Immune Responses and Performance Are Influenced by Respiratory Vaccine Antigen Type and Stress in Beef Calves

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Feed Delivery and Health Observation
2.3. Treatments and Vaccination
2.4. Blood Collection and Serology
2.5. Statistical Analyses
3. Results and Discussion
3.1. Health and Performance
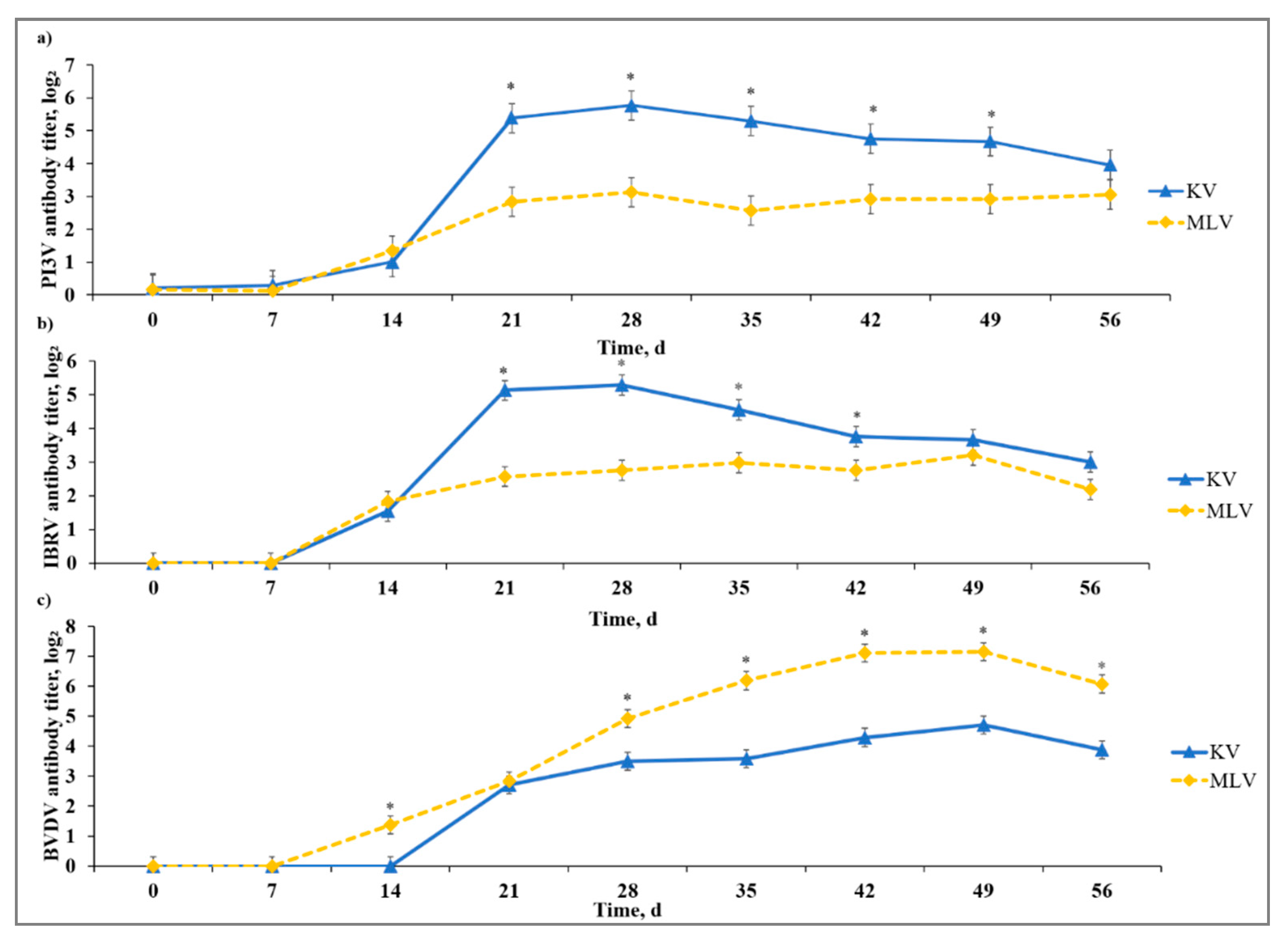
3.2. Virus-Specific Antibody Response
3.3. Serum Haptoglobin
3.4. Serum Cortisol
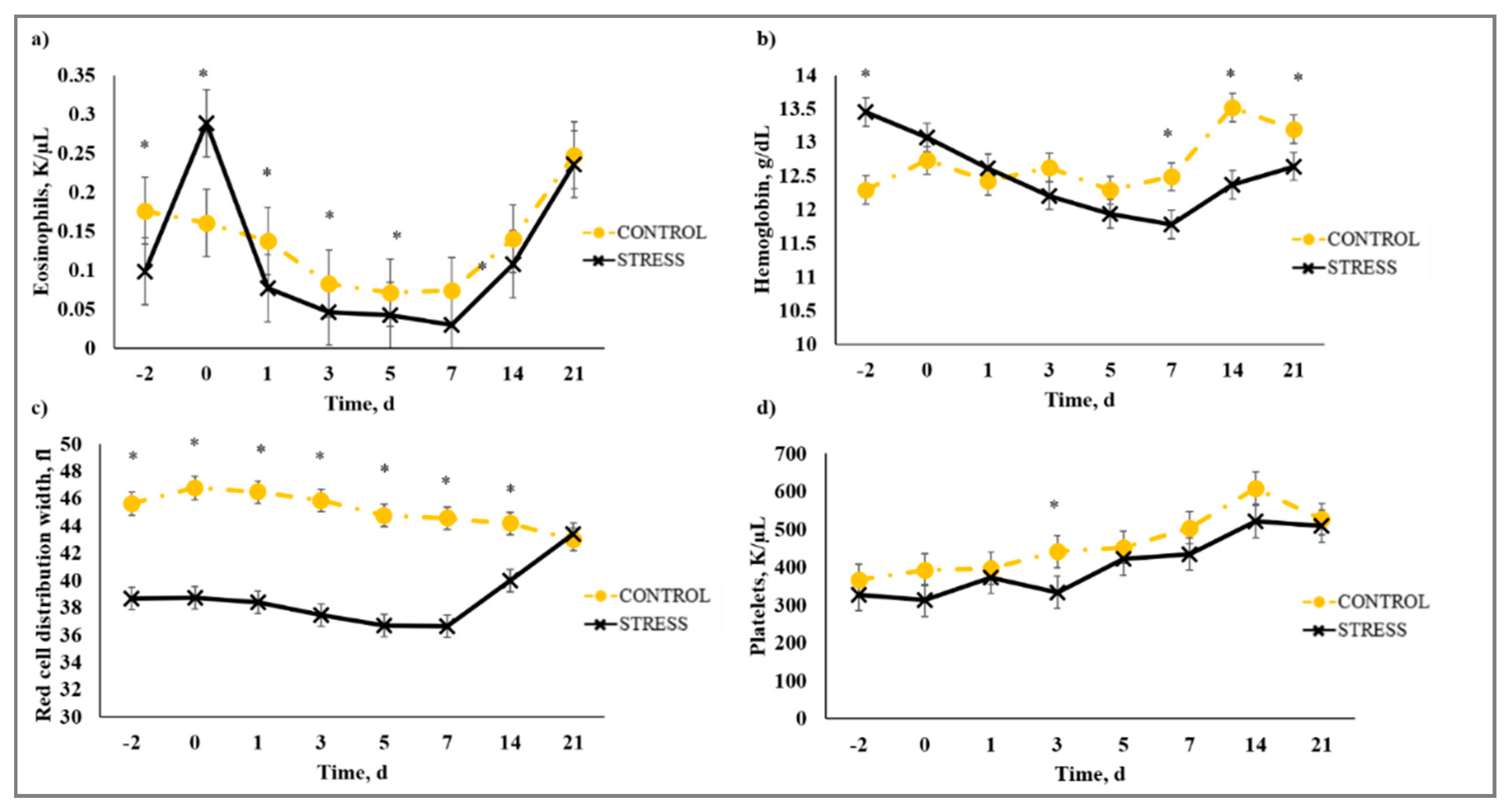
3.5. Complete Blood Count
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Comerford, J. Causes of Vaccine Failure in Beef Cattle. PennState Extension. 2017. Available online: https://extension.psu.edu/causes-of-vaccine-failure-in-beef-cattle (accessed on 26 June 2020).
- Dragoş, D.; Tănăsescu, M.D. The effect of stress on the defense systems. J. Med. Life 2010, 3, 10. [Google Scholar]
- Hughes, H.D.; Carroll, J.A.; Burdick Sanchez, N.C.; Richeson, J.T. Natural variations in the stress and acute phase responses of cattle. Innate Immun. 2013, 20, 888–896. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.A.; Burdick Sanchez, N.C. Relationship between Stress and Health in Cattle—Part 2. Technical Bulletin; Prince Agri Products: Quincy, IL, USA, 2013. [Google Scholar]
- Roth, J.A. Cortisol as mediator of stress-associated immunosuppression in cattle. In Animal Stress; Moberg, G.P., Ed.; Springer: New York, NY, USA, 1985; pp. 225–243. [Google Scholar]
- Burton, J.L.; Kehrli, M.E.; Kapil, S.; Horst, R.L. Regulation of L-selectin and CD18 on bovine neutrophils by glucocorticoids: Effects of cortisol and dexamethasone. J. Leukoc. Biol. 1995, 57, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Lynch, E.M.; Earley, B.; McGee, M.; Doyle, S. Effect of abrupt weaning at housing on leukocyte distribution, functional activity of neutrophils, and acute phase protein response of beef calves. BMC Vet. Res. 2010, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Roth, J.A.; Kaeberle, M.L.; Hsu, W.H. Effects of ACTH administration on bovine polymorphonuclear leukocyte function and lymphocyte blastogenesis. Am. J. Vet. Res. 1982, 43, 412–416. [Google Scholar]
- Chrousos, G.P. The hypothalamic–pituitary–adrenal axis and immune-mediated inflammation. N. Engl. J. Med. 1995, 332, 1351–1363. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richeson, J.T.; Carroll, J.A.; Burdick Sanchez, N.C.; May, N.D.; Hughes, H.D.; Roberts, S.L.; Broadway, P.R.; Sharon, K.P.; Ballou, M.A. Dexamethasone treatment differentially alters viral shedding and the antibody and acute phase protein response after multivalent respiratory vaccination in beef steers. J. Anim. Sci. 2016, 94, 3501–3509. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.B.; Cooke, R.F.; Hansen, G.R.; Staples, C.R.; Arthington, J.D. Effects of rumen-protected polyunsaturated fatty acid supplementation on performance and physiological responses of growing cattle after transportation and feedlot entry. J. Anim. Sci. 2010, 88, 4120–4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, J.A.; Forsberg, N.E. Influence of stress and nutrition on cattle immunity. Vet. Clin. North Am. Food Anim. Pract. 2007, 23, 105–149. [Google Scholar] [CrossRef] [PubMed]
- Cooke, R.F. Nutritional and management considerations for beef cattle experiencing stress-induced inflammation. Prof. Anim. Sci. 2017, 33, 1–11. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle: Eight Revised Edition; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar] [CrossRef]
- Rosenbaum, M.J.; Edwards, E.A.; Sullivan, E.J. Micromethods for respiratory virus sero-epidemiology. Health Lab. Sci. 1970, 7, 42–52. [Google Scholar] [PubMed]
- Robinson, P.H.; Wiseman, J.; Uden, P.; Mateos, G. Some experimental design and statistical criteria for analysis of studies in manuscripts submitted for consideration for publication. Anim. Feed Sci. Technol. 2016, 129, 1–11. [Google Scholar] [CrossRef]
- Grooms, D.L.; Coe, P. Neutralizing antibody responses in preconditioned calves following vaccination for respiratory viruses. Vet. Ther. 2002, 3, 119–127. [Google Scholar] [PubMed]
- Fulton, R.W.; Confer, A.W.; Burge, L.J.; Perino, L.J.; D’Offay, J.M.; Payton, M.E.; Mock, R.E. Antibody responses by cattle after vaccination with commercial viral vaccines containing bovine herpesvirus-1, bovine viral diarrhea virus, parainfluenza-3 virus, and bovine respiratory syncytial virus immunogens and subsequent revaccination at day 140. Vaccine 1995, 13, 725–733. [Google Scholar] [CrossRef]
- Roth, J.A.; Kaeberle, M.L. Suppression of neutrophil and lymphocyte function induced by a vaccinal strain of bovine viral diarrhea virus with and without the administration of ACTH. Am. J. Vet. Res. 1983, 44, 2366–2372. [Google Scholar] [PubMed]
- Baumann, H.; Gauldie, J. The acute phase response. Immunol. Today 1994, 15, 74–80. [Google Scholar] [CrossRef]
- Godson, D.L.; Campos, M.; Attah-Poku, S.K.; Redmond, M.J.; Cordeiro, D.M.; Sethi, M.S.; Harland, R.J.; Babiuk, L.A. Serum haptoglobin as an indicator of the acute phase response in bovine respiratory disease. Vet. Immunol. Immunopathol. 1996, 51, 277–292. [Google Scholar] [CrossRef]
- Deak, T.; Meriwether, J.L.; Fleshner, M.; Spencer, R.L.; Abouhamze, A.; Moldawer, L.L.; Grahn, R.E.; Watkins, L.R.; Maier, S.F. Evidence that brief stress may induce the acute phase response in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1997, 273, R1998–R2004. [Google Scholar] [CrossRef]
- Nukina, H.; Sudo, N.; Aiba, Y.; Oyama, N.; Koga, Y.; Kubo, C. Restraint stress elevates the plasma interleukin-6 levels in germ-free mice. J. Neuroimmunol. 2001, 115, 46–52. [Google Scholar] [CrossRef]
- Murata, H.; Shimada, N.; Yoshioka, M. Current research on acute phase proteins in veterinary diagnosis: An overview. Vet. J. 2004, 168, 28–40. [Google Scholar] [CrossRef]
- Arthington, J.D.; Cooke, R.F.; Maddock, T.D.; Araujo, D.B.; Moriel, P.; DiLorenzo, N.; Lamb, G.C. Effects of vaccination on the acute-phase protein response and measures of performance in growing beef calves. J. Anim. Sci. 2013, 91, 1831–1837. [Google Scholar] [CrossRef]
- Grandin, T. Assessment of stress during handling and transport. J. Anim. Sci. 1997, 75, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queyras, A.; Carosi, M. Non-invasive techniques for analyzing hormonal indicators of stress. Ann. Ist. Super. Sanita 2003, 40, 211–221. [Google Scholar]
- Baum, A.; Grunberg, N. Measurement of stress hormones. In Measuring Stress: A Guide for Health and Social Scientists; Cohen, S., Kessler, R.C., Gordon, L.U., Eds.; Oxford University Press: New York, NY, USA, 1997; pp. 175–192. [Google Scholar]
- Burton, J.L.; Madsen, S.A.; Chang, L.C.; Weber, P.S.; Buckham, K.R.; Van Dorp, R.; Hickey, M.C.; Earley, B. Gene expression signatures in neutrophils exposed to glucocorticoids: A new paradigm to help explain “neutrophil dysfunction” in parturient dairy cows. Vet. Immunol. Immunopathol. 2005, 105, 197–219. [Google Scholar] [CrossRef]
- Aguilera, G. Corticotropin releasing hormone, receptor regulation and the stress response. Trends Endocrinol. Metab. 1998, 329–336. [Google Scholar] [CrossRef]
- Hughes, H.D.; Carroll, J.A.; Burdick Sanchez, N.C.; Roberts, S.L.; Broadway, P.R.; May, N.D.; Ballou, M.A.; Richeson, J.T. Effects of dexamethasone treatment and respiratory vaccination on rectal temperature, complete blood count, and functional capacities of neutrophils in beef steers. J. Anim. Sci. 2017, 95, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; McEwen, B.S. Bidirectional effects of stress and glucocorticoid hormones on immune function: Possible explanations for paradoxical observations. Psychoneuroimmunology 2001, 1, 301–338. [Google Scholar]
- Heidt, T.; Sager, H.B.; Courties, G.; Dutta, P.; Iwamoto, Y.; Zaltsman, A.; Muhlen, C.; Bode, C.; Fricchione, G.L.; Denninger, J.; et al. Chronic variable stress activates hematopoietic stem cells. Nat. Med. 2014, 20, 754–758. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, E.L.; Davidson, J.M.; Beck, P.A.; Roberts, S.L.; Hughes, H.D.; Richeson, J.T. Effect of water restriction on performance, hematology, and antibody responses in parenteral or intranasal modified-live viral vaccinated beef calves. Bov. Pract. 2017, 51, 174–183. [Google Scholar]
- Anderson, B.H.; Watson, D.L.; Colditz, I.G. The effect of dexamethasone on some immunological parameters in cattle. Vet. Res. Commun. 1999, 23, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Saini, N.S.; Kumar, A.; Mahajan, S.K.; Sood, A.C. The use of ultrasonography, radiography, and surgery in the successful recovery from diaphragmatic hernia in a cow. Can. Vet. J. 2007, 48, 757. [Google Scholar] [PubMed]
- Schaefer, A.L.; Jones SD, M.; Tong AK, W.; Lepage, P.; Murray, N.L. The effects of withholding feed and water on selective blood metabolites in market-weight beef steers. Can. J. Anim. Sci. 1990, 70, 1155–1158. [Google Scholar] [CrossRef]
- Ellis, J.A.; Davis, W.C.; Belden, E.L.; Pratt, D.L. Flow cytofluorimetric analysis of lymphocyte subset alterations in cattle infected with bovine viral diarrhea virus. Vet. Pathol. 1988, 25, 231–236. [Google Scholar] [CrossRef]
- Gleich, G.J. Mechanisms of eosinophil-associated inflammation. J. Allergy Clin. Immunol. 2000, 105, 651–663. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment 1 | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CKV | CMLV | SKV | SMLV | Stress Model | Vaccine Type | Interaction | ||
| DMI, kg/day | ||||||||
| Days 0–7 | 6.20 | 6.18 | 4.70 | 4.57 | 0.15 | 0.01 | 0.61 | 0.70 |
| Days 7–14 | 6.16 | 5.00 | 5.24 | 4.89 | 0.54 | 0.37 | 0.20 | 0.47 |
| Days 14–21 | 6.99 | 6.22 | 6.00 | 5.98 | 0.47 | 0.23 | 0.43 | 0.45 |
| Days 21–28 | 7.80 | 7.13 | 7.02 | 7.06 | 0.41 | 0.33 | 0.47 | 0.42 |
| Days 28–35 | 8.00 | 7.86 | 7.68 | 7.83 | 0.41 | 0.68 | 0.98 | 0.73 |
| Days 35–24 | 8.46 | 7.76 | 7.98 | 7.49 | 0.54 | 0.50 | 0.30 | 0.85 |
| Days 42–49 | 8.66 | 7.76 | 8.16 | 7.76 | 0.59 | 0.68 | 0.30 | 0.68 |
| Days 49–56 | 9.38 | 8.42 | 8.54 | 7.97 | 0.46 | 0.20 | 0.13 | 0.68 |
| Days 0–14 | 6.18 | 5.59 | 4.97 | 4.73 | 0.33 | 0.01 | 0.24 | 0.61 |
| Days 0–28 | 6.79 | 6.13 | 5.74 | 5.62 | 0.38 | 0.08 | 0.34 | 0.50 |
| Days 0–56 | 7.71 | 7.04 | 6.92 | 6.69 | 0.40 | 0.20 | 0.30 | 0.60 |
| ADG, kg/day | ||||||||
| Days 0–7 | 0.31 | 0.27 | 0.31 | 0.78 | 0.36 | 0.46 | 0.55 | 0.47 |
| Days 7–14 | 0.95 | 0.02 | 0.98 | 0.22 | 2.53 | 0.96 | 0.74 | 0.97 |
| Days 14–21 | 1.75 | 2.09 | 1.08 | 1.24 | 2.63 | 0.76 | 0.92 | 0.97 |
| Days 21–28 | 2.05 ᵃ | 1.61 ᵇ | 1.85 ᵃᵇ | 1.97 ᵃᵇ | 0.13 | 0.54 | 0.21 | 0.04 |
| Days 28–35 | 1.40 | 1.81 | 1.28 | 1.86 | 0.24 | 0.88 | 0.04 | 0.72 |
| Days 35–24 | 1.53 | 0.89 | 1.34 | 0.55 | 0.22 | 0.23 | 0.01 | 0.24 |
| Days 42–49 | 1.97 | 1.27 | 1.83 | 1.60 | 0.20 | 0.62 | 0.02 | 0.21 |
| Days 49–56 | 1.94 | 1.94 | 2.02 | 1.75 | 0.25 | 0.81 | 0.57 | 0.56 |
| Days 0–14 | 0.63 | −0.15 | 0.65 | 0.53 | 1.29 | 0.78 | 0.72 | 0.79 |
| Days 0–28 | 1.26 | 0.85 | 1.06 | 1.08 | 0.14 | 0.92 | 0.15 | 0.10 |
| Days 0–56 | 1.48 | 1.16 | 1.34 | 1.28 | 0.10 | 0.86 | 0.05 | 0.18 |
| Item | Treatment 1 | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CKV | CMLV | SKV | SMLV | Stress Model | Vaccine Type | Interaction | ||
| BW, kg | ||||||||
| Day 0 | 217 | 239 | 224 | 220 | 11.30 | 0.60 | 0.41 | 0.26 |
| Day 7 | 219 | 239 | 226 | 226 | 11.52 | 0.79 | 0.38 | 0.37 |
| Day 14 | 226 | 237 | 233 | 226 | 11.75 | 0.86 | 0.85 | 0.42 |
| Day 21 | 238 | 252 | 241 | 236 | 11.76 | 0.57 | 0.69 | 0.42 |
| Day 28 | 252 | 263 | 254 | 250 | 12.05 | 0.60 | 0.77 | 0.53 |
| Day 35 | 262 | 276 | 263 | 263 | 11.97 | 0.59 | 0.55 | 0.57 |
| Day 42 | 272 | 282 | 272 | 267 | 12.50 | 0.52 | 0.86 | 0.54 |
| Day 49 | 286 | 291 | 285 | 273 | 13.24 | 0.44 | 0.78 | 0.52 |
| Day 56 | 300 | 305 | 299 | 291 | 12.98 | 0.56 | 0.90 | 0.62 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hudson, R.E.; Tomczak, D.J.; Kaufman, E.L.; Adams, A.M.; Carroll, J.A.; Broadway, P.R.; Ballou, M.A.; Richeson, J.T. Immune Responses and Performance Are Influenced by Respiratory Vaccine Antigen Type and Stress in Beef Calves. Animals 2020, 10, 1119. https://doi.org/10.3390/ani10071119
Hudson RE, Tomczak DJ, Kaufman EL, Adams AM, Carroll JA, Broadway PR, Ballou MA, Richeson JT. Immune Responses and Performance Are Influenced by Respiratory Vaccine Antigen Type and Stress in Beef Calves. Animals. 2020; 10(7):1119. https://doi.org/10.3390/ani10071119
Chicago/Turabian StyleHudson, Rachel E., Dexter J. Tomczak, Emily L. Kaufman, Ashlee M. Adams, Jeffery A. Carroll, Paul R. Broadway, Michael A. Ballou, and John T. Richeson. 2020. "Immune Responses and Performance Are Influenced by Respiratory Vaccine Antigen Type and Stress in Beef Calves" Animals 10, no. 7: 1119. https://doi.org/10.3390/ani10071119