Effects of Drinking Water Temperature and Flow Rate during Cold Season on Growth Performance, Nutrient Digestibility and Cecum Microflora of Weaned Piglets

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Feeding and Diarrhea Occurrence
2.3. Sample Collection
2.4. Apparent Digestibility of Nutrients
2.5. Microflora Analysis
2.6. Statistical Analysis
3. Results
3.1. Growth Performance and Diarrhea Occurrence
3.2. Nutrient Digestibility
3.3. Sequencing, Richness and Diversity of the Bacterial Community
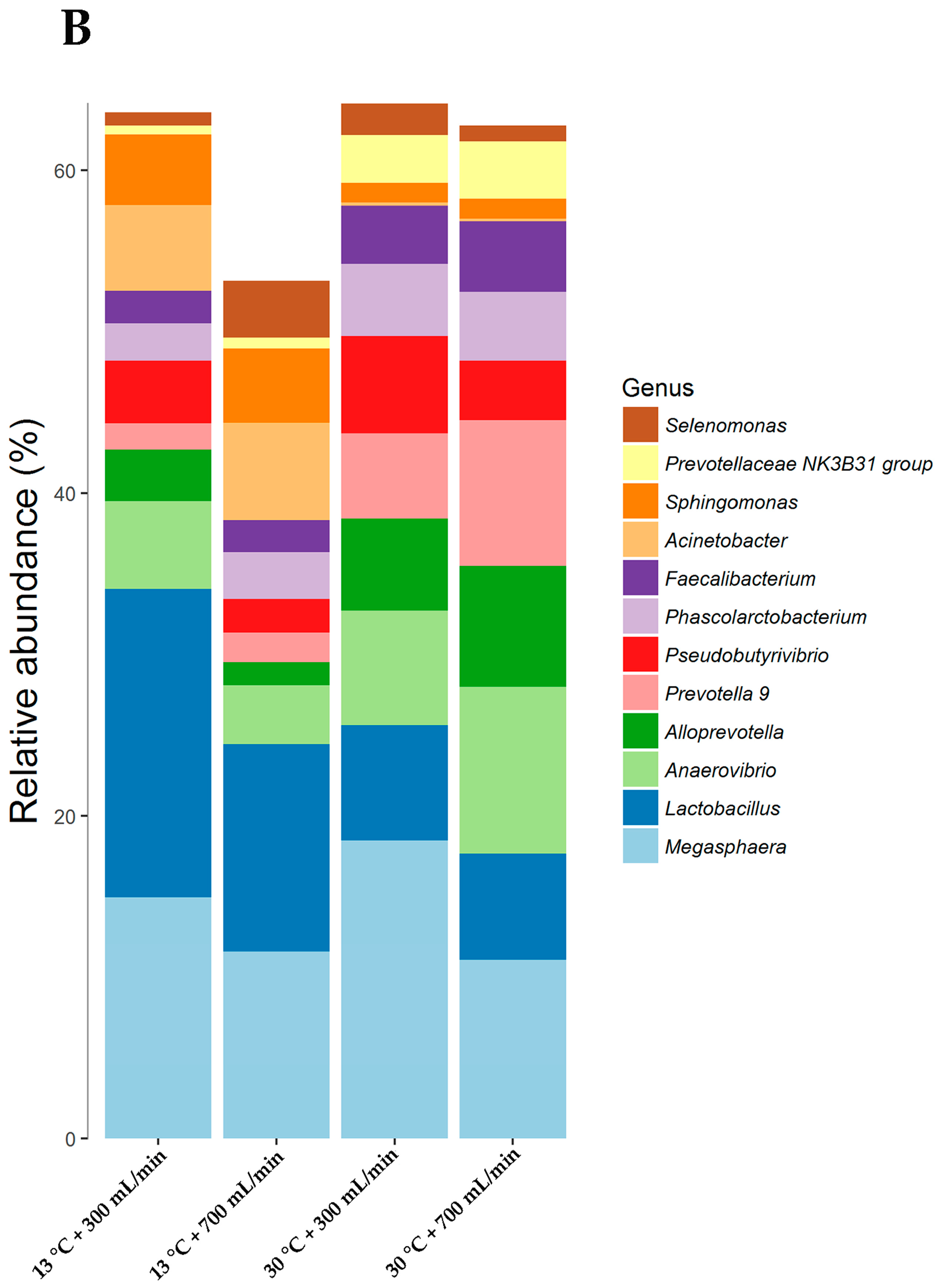
3.4. Microbiome Modulation at Taxonomic Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lybrand, T.P.; Kollman, P.A. Water–water and water–ion potential functions including terms for many body effects. J. Chem. Phys. 1985, 83, 2923–2933. [Google Scholar] [CrossRef]
- Yin, J.; Wu, M.M.; Xiao, H.; Ren, W.K.; Duan, J.L.; Yang, G.; Li, T.J.; Yin, Y.L. Development of an antioxidant system after early weaning in piglets. J. Anim. Sci. 2014, 92, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Klose, V.; Bayer, K.; Bruckbeck, R.; Schatzmayr, G.; Loibner, A.P. In vitro antagonistic activities of animal intestinal strains against swine-associated pathogens. Vet. Microbiol. 2010, 144, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Huang, X.; Fang, S.; He, M.; Zhao, Y.; Wu, Z.; Yang, M.; Zhang, Z.; Chen, C.; Huang, L. Unraveling the Fecal Microbiota and Metagenomic Functional Capacity Associated with Feed Efficiency in Pigs. Front. Microbiol. 2017, 8, 1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormack, U.M.; Curiao, T. Exploring a Possible Link between the Intestinal Microbiota and Feed Efficiency in Pigs. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Committee on Nutrient Requirements of Swine, National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Marquardt, R.R.; Jin, L.Z.; Kim, J.W.; Fang, L.; Frohlich, A.A.; Baidoo, S.K. Passive protective effect of egg-yolk antibodies against enterotoxigenic Escherichia coli K88+ infection in neonatal and early-weaned piglets. FEMS Immunol. Med. Microbiol. 1999, 23, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Hu, H.; Xiao, X.; Chen, D.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Luo, J.; Luo, Y.; et al. Lentinan administration relieves gut barrier dysfunction induced by rotavirus in a weaned piglet model. Food Funct. 2019, 10, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, H.; Pfirter, H.P.; Prabucki, A.L. A new method of determining metabolisability of energy and digestibility of fatty acids in broiler diets. Br. Poult. Sci. 1975, 16, 531–534. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of the Association of Official Analytical Chemists, 18th ed.; AOAC International: Gaitherburg, MD, USA, 2007; pp. 11–23. [Google Scholar]
- Bovera, F.; Lestingi, A.; Iannaccone, F.; Tateo, A.; Nizza, A. Use of dietary mannanoligosaccharides during rabbit fattening period: Effects on growth performance, feed nutrient digestibility, carcass traits, and meat quality. J. Anim. Sci. 2012, 90, 3858–3866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013. [Google Scholar] [CrossRef] [PubMed]
- Nienaber, J.A.; Hahn, G.L. Effects of water flow restriction and environmental factors on performance of nursery-age pigs. J. Anim. Sci. 1984, 59, 1423–1429. [Google Scholar] [CrossRef]
- Leibbrandt, V.D.; Johnston, L.J.; Shurson, G.C.; Crenshaw, J.D.; Libal, G.W.; Arthur, R.D. Effect of nipple drinker water flow rate and season on performance of lactating swine. J. Anim. Sci. 2001, 79, 2770–2775. [Google Scholar] [CrossRef] [PubMed]
- Browning, K.N.; Travagli, R.A. Central nervous system control of gastrointestinal motility and secretion and modulation of gastrointestinal functions. Compr. Physiol. 2014, 4, 1339–1368. [Google Scholar]
- Jimenez, C.; Regnard, J.; Robinet, C.; Mourot, L.; Gomez-Merino, D.; Chennaoui, M.; Jammes, Y.; Dumoulin, G.; Desruelle, A.V.; Melin, B. Whole body immersion and hydromineral homeostasis: Effect of water temperature. Eur. J. Appl. Physiol. 2010, 108, 49–58. [Google Scholar] [CrossRef]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet. Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Salyers, A.A. Bacteroides of the human lower intestinal tract. Annu. Rev. Microbiol. 1984, 38, 293–313. [Google Scholar] [CrossRef] [PubMed]
- Grover, M. Role of gut pathogens in development of irritable bowel syndrome. Indian. J. Med. Res. 2014, 139, 11–18. [Google Scholar] [PubMed]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD-what role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends. Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Durban, A.; Abellan, J.J.; Jimenez-Hernandez, N.; Artacho, A.; Garrigues, V.; Ortiz, V.; Ponce, J.; Latorre, A.; Moya, A. Instability of the faecal microbiota in diarrhoea-predominant irritable bowel syndrome. FEMS Microbiol. Ecol. 2013, 86, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothe, E. Special Focus: Actinobacteria. J. Basic. Microbiol. 2018, 58. [Google Scholar] [CrossRef] [PubMed]
- Pankratov, T.A.; Kirsanova, L.A.; Kaparullina, E.N.; Kevbrin, V.V.; Dedysh, S.N. Telmatobacter bradus gen. nov., sp. nov., a cellulolytic facultative anaerobe from subdivision 1 of the Acidobacteria, and emended description of Acidobacterium capsulatum Kishimoto et al. 1991. Int. J. Syst. Evol. Microbiol. 2012, 62, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichorst, S.A.; Kuske, C.R.; Schmidt, T.M. Influence of plant polymers on the distribution and cultivation of bacteria in the phylum Acidobacteria. Appl. Environ. Microbiol. 2011, 77, 586–596. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Sekiguchi, Y.; Imachi, H.; Kamagata, Y.; Ohashi, A.; Harada, H. Diversity, localization, and physiological properties of filamentous microbes belonging to Chloroflexi subphylum I in mesophilic and thermophilic methanogenic sludge granules. Appl. Environ. Microbiol. 2005, 71, 7493–7503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huws, S.A.; Lee, M.R.; Muetzel, S.M.; Scott, M.B.; Wallace, R.J.; Scollan, N.D. Forage type and fish oil cause shifts in rumen bacterial diversity. FEMS Microbiol. Ecol. 2010, 73, 396–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhao, Y.; Xu, J.; Xue, Z.; Zhang, M.; Pang, X.; Zhang, X.; Zhao, L. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwiertz, A.; Taras, D.; Schafer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity (Silver Spring) 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | DWT | 13 °C | 30 °C | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| DWF | 300 mL/min | 700 mL/min | 300 mL/min | 700 mL/min | DWT | DWF | DWT × DWF | |
| Initial BW (kg) | 8.60 ± 0.14 | 8.60 ± 0.24 | 8.60 ± 0.18 | 8.60 ± 0.08 | 1.000 | 1.000 | 1.000 | |
| Final BW (kg) | 20.54 ± 1.07 | 18.95 ± 1.57 | 20.79 ± 0.64 | 21.04 ± 1.12 | 0.065 | 0.267 | 0.139 | |
| ADG (kg/d) | 0.43 ± 0.04 | 0.37 ± 0.05 | 0.44 ± 0.02 | 0.44 ± 0.04 | 0.043 | 0.220 | 0.103 | |
| ADFI (kg/d) | 0.65 ± 0.06 | 0.63 ± 0.03 | 0.65 ± 0.04 | 0.66 ± 0.06 | 0.636 | 0.790 | 0.583 | |
| F: G | 1.53 ± 0.05 | 1.73 ± 0.25 | 1.49 ± 0.04 | 1.48 ± 0.02 | 0.045 | 0.168 | 0.124 | |
| Items | 13 °C | 30 °C | p-Value | ||||
|---|---|---|---|---|---|---|---|
| 300 mL/min | 700 mL/min | 300 mL/min | 700 mL/min | DWT | DWF | DWT × DWF | |
| Diarrhea rate (%) | 16.07 ± 2.31 | 17.63 ± 1.98 | 7.59 ± 4.28 | 9.37 ± 5.12 | 0.001 | 0.380 | 0.953 |
| Diarrhea index | 0.55 ± 0.04 | 0.63 ± 0.06 | 0.32 ± 0.06 | 0.41 ± 0.08 | 0.000 | 0.024 | 0.917 |
| Items | 13 °C | 30 °C | p-Value | ||||
|---|---|---|---|---|---|---|---|
| 300 mL/min | 700 mL/min | 300 mL/min | 700 mL/min | DWT | DWF | DWT × DWF | |
| CP (%) | 65.78 ± 2.38 | 67.21 ± 3.02 | 71.81 ± 2.21 | 72.96 ± 1.30 | 0.000 | 0.287 | 0.906 |
| Ash (%) | 48.05 ± 3.22 | 46.39 ± 1.67 | 48.85 ± 3.17 | 49.56 ± 1.81 | 0.266 | 0.785 | 0.499 |
| EE (%) | 56.60 ± 4.58 | 52.13 ± 3.22 | 59.80 ± 1.80 | 58.42 ± 5.44 | 0.035 | 0.169 | 0.456 |
| Energy (%) | 74.89 ± 2.30 | 73.54 ± 2.30 | 77.44 ± 0.98 | 76.99 ± 1.51 | 0.007 | 0.354 | 0.640 |
| Items | 13 °C | 30 °C | p-Value | ||||
|---|---|---|---|---|---|---|---|
| 300 mL/min | 700 mL/min | 300 mL/min | 700 mL/min | DWT | DWF | DWT × DWF | |
| Chao 1 | 885.12 ± 185.78 | 1212.21 ± 239.51 | 818.24 ± 946.79 | 946.79 ± 218.81 | 0.110 | 0.036 | 0.323 |
| Shannon | 4.13 ± 0.50 | 4.71 ± 0.22 | 4.19 ± 0.58 | 4.62 ± 0.32 | 0.960 | 0.035 | 0.715 |
| Simpson | 0.93 ± 0.02 | 0.97 ± 0.01 | 0.93 ± 0.06 | 0.96 ± 0.01 | 0.906 | 0.072 | 0.963 |
| PD | 48.76 ± 11.34 | 65.22 ± 13.85 | 43.47 ± 3.69 | 50.92 ± 8.32 | 0.075 | 0.035 | 0.387 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Li, Z.; Zhao, H.; Chen, X.; Tian, G.; Liu, G.; Cai, J.; Jia, G. Effects of Drinking Water Temperature and Flow Rate during Cold Season on Growth Performance, Nutrient Digestibility and Cecum Microflora of Weaned Piglets. Animals 2020, 10, 1048. https://doi.org/10.3390/ani10061048
Zhang Z, Li Z, Zhao H, Chen X, Tian G, Liu G, Cai J, Jia G. Effects of Drinking Water Temperature and Flow Rate during Cold Season on Growth Performance, Nutrient Digestibility and Cecum Microflora of Weaned Piglets. Animals. 2020; 10(6):1048. https://doi.org/10.3390/ani10061048
Chicago/Turabian StyleZhang, Zhenyu, Zeqiang Li, Hua Zhao, Xiaoling Chen, Gang Tian, Guangmang Liu, Jingyi Cai, and Gang Jia. 2020. "Effects of Drinking Water Temperature and Flow Rate during Cold Season on Growth Performance, Nutrient Digestibility and Cecum Microflora of Weaned Piglets" Animals 10, no. 6: 1048. https://doi.org/10.3390/ani10061048