Characteristics and Demography of a Free-Ranging Ethiopian Hedgehog, Paraechinus aethiopicus, Population in Qatar

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Animal Capture
2.2. Data Analysis
3. Results
3.1. Model Fit
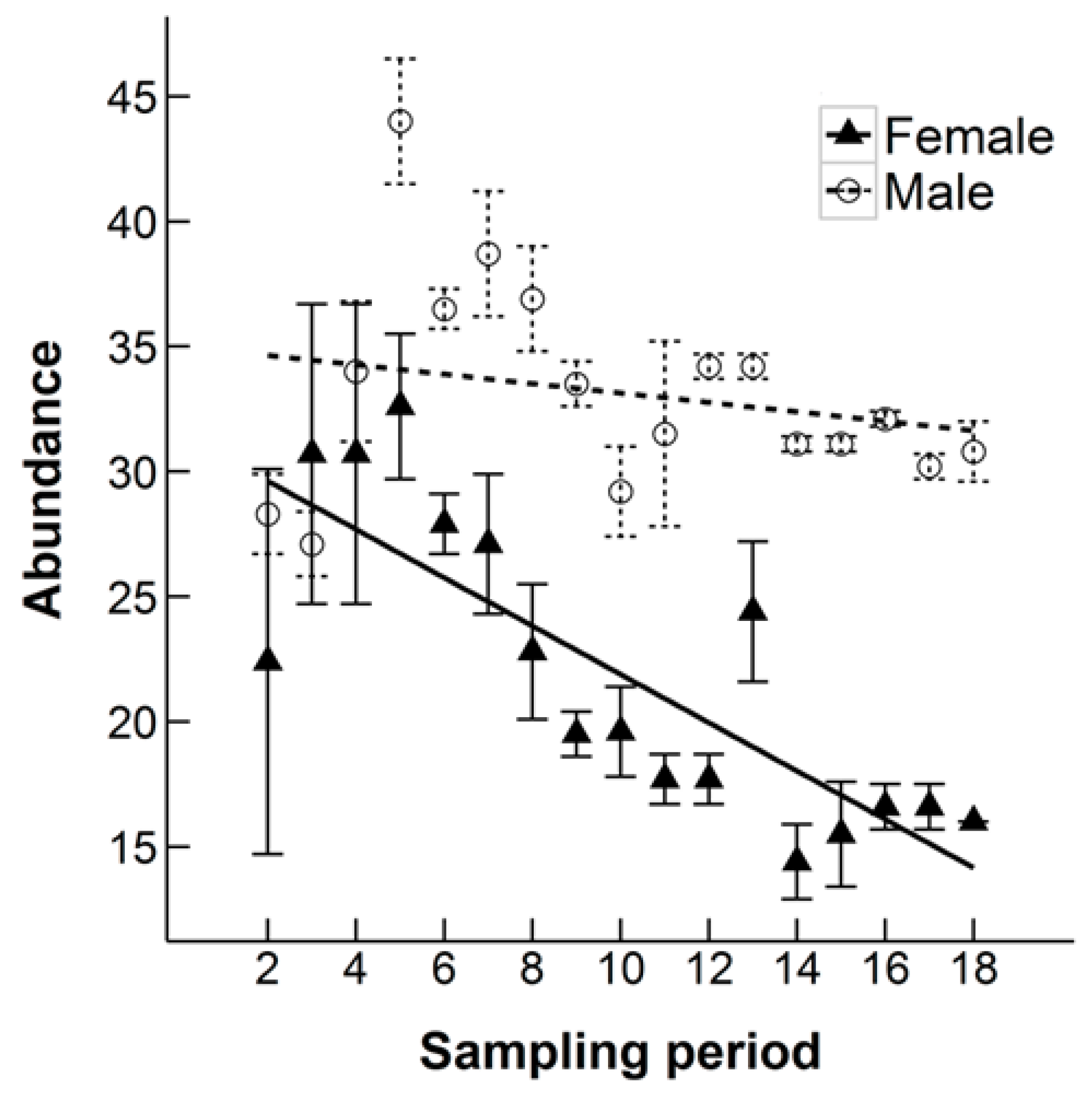
3.2. Population Size
3.3. Capture Probability
3.4. Survival
3.5. New Entrants
3.6. Evaluation of Methodology
4. Discussion
4.1. Comparison with Other Hedgehog Species
4.2. Evaluation of Methodology
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Reeve, N. Hedgehogs; Poyser: London, UK, 1994; ISBN 085661081X. [Google Scholar]
- He, K.; Chen, J.-H.; Gould, G.C.; Yamaguchi, N.; Ai, H.-S.; Wang, Y.-X.; Zhang, Y.-P.; Jiang, X.-L. An Estimation of Erinaceidae Phylogeny: A Combined Analysis Approach. PLoS ONE 2012, 7, e39304. [Google Scholar] [CrossRef] [PubMed]
- Al-Musfir, H.M.; Yamaguchi, N. Timings of hibernation and breeding of Ethiopian Hedgehogs, Paraechinus aethiopicus, in Qatar. Zool. Middle East 2008, 45, 3–10. [Google Scholar] [CrossRef]
- Zapletal, M.; Sodnompil, B.; Atwood, J.; Murdoch, J.D.; Reading, R.P. Home range characteristics and habitat selection by Daurian hedgehogs (Mesechinus dauuricus) in Ikh Nart Nature Reserve, Mongolia. Sciences 2012, 10, 41–50. [Google Scholar]
- Yamaguchi, N.; Al-Hajri, A.; Al-Jabiri, H. Timing of breeding in free-ranging Ethiopian hedgehogs, Paraechinus aethiopicus, from Qatar. J. Arid Environ. 2013, 99, 1–4. [Google Scholar] [CrossRef]
- Pettett, C.E.; Al-Hajri, A.; Al-Jabiry, H.; Macdonald, D.W.; Yamaguchi, N. A comparison of the Ranging behaviour and habitat use of the Ethiopian hedgehog (Paraechinus aethiopicus) in Qatar with hedgehog taxa from temperate environments. Sci.Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Murdoch, J.D.; Buyandelger, S.; Kenny, D.; Reading, R.P. Ecology of the Daurian hedgehog (Hemiechinus dauuricus) in Ikh Nart Nature Reserve, Mongolia: Preliminary findings. Mong. J. Biol. Sci. 2006, 4, 25–32. [Google Scholar]
- Boyles, J.G.; Bennett, N.C.; Mohammed, O.B.; Alagaili, A.N. Torpor patterns in Desert Hedgehogs (Paraechinus aethiopicus) represent another new point along a thermoregulatory continuum. Physiol. Biochem. Zool. 2017, 90, 445–452. [Google Scholar] [CrossRef]
- Alagaili, A.N.; Bennett, N.C.; Mohammed, O.B.; Hart, D.W. The reproductive biology of the Ethiopian hedgehog, Paraechinus aethiopicus, from central Saudi Arabia: The role of rainfall and temperature. J. Arid Environ. 2017, 145, 1–9. [Google Scholar] [CrossRef]
- Alagaili, A.N.; Bennett, N.C.; Amor, N.M.; Hart, D.W. The locomotory activity patterns of the arid-dwelling desert hedgehog, Paraechinus aethiopicus, from Saudi Arabia. J. Arid Environ. 2020, 177, 104141. [Google Scholar] [CrossRef]
- Abu Baker, M.A.; Reeve, N.; Mohedano, I.; Conkey, A.A.T.; Macdonald, D.W.; Yamaguchi, N. Caught basking in the winter sun: Preliminary data on winter thermoregulation in the Ethiopian hedgehog, Paraechinus aethiopicus, in Qatar. J. Arid Environ. 2016, 125, 52–55. [Google Scholar] [CrossRef]
- Zapletal, M.; Sodnompil, B.; Atwood, J.L.; Murdoch, J.D.; Reading, R.P. Fine-scale habitat use by Daurian hedgehogs in the Gobi-steppe of Mongolia. J. Arid Environ. 2015, 114, 100–103. [Google Scholar] [CrossRef]
- Abu Baker, M.A.; Reeve, N.; Conkey, A.A.T.; Macdonald, D.W.; Yamaguchi, N. Hedgehogs on the move: Testing the effects of land use change on home range size and movement patterns of free-ranging Ethiopian hedgehogs. PLoS ONE 2017, 12, e0180826. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.L.; Bates, P.J.J. The mammals of Arabia; Harrison Zoological Museum: Sevenoaks, UK, 1991; Volume 354. [Google Scholar]
- Micol, T.; Doncaster, C.P.; Mackinlay, L.A. Correlates of local variation in the abundance of hedgehogs Erinaceus europaeus. J. Anim. Ecol. 1994, 63, 851–860. [Google Scholar] [CrossRef]
- Haigh, A.; Butler, F.; Riordan, R.M.O. An investigation into the techniques for detecting hedgehogs in a rural landscape. J. Negat. Results Ecol. Evol. Biol. 2012, 9, 15–26. [Google Scholar]
- Doncaster, C.P. Factors regulating local variations in abundance: Field tests on hedgehogs, Erinaceus europaeus. Oikos 1994, 69, 182–192. [Google Scholar] [CrossRef]
- Jackson, D.B. Factors affecting the abundance of introduced hedgehogs (Erinaceus europaeus) to the Hebridean island of South Uist in the absence of natural predators and implications for nesting birds. J. Zool. 2007, 271, 210–217. [Google Scholar] [CrossRef]
- Yarnell, R.W. Using occupancy analysis to validate the use of footprint tunnels as a method for monitoring the hedgehog Erinaceus europaeus. Mamm. Rev. 2014, 44, 234–238. [Google Scholar] [CrossRef]
- Huijser, M.P.; Bergers, P.J.M. The effect of roads and traffic on hedgehog (Erinaceus europaeus) populations. Biol. Conserv. 2000, 95, 6–9. [Google Scholar] [CrossRef]
- Williams, B.; Mann, N.; Neumann, J.L.; Yarnell, R.W.; Baker, P.J. A prickly problem: Developing a volunteer-friendly tool for monitoring populations of a terrestrial urban mammal, the West European hedgehog (Erinaceus europaeus). Urban Ecosyst. 2018, 21, 1075–1086. [Google Scholar] [CrossRef]
- Williams, B.M.; Baker, P.J.; Thomas, E.; Wilson, G.; Judge, J.; Yarnell, R.W. Reduced occupancy of hedgehogs (Erinaceus europaeus) in rural England and Wales: The influence of habitat and an asymmetric intra-guild predator. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. The value of green-spaces in built-up areas for western hedgehogs. Lutra 2009, 52, 69–82. [Google Scholar]
- Van De Poel, J.L.; Dekker, J.; Van Langevelde, F. Dutch hedgehogs Erinaceus europaeus are nowadays mainly found in urban areas, possibly due to the negative effects of badgers Meles meles. Wildl. Biol. 2015, 21, 51–55. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. Quantifying the long-term decline of the West European hedgehog in England by subsampling citizen-science datasets. Eur. J. Wildl. Res. 2016, 62, 407–413. [Google Scholar] [CrossRef]
- Aebischer, N.J.; Davey, P.D.; Kingdon, N.G. National Gamebag Census: Mammal Trends to 2009. Available online: http://www.gwct.org.uk/ngcmammals (accessed on 23 April 2020).
- Pettett, C.E.; Johnson, P.J.; Moorhouse, T.P.; Macdonald, D.W. National predictors of hedgehog Erinaceus europaeus distribution and decline in Britain. Mamm. Rev. 2018, 48, 1–6. [Google Scholar] [CrossRef]
- Reeve, N.J. A Field Study of the Hedgehog (Erinaceus europaeus) with Particular Referance to Movements and Behaviour. Ph.D. Thesis, University of London, London, UK, 1981. [Google Scholar]
- Kristiansson, H. Population variables and causes of mortality in a hedgehog (Erinaceous europaeus) population in southern Sweden. J. Zool. 1990, 220, 391–404. [Google Scholar] [CrossRef]
- Jackson, D.B.; Green, R.E. The importance of the introduced hedgehog (Erinaceus europaeus) as a predator of the eggs of waders (Charadrii) on machair in South Uist, Scotland. Biol. Conserv. 2000, 93, 333–348. [Google Scholar] [CrossRef]
- White, G.C. Capture-Recapture and Removal Methods for Sampling Closed Populations; Los Alamos National Laboratory: Los Alamos, NM, USA, 1982. [Google Scholar]
- Otis, D.L.; Burnham, K.P.; White, G.C.; Anderson, D.R. Statistical inference from capture data on closed animal populations. Wildl. Monogr. 1978, 62, 3–135. [Google Scholar]
- Cormack, R.M. Examples of the use of GLIM to analyse capture-recapture studies. In Statistics in Ornithology; Springer: Berlin/Heidelberg, Germany, 1985; pp. 243–273. [Google Scholar]
- Cormack, R.M. Log-linear models for capture-recapture. Biometrics 1989, 395–413. [Google Scholar] [CrossRef]
- Rivest, L.; Daigle, G. Loglinear models for the robust design in mark–recapture experiments. Biometrics 2004, 60, 100–107. [Google Scholar] [CrossRef]
- Seber, G.A.F. The Estimation of Animal Abundance and Related Parameters; Blackburn Press: Caldwell, NJ, USA, 1982; Volume 8. [Google Scholar]
- Baillargeon, S.; Rivest, L.-P. Rcapture: Loglinear models for capture-recapture in R. J. Stat. Softw. 2007, 19, 1–31. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. The value of agri-environment schemes for macro-invertebrate feeders: Hedgehogs on arable farms in Britain. Anim. Conserv. 2010, 13, 467–473. [Google Scholar] [CrossRef]
- Wembridge, D.E.; Newman, M.R.; Bright, P.W.; Morris, P.A. An estimate of the annual number of hedgehog (Erinaceus europaeus) road casualties in Great Britain. Mammal Commun. 2016, 2, 8–14. [Google Scholar]
- Walhovd, H. Records of young hedgehogs (Erinaceus europaeus L.) in a private garden. Z. Säugetierkd. 1990, 55, 289–297. [Google Scholar]
- Baker, M.A.A.; Villamil, L.A.; Reeve, N.; Karanassos, C.; Mahtab, H.; Yamaguchi, N. Into the wild: Survival, movement patterns, and weight changes in captive Ethiopian hedgehogs, Paraechinus aethiopicus following their release. J. Arid Environ. 2018, 158, 43–46. [Google Scholar] [CrossRef]
- Pettett, C.E.; Moorhouse, T.P.; Johnson, P.J.; Macdonald, D.W. Factors affecting hedgehog (Erinaceus europaeus) attraction to rural villages in arable landscapes. Eur. J. Wildl. Res. 2017, 63, 54. [Google Scholar] [CrossRef]
- Kristiansson, H. Ecology of a Hedgehog (Erinaceus europaeus) Population in Southern Sweden. Ph.D. Thesis, Department of Animal Ecology, University of Lund, Lund, Sweden, 1984. [Google Scholar]
- Kristiansson, H. Young Production of European Hedgehog in Sweden and Britain. Acta Theriol. 1981, 26, 504–507. [Google Scholar] [CrossRef]
- Hubert, P.; Julliard, R.; Biagianti, S.; Poulle, M.L. Ecological factors driving the higher hedgehog (Erinaceus europeaus) density in an urban area compared to the adjacent rural area. Landsc. Urban Plan. 2011, 103, 34–43. [Google Scholar] [CrossRef]
- Young, R.P.; Davison, J.; Trewby, I.D.; Wilson, G.J.; Delahay, R.J.; Doncaster, C.P. Abundance of hedgehogs (Erinaceus europaeus) in relation to the density and distribution of badgers (Meles meles). J. Zool. 2006, 269, 349–356. [Google Scholar] [CrossRef]
- Warwick, H.; Morris, P.; Walker, D. Survival and weight changes of hedgehogs (Erinaceus europaeus) translocated from the Hebrides to Mainland Scotland. Lutra 2006, 49, 89–102. [Google Scholar]
- Morris, P.A.; Warwick, H. A study of rehabilitated juvenile hedgehogs after release into the wild. Anim. Welf. 1994, 3, 163–177. [Google Scholar]
- Molony, S.E.; Dowding, C.V.; Baker, P.J.; Cuthill, I.C.; Harris, S. The effect of translocation and temporary captivity on wildlife rehabilitation success: An experimental study using European hedgehogs (Erinaceus europaeus). Biol. Conserv. 2006, 130, 530–537. [Google Scholar] [CrossRef]
- Morris, P.A. Released, rehabilitated hedgehogs: A follow-up study in Jersey. Anim. Welf. 1997, 6, 317–327. [Google Scholar]
- Rasmussen, S.L.; Berg, T.B.; Dabelsteen, T.; Jones, O.R. The ecology of suburban juvenile European hedgehogs (Erinaceus europaeus) in Denmark. Ecol. Evol. 2019, 9, 13174–13187. [Google Scholar] [CrossRef] [PubMed]
- Pettett, C.E. Factors Affecting Hedgehog Distribution and Habitat Selection in Rural Landscapes; University of Oxford: Oxford, UK, 2016. [Google Scholar]
- Amori, G.; De Silvestro, V.; Ciucci, P.; Luiselli, L. Quantifying whether different demographic models produce incongruent results on population dynamics of two long-term studied rodent species. Eur. J. Ecol. 2017, 3, 18–26. [Google Scholar] [CrossRef]
- Casula, P.; Luiselli, L.; Amori, G. Which population density affects home ranges of co-occurring rodents? Basic Appl. Ecol. 2019, 34, 46–54. [Google Scholar] [CrossRef]
- Reeve, N.; Bowen, C.; Gurnell, J. An Improved Identification Marking Method for Hedgehogs; The Mammal Socity: Southampton, UK, 2019. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subset | Block | No. in Model 1 | Adjustments to Improve Model Fit | AIC | Ntot 2 | SE | N 3 | SE | Density (km−2) 4 |
|---|---|---|---|---|---|---|---|---|---|
| All | All 19 months | 144 | None | 1180 | 149 | 2.7 | 55 | 2.4 | 6.5 |
| All | All 19 months | 144 | Excluding those captured all 19 times and including residuals <10 | 439 | 151 | 0 | 60 | 2.9 | 7.0 |
| Males | All 19 months | 75 | None | 787 | 77 | 1.6 | 32 | 1.1 | 3.8 |
| Males | All 19 months | 75 | Excluding those captured all 19 times and including residuals <50 | 489 | 77 | 0 | 33 | 1.0 | 3.9 |
| Females | All 19 months | 62 | None | 540 | 65 | 2.3 | 22 | 1.5 | 2.6 |
| Females | All 19 months | 62 | Capture constant on model and including residuals <800 5 | 511 | 65 | 1.9 | 21 | 1.5 | 2.5 |
| All | Excluding winter | 112 | None | 346 | 115 | 2.3 | 52 | 3.8 | 6.1 |
| All | Excluding winter | 112 | Excluding those captured all 9 times | 327 | 117 | 1.9 | 54 | 3.7 | 6.3 |
| Males | Excluding winter | 59 | None | 244 | 60 | 1.2 | 30 | 1.9 | 3.5 |
| Males | Excluding winter | 59 | Excluding those captured 8 or 9 times and including residuals <6 | 177 | 61 | 0 | 30 | 1.7 | 3.5 |
| Females | Excluding winter | 47 | None | 180 | 47 | 1.1 | 19 | 1.9 | 2.2 |
| Females | Excluding winter | 47 | Excluding those captured 8 or 9 times | 142 | 47 | 0 | 19 | 1.9 | 2.2 |
| Adults | Excluding winter | 92 | None | 304 | 93 | 1.5 | 45 | 3.9 | 5.3 |
| Adults | Excluding winter | 92 | Excluding those caught all 9 times and including residuals <10 | 266 | 94 | 0 | 46 | 3.9 | 5.4 |
| Juveniles | Excluding winter | 21 | None | 100 | 26 | 5.1 | 7 | 1.8 | 0.8 6 |
| Subset | Block | No. Hedgehogs 1 | Ntot | % of Population Captured |
|---|---|---|---|---|
| All | All 19 months | 144 | 151 | 95.36 |
| Males | All 19 months | 75 | 77 | 97.40 |
| Females | All 19 months | 62 | 65 | 95.38 |
| All | Excluding winter | 112 | 117 | 95.73 |
| Males | Excluding winter | 59 | 61 | 96.72 |
| Females | Excluding winter | 47 | 47 | 100.00 |
| Adults | Excluding winter | 92 | 94 | 97.87 |
| Juveniles | Excluding winter | 21 | 23 | 91.30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pettett, C.; W. Macdonald, D.; Al-Hajiri, A.; Al-Jabiry, H.; Yamaguchi, N. Characteristics and Demography of a Free-Ranging Ethiopian Hedgehog, Paraechinus aethiopicus, Population in Qatar. Animals 2020, 10, 951. https://doi.org/10.3390/ani10060951
Pettett C, W. Macdonald D, Al-Hajiri A, Al-Jabiry H, Yamaguchi N. Characteristics and Demography of a Free-Ranging Ethiopian Hedgehog, Paraechinus aethiopicus, Population in Qatar. Animals. 2020; 10(6):951. https://doi.org/10.3390/ani10060951
Chicago/Turabian StylePettett, Carly, David W. Macdonald, Afra Al-Hajiri, Hayat Al-Jabiry, and Nobuyuki Yamaguchi. 2020. "Characteristics and Demography of a Free-Ranging Ethiopian Hedgehog, Paraechinus aethiopicus, Population in Qatar" Animals 10, no. 6: 951. https://doi.org/10.3390/ani10060951
APA StylePettett, C., W. Macdonald, D., Al-Hajiri, A., Al-Jabiry, H., & Yamaguchi, N. (2020). Characteristics and Demography of a Free-Ranging Ethiopian Hedgehog, Paraechinus aethiopicus, Population in Qatar. Animals, 10(6), 951. https://doi.org/10.3390/ani10060951