 863 | 108 | 200 | 1.04 | 0.998 | 5 ng–5 pg | MH350431 |
| Reverse | TGTTGGCGTACAGGTCCTTACGG | 970 | |||||||
| Q-atp4a | Forward | TTGCCTACACGCTAACCAAA | 2027 | 112 | 200 | 0.93 | 0.996 | 5 ng–5 pg | MH350434 |
| Reverse | GCCAGTTCGATGAAGAGGAT | 2138 | |||||||
| Q-pla2 | Forward | ACACCTGTTGATGACCTGGA | 37 | 143 | 200 | 0.94 | 0.997 | 5 ng–5 pg | MH350433 |
| Reverse | GTCTTGGTGGCCTTGTCAC | 179 | |||||||
| Q-cel | Forward | CTGACCATGCTGATGACCTG | 1360 | 101 | 200 | 1 | 0.998 | 5 ng–500 fg | MH350432 |
| Reverse | GGCAATCATGTAACCGGAGA | 1460 | |||||||
| Q-pga2 2 | Forward | AAGATGAAGTGGCTCGTGGTT | - | 122 | 200 | 0.94 | 0.999 | 5 ng–50 pg | KC195968 |
| Reverse | TCTTCCCACAATCCTTTCTCC | - | |||||||
| Q-try2 2 | Forward | CTCCAGAACACAGCCATGAAG | - | 140 | 400 | 0.99 | 0.999 | 5 ng–500 fg | KF684940 |
| Reverse | ACGTTCAGAGAGGCCTGGTAG | - | |||||||
| Q-ctr 2 | Forward | CGTCCCTTCAGGATTATACCG | - | 138 | 400 | 0.97 | 0.998 | 5 ng–500 fg | KC195969 |
| Reverse | AGTTGGAGGAACGGTCATGTT | - | |||||||
| Q-amy2a 2 | Forward | CCAAACTGGGAACTGTCATCAG | - | 129 | 400 | 0.94 | 0.996 | 5 ng–500 fg | KF684941 |
| Reverse | TCTGGTTGTCGTGGTTGTCA | - | |||||||
| Q-gh | Forward | ATCTTCCCTGACGACTCTGC | - | 121 | 200 | 1.03 | 0.998 | 5 ng–50 pg | KC195966 |
| Reverse | GGTATGTCTCCACCTTGTGC | - | |||||||
| Q-igf1 | Forward | GGAACACACAGGTCAAACGA | - | 132 | 200 | 1 | 0.995 | 5 ng–5 pg | KC195967 |
| Reverse | CGCTCCCTTTCTCATAGTTG | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilannejad, N.; de las Heras, V.; Martos-Sitcha, J.A.; Moyano, F.J.; Yúfera, M.; Martínez-Rodríguez, G. Ontogeny of Expression and Activity of Digestive Enzymes and Establishment of gh/igf1 Axis in the Omnivorous Fish Chelon labrosus. Animals 2020, 10, 874. https://doi.org/10.3390/ani10050874
Gilannejad N, de las Heras V, Martos-Sitcha JA, Moyano FJ, Yúfera M, Martínez-Rodríguez G. Ontogeny of Expression and Activity of Digestive Enzymes and Establishment of gh/igf1 Axis in the Omnivorous Fish Chelon labrosus. Animals. 2020; 10(5):874. https://doi.org/10.3390/ani10050874
Chicago/Turabian StyleGilannejad, Neda, Verónica de las Heras, Juan Antonio Martos-Sitcha, Francisco J. Moyano, Manuel Yúfera, and Gonzalo Martínez-Rodríguez. 2020. "Ontogeny of Expression and Activity of Digestive Enzymes and Establishment of gh/igf1 Axis in the Omnivorous Fish Chelon labrosus" Animals 10, no. 5: 874. https://doi.org/10.3390/ani10050874
APA StyleGilannejad, N., de las Heras, V., Martos-Sitcha, J. A., Moyano, F. J., Yúfera, M., & Martínez-Rodríguez, G. (2020). Ontogeny of Expression and Activity of Digestive Enzymes and Establishment of gh/igf1 Axis in the Omnivorous Fish Chelon labrosus. Animals, 10(5), 874. https://doi.org/10.3390/ani10050874