Effect of Social Rank upon Estrus Induction and Some Reproductive Outcomes in Anestrus Goats Treated With Progesterone + eCG

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. General
2.2. Location, Environmental Conditions, Animals, and their Management
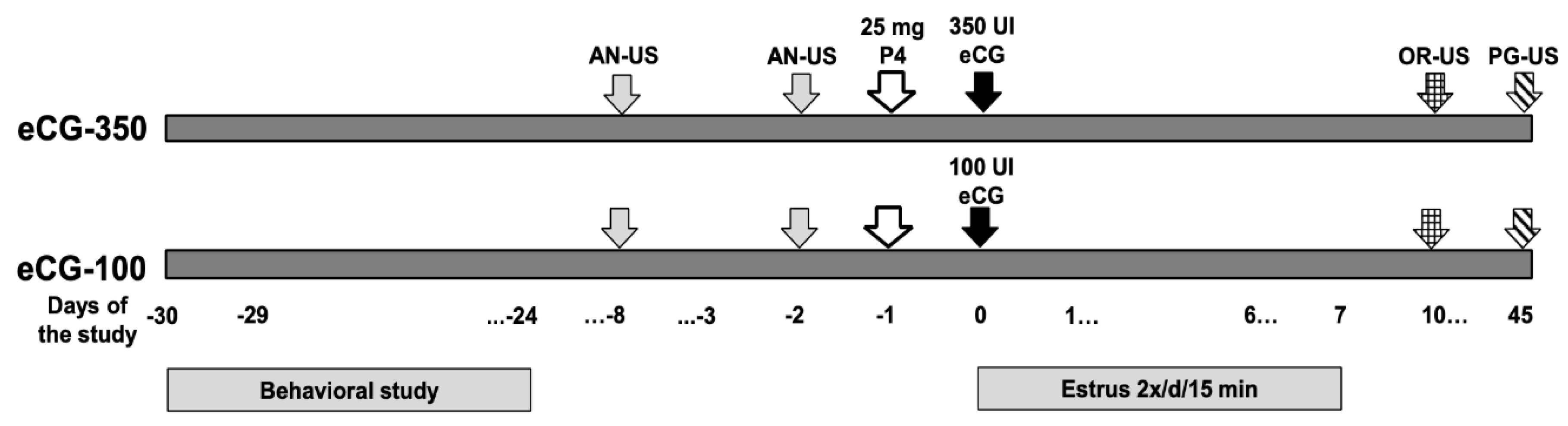
2.3. Behavioral Study, Social Rank, Treatment Groups, Measurements, and Response Variables
2.4. Statistical Analyses
3. Results
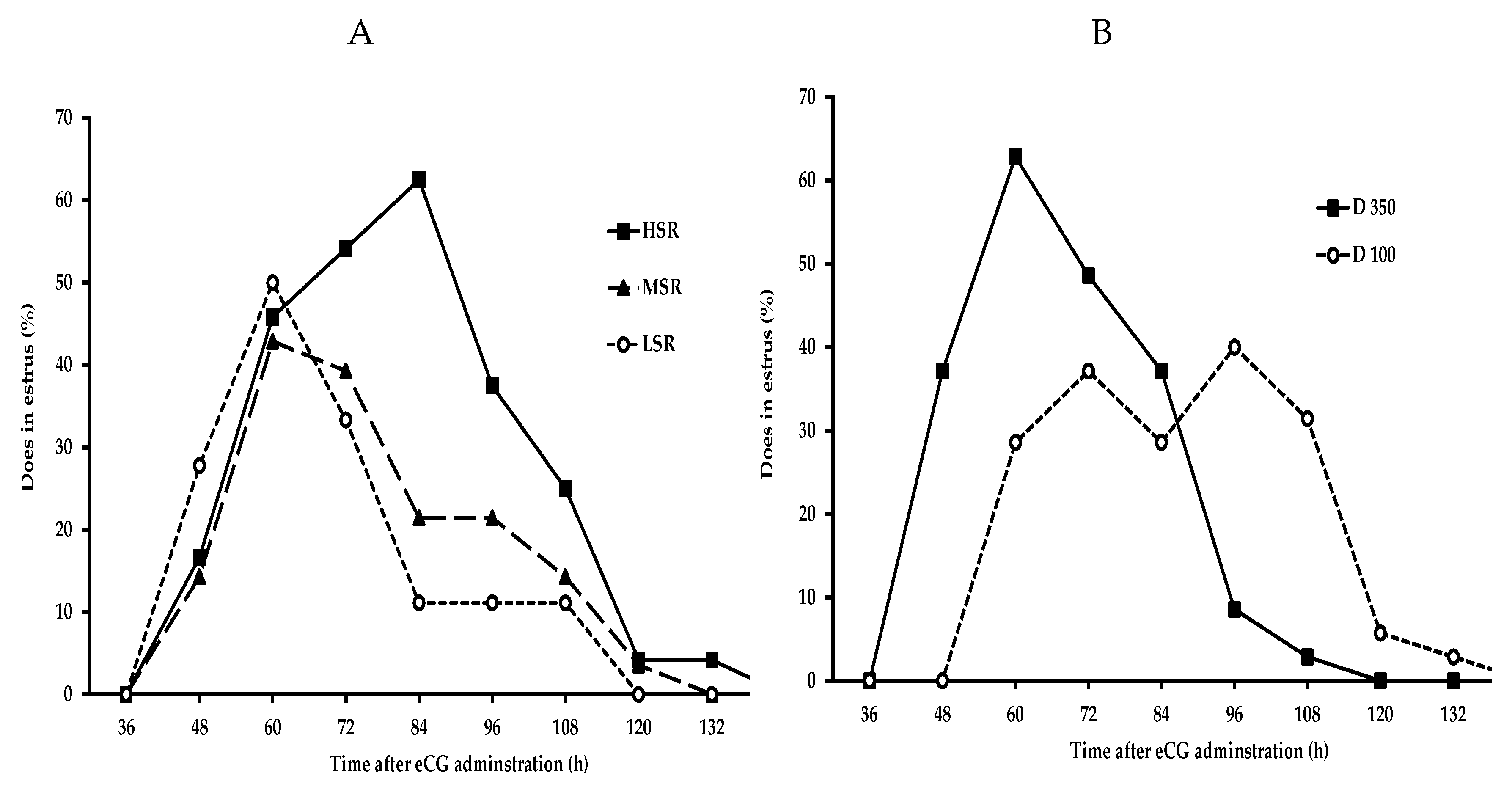
3.1. Effect of Social Rank and the eCG Dose upon the Response Variables
3.2. Effect of Social Rank × eCG Dose Interaction upon the Response Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alvarez, L.; Martin, G.B.; Galindo, F.; Zarco, L.A. Social dominance of female goats affects their response to the male effect. Appl. Anim. Behav. Sci. 2003, 84, 119–126. [Google Scholar] [CrossRef]
- Côté, S.D. Dominance hierarchies in female mountain goats: Stability, aggressiveness and determinants of rank. Behaviour 2000, 137, 1541–1566. [Google Scholar] [CrossRef]
- Fournier, F.; Festa-Bianchet, M. Social dominance in adult female mountain goats. Anim. Behav. 1995, 49, 1449–1459. [Google Scholar] [CrossRef]
- Pelletier, F.; Festa-Bianchet, M. Sexual selection and social rank in bighorn rams. Anim. Behav. 2006, 71, 649–655. [Google Scholar] [CrossRef]
- Ungerfeld, R.; Correa, O. Social dominance of female dairy goats influences the dynamics of gastrointestinal parasite eggs. Appl. Anim. Behav. Sci. 2007, 105, 249–253. [Google Scholar] [CrossRef]
- Aguirre, V.; Orihuela, A.; Vázquez, R. Seasonal variations in sexual behavior, testosterone, testicular size and semen characteristics, as affected by social dominance, of tropical hair rams (Ovis aries). Anim. Sci. J. 2007, 78, 417–423. [Google Scholar] [CrossRef]
- Alvarez, L.; Zarco, L.; Galindo, F.; Blache, D.; Martin, G.B. Social rank and response to the “male effect” in the Australian Cashmere goat. Anim. Reprod. Sci. 2007, 102, 258–266. [Google Scholar] [CrossRef]
- Côte, S.D.; Festa-Bianchet, M. Reproductive success in female mountain goats: The influence of age and social rank. Anim. Behav. 2001, 62, 173–181. [Google Scholar] [CrossRef]
- Santiago-Moreno, J.; Gómez-Brunet, A.; Toledano-Díaz, A.; Pulido-Pastor, A.; López-Sebastián, A. Social dominance and breeding activity in Spanish ibex (Capra pyrenaica) maintained in captivity. Reprod. Fert. Dev. 2006, 19, 436–442. [Google Scholar] [CrossRef]
- Alvarez, L.; Ramos, A.L.; Zarco, L. The ovulatory and LH responses to the male effect in dominant and subordinate goats. Small Rumin. Res. 2009, 83, 29–33. [Google Scholar] [CrossRef]
- Alvarez, L.; Arvizu, R.R.; Luna, J.A.; Zarco, L.A. Social ranking and plasma progesterone levels in goats. Small Rumin. Res. 2010, 90, 161–164. [Google Scholar] [CrossRef]
- Ungerfeld, R.; González-Pensado, S.P.; Dago, A.L.; Vilariño, M.; Menchaca, A. Social dominance of female dairy goats and response to oestrus synchronisation and superovulatory treatments. Appl. Anim. Behav. Sci. 2007, 105, 115–121. [Google Scholar] [CrossRef]
- Chemineau, P.; Malpaux, B.; Brillard, J.P.; Fostier, A. Seasonality of reproduction and production in farm fishes, birds and mammals. Animal 2007, 1, 419–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatet, A.; Pellicer-Rubio, M.T.; Leboeuf, B. Reproductive cycle of goats. Anim. Reprod. Sci. 2011, 124, 211–219. [Google Scholar] [CrossRef]
- Delgadillo, J.A. Environmental and social cues can be used in combination to develop sustainable breeding techniques for goat reproduction in the subtropics. Animal 2011, 5, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Razo, M.A.; Sánchez, F.; Torres-Hernández, G.; Becerril-Pérez, C.; Gallegos-Sánchez, J.; González-Cosío, F.; Meza-Herrera, C.A. Risk factors associated with dairy goats stayability. Livest. Prod. Sci. 2004, 89, 139–146. [Google Scholar]
- Navarrete-Molina, C.; Meza-Herrera, C.A.; Herrera-Machuca, M.A.; Macias-Cruz, U.; Véliz-Deras, F.G. Not all ruminants were created equal: Environmental and socio-economic sustainability of goats under a marginal-extensive production system. J. Clean. Prod. 2020, 255, 120237. [Google Scholar] [CrossRef]
- Simões, J. Recent advances on synchronization of ovulation in goats, out of season, for a more sustainable production. Asian Pac. J. Reprod. 2015, 4, 157–165. [Google Scholar] [CrossRef]
- Contreras-Villarreal, V.; Meza-Herrara, C.A.; Rivas-Muños, R.; Angel-García, O.; Luna-Orozco, J.R.; Carrillo, E.; Mellado, M.; Veliz-Deras, F.G. Reproductive performance of seasonally anovular mixed-bred dairy goats induced to ovulate with a combination of progesterone and eCG or estradiol. Anim. Sci. J. 2016, 87, 750–755. [Google Scholar] [CrossRef]
- Martemucci, G.; D’Alessandro, A.G. Induction/synchronization of oestrus and ovulation in dairy goats with different short term treatments and fixed time intrauterine or exocervical insemination system. Anim. Reprod. Sci. 2011, 126, 187–194. [Google Scholar] [CrossRef]
- Abecia, J.A.; Forcada, F.; González-Bulnes, A. Hormonal control of reproduction in small ruminants. Anim. Reprod. Sci. 2012, 130, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Espino, A.S.; Meza-Herrera, C.A.; Carrillo, E.; González-Álvarez, V.H.; Guillen-Muñoz, J.M.; Ángel-García, O.; Mellado, M.; Véliz-Deras, F.G. Reproductive outcomes of Alpine goats primed with progesterone and treated with human chorionic gonadotropin during the anestrus-to-estrus transition season. Anim. Reprod. Sci. 2016, 167, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martinez, R.; Meza-Herrera, C.A.; Tapia-Robles, K.I.; Alvarado-Espino, A.S.; Luna-Orozco, J.R.; Leyva, C.; Mellado, M.; Veliz-Deras, F.G. Effect of two routes of administration of human chorionic gonadotropin upon estrus induction and reproductive outcomes in adult acyclic mixbreed goats. J. Appl. Anim. Res. 2018, 46, 190–194. [Google Scholar] [CrossRef] [Green Version]
- FASS. Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching, 3rd ed.; Federation Animal Science Society: Champaing, IL, USA, 2010; p. 177. [Google Scholar]
- NAM-National Academy of Medicine. Guide for the Care and Use of Laboratory Animals. Co-Produced by the National Academy of Medicine–Mexico and the Association for Assessment and Accreditation of Laboratory Animal Care International, 1st ed.; Harlan: Mexico City, Mexico, 2010. [Google Scholar]
- Walkden-Brown, S.W.; Restall, B.J.; Scaramuzzy, R.J.; Martin, G.B.; Blackberry, M.A. Seasonality in male Australian cashmere goats: Long term effects of castration and testosterone or oestradiol treatment on changes in LH, FSH and prolactin concentrations, and body growth. Small Rumin. Res. 1997, 26, 239–252. [Google Scholar] [CrossRef]
- Luna-Orozco, J.R.; Guillen-Muñoz, J.M.; De Santiago-Miramontes, M.d.l.A.; García, J.E.; Rodríguez-Martínez, R.; Meza-Herrera, C.A.; Mellado, M.; Véliz, F.G. Influence of sexually inactive bucks subjected to long photoperiod or testosterone on the induction of estrus in anovulatory goats. Trop. Anim. Health Prod. 2012, 44, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Festing, F.W.; Nevalainen, T. The design and statistical analysis of animal experiments: Introduction to this issue. ILAR J. 2014, 55, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Higham, J.P.; Heistermann, M.; Saggau, C.; Agil, M.; Perwitasari-Farajallah, D.; Engelhardt, A. Sexual signaling in female crested macaques and the evolution of primate fertility signals. BMC Evol. Biol. 2012, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Ceacero, F.; García, A.J.; Landete-Castillejos, T.; Bartošová, J.; Bartoš, L.; Gallego, L. Benefits for dominant red deer hinds under a competitive feeding system: Food access behavior, diet and nutrient selection. PLoS ONE 2012, 7, e32780. [Google Scholar] [CrossRef] [Green Version]
- Esposito, L.; de Nicola, D.; Balestrieri, A.; Petrovas, G.; Licitra, F.; Salzano, A.; Neglia, G. Effect of live body weight and method of synchronization on ovulation, pregnancy rate and embryo and fetal loss in buffalo heifers. Anim. Reprod. 2019, 16, 859–863. [Google Scholar] [CrossRef] [Green Version]
- Meza-Herrera, C.A.; Tena-Sempere, M. Interface between Nutrition and Reproduction. In Animal Reproduction in Livestock—Encyclopedia of Life Support Systems; Astiz, S., Gonzalez, A., Eds.; Eolss Publishers: Oxford, UK, 2012. [Google Scholar]
- Urrutia-Morales, J.; Meza-Herrera, C.A.; Tello-Varela, L.; Diaz-Gomez, M.O.; Beltran-Lopez, S. Effect of nutritional supplementation upon pregnancy rates of goats reared under semiarid rangelands and exposed to the male effect. Trop. Anim. Health Prod. 2012, 44, 1473–1477. [Google Scholar] [CrossRef]
- Meza-Herrera, C.A.; Vicente-Pérez, A.; Osorio-Marin, Y.; Giron-Gomez, B.S.; Beltran-Calderon, E.; Avendaño-Reyes, L.; Correa-Calderon, A.; Macias-Cruz, U. Heat stress, divergent nutrition level and late pregancy in hair sheep: Effects upon cotyledon development and litter weight at birth. Trop. Anim. Health Prod. 2015, 47, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Flores-Najera, M.J.; Meza-Herrera, C.A.; Echavarria, F.G.; Villagomez, E.; Iñiguez, L.; Salinas, H.; Gonzalez-Bulnes, A. Influence of nutritional and socio-sexual cues upon reproductive efficiency of goats exposed to the male effect under extensive conditions. Anim. Prod. Sci. 2010, 50, 897–901. [Google Scholar] [CrossRef]
- Barroso, F.G.; Alados, C.L.; Boza, J. Social hierarchy in the domestic goat: Effect on food habits and production. Appl. Anim. Behav. Sci. 2000, 69, 35–53. [Google Scholar] [CrossRef]
- Scaramuzzi, R.J.; Martin, G.B. The importance of interactions among nutrition, seasonality and socio-sexual factors in the development of hormone-free methods for controlling fertility. Repr. Dom. Anim. 2008, 43, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Muñoz, R.; Carrillo, E.; Rodriguez-Martinez, R.; Leyva, C.; Mellado, M.; Véliz, F.G. Effect of body condition score of does and use of bucks. Trop. Anim. Health Prod. 2010, 42, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Meza-Herrera, C.A.; Ross, T.; Hallford, D.; Hawkins, D.; Gonzalez-Bulnes, A. Effects of body condition and protein supplementation on LH secretion and luteal function in sheep. Reprod. Domest. Anim. 2007, 42, 461–465. [Google Scholar] [CrossRef]
- Lopez-Flores, N.M.; Meza-Herrera, C.A.; Perez-Marin, C.; Blache, D.; Arellano-Rodríguez, G.; Zuñiga-Garcia, S.; Navarrete-Molina, C.; Garcia De la Peña, C.; Rosales-Nieto, C.A.; Veliz-Deras, F.G. Precision betacarotene supplementation enhanced ovarian function and the LH release pattern in yearling crossbred anestrus goats. Animals 2020, 10, 659. [Google Scholar] [CrossRef] [Green Version]
- Gamez-Vazquez, H.G.; Rosales-Nieto, C.A.; Bañuelos-Valenzuela, R.; Urrutia-Morales, J.; Diaz-Gomez, M.O.; Silva-Ramos, J.M.; Meza-Herrera, C.A. Body condition score positively influence plasma leptin concentrations in criollo goats. J. Anim. Vet. Adv. 2008, 7, 1237–1240. [Google Scholar]
- Guerra-Garcia, M.; Meza-Herrera, C.A.; Sanchez-Torres-Esqueda, M.T.; Gallegos-Sanchez, J.; Torres-Hernandez, G.; Pro-Martinez, A. IGF-1 and ovarian activity of goats in divergent body condition and supplemented with non-degradable ruminal protein. Agrociencia 2009, 43, 241–247. [Google Scholar]
- Meza-Herrera, C.A.; Torres-Moreno, M.; Lopez-Medrano, J.I.; Gonzalez-Bulnes, A.; Veliz, F.G.; Mellado, M.; Wurzinger, M.; Soto-Sanchez, M.J.; Calderon-Leyva, M.G. Glutamate supply positively affects serum release of triiodothyronine and insulin across time without increases of glucose during the onset of puberty in the female goat. Anim. Reprod. Sci. 2011, 125, 74–80. [Google Scholar] [CrossRef]
- Lopez-Flores, N.M.; Meza-Herrera, C.A.; Galán-Soldevilla, C.; Bautista-Rodriguez, D.A.; Veliz-Deras, F.G.; Arellano-Rodriguez, G.; Garcia-De la Peña, C.; Rosales-Nieto, C.A.; Macias-Cruz, U. The key role of targeted betacarotene supplementation upon endocrine and reproductive outcomes in goats: Follicular development, ovulation rate & the GH-IGF-1 axis. Small Rumin. Res. 2018, 163, 29–33. [Google Scholar]
- Meza-Herrera, C.A.; Veliz-Deras, F.G.; Wurzinger, M.; Lopez-Ariza, B.; Arellano-Rodriguez, G.; Rodriguez-Martinez, R. The kiss-1, kisspeptin, gpr-54 complex: A critical modulator of GnRH neurons during pubertal activation. J. Appl. Biomed. 2010, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Meza-Herrera, C.A.; Gonzalez-Bulnes, A.; Kridli, R.; Mellado, M.; Arechiga-Flores, C.F.; Salinas, H.; Luginbhul, J.M. Neuroendocrine, metabolic and genomic cues signaling the onset of puberty in females. Reprod. Dom. Anim. 2010, 45, e495–e502. [Google Scholar] [CrossRef]
- Meza-Herrera, C.A.; Gonzalez-Velazquez, A.; Veliz-Deras, F.G.; Rodriguez-Martinez, R.; Arellano-Rodriguez, G.; Serradilla, J.M.; Garcia-Martinez, A.; Avendaño-Reyes, L.; Macias-Cruz, U. Short-term glutamate administration positively affects the number of antral follicles and the ovulation rate in cycling adult goats. Reprod. Biol. 2014, 13, 298–301. [Google Scholar] [CrossRef]
- Meza-Herrera, C.A.; Vergara-Hernandez, H.P.; Paleta-Ochoa, A.; Alvarez-Ruiz, A.R.; Veliz-Deras, F.G.; Arellano-Rodriguez, G.; Rosales-Nieto, C.A.; Macias-Cruz, U.; Rodriguez-Martinez, R.; Carrillo, E. Glutamate supply reactivates ovarian function while increases serum insulin and triiodothyronine concentrations in Criollo x Saanen-Alpine yearlings’ goats during the anestrus season. Animals 2020, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Meza-Herrera, C.A.; Pacheco-Alvarez, P.; Castro, O.E.; Macias-Cruz, U.; Avendaño-Reyes, L.; Mellado, M.; Veliz-Deras, F.G.; Contreras-Villarreal, V.; Abad-Zavaleta, J.; Rodriguez-Martinez, R.; et al. Betacarotene supplementation positively affects selected blood metabolites across time around the onset of puberty in goats. Czech. J. Anim. Sci. 2017, 62, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Scaramuzzi, R.J.; Campbell, B.K.; Downing, J.A.; Kendall, N.R.; Khalid, M.; Muñoz-Gutiérrez, M.; Somchit, A. A review of the effects of supplementary nutrition in the ewe on the concentrations of reproductive and metabolic hormones and the mechanisms that regulate folliculogenesis and ovulation rate. Reprod. Nutr. Dev. 2006, 46, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Faria, T.; de Bittencourt Brasil, F.; Sampaio, F.J.; da Fonte Ramos, C. Maternal malnutrition during lactation affects folliculogenesis, gonadotropins, and leptin receptors in adult rats. Nutrition 2010, 26, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Girard-Buttoz, C.; Heistermann, M.; Rahmi, E.; Agil, M.; Ahmad Fauzan, P.; Engelhardt, A. Androgen correlates of male reproductive effort in wild male long-tailed macaques (Macaca fascicularis): A multi-level test of the challenge hypothesis. Physiol. Behav. 2015, 141, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Fritzsche, P.; Riek, M.; Gattermann, R. Effects of social stress on behavior and corpus luteum in female golden hamsters (Mesocricetus auratus). Physiol. Behav. 2000, 68, 625–630. [Google Scholar] [CrossRef]
- Murphy, B.D. Equine chorionic gonadotropin: An enigmatic but essential tool. Anim. Reprod. 2012, 9, 223–230. [Google Scholar]

{kind=link}
{kind=link}
| Behaviors | Social Rank | ||
|---|---|---|---|
| LSR | MSR | HSR | |
| Threats | 25 ± 5.6 | 106 ± 8.7 | 231 ± 19.5 |
| Bumps | 6 ± 1.4 | 27 ± 3.6 | 46 ± 7.7 |
| Shoves | 2 ± 0.5 | 5 ± 1.0 | 13 ± 3.2 |
| Chases | 0 ± 0 | 0 ± 0 | 1 ± 0.5 |
| Evasions | 345 ± 48.1 | 125 ± 11.1 | 67 ± 8.9 |
| Escapes | 1 ± 0.4 | 0 ± 0 | 0 ± 0 |
| Success Index 1 | 0–0.33 | 0.34–0.63 | 0.63–1.0 |
| Variables | Social Rank (R) | eCG Dose (D) | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| LSR | MSR | HSR | 100 | 350 | R | D | R × D | |
| LW (kg) | 31.6 ± 1.6 b | 44.0 ± 1.3 a | 49.0 ± 1.4 a | 41.9 ± 1.5 a | 41.8 ± 1.4 a | 0.001 | 0.615 | 0.001 |
| BC (units) | 1.8 ± 0.07 a | 1.9 ± 0.06 a | 1.9 ± 0.06 a | 1.9 ± 0.05 a | 1.9 ± 0.05 a | 0.768 | 0.733 | 0.978 |
| EI (%) | 10/18 (56) b | 20/28 (71) b | 23/24 (96) a | 26/35 (74) a | 27/35 (77) a | 0.007 | 0.599 | 0.019 |
| LAT (h) | 57.6 ± 4.6 a | 69.6 ± 3.2 a | 68.3 ± 3.0 a | 76.1 ± 3.0 b | 57.7 ± 2.8 a | 0.373 | 0.001 | 0.001 |
| DUR (h) | 17.3 ± 4.0 b | 18.9 ± 3.2 b | 29.0 ± 3.5 a | 20.2 ± 2.9 a | 23.6 ± 2.9 a | 0.004 | 0.313 | 0.051 |
| OVU (%) | 14/18 b (78) | 26/28 ab (93) | 23/24 a (96) | 30/35 a (86) | 33/35 a (94) | 0.05 | 0.177 | 0.294 |
| OR (n) | 1.27 ± 0.17 a | 1.77 ± 0.13 a | 1.58 ± 0.14 a | 1.09 ± 0.10 b | 2.06 ± 0.10 a | 0.079 | 0.001 | 0.001 |
| CLS (cm) | 0.8 ± 0.09 b | 1.05 ± 0.07 a | 1.04 ± 0.07 a | 0.91 ± 0.06 a | 1.05 ± 0.06 a | 0.046 | 0.08 | 0.13 |
| PREG (%) | 9/18 a (50) | 14/28 a (50) | 14/24 a (58) | 16/35 a (46) | 21/35 a (60) | 0.651 | 0.147 | 0.037 |
| KIDD (%) | 5/18 a (28) | 10/28 a (36) | 12/24 a (50) | 11/35 a (31) | 16/35 a (46) | 0.279 | 0.184 | 0.119 |
| LS (n) | 1.60 ± 0.4 a | 1.80 ± 0.3 a | 1.83± 0.2 a | 1.36 ± 0.2 b | 2.06 ± 0.2 a | 0.228 | 0.023 | 0.139 |
| Variables | eCG-100 | eCG-350 | ||||
|---|---|---|---|---|---|---|
| LSR | MSR | HSR | LSR | MSR | HSR | |
| LW (kg) | 30.0 ± 2.4 b | 43.4 ± 1.8 a | 47.6 ± 1.9 a | 33.0 ± 2.1 b | 44.7 ± 1.8 a | 47.4 ± 2.1 a |
| EI (%) | 3/8 c (38) | 11/14 ab (79) | 12/13 ab (92) | 7/10 ab (70) | 9/14 bc (64) | 11/11 a (100) |
| LAT (h) | 72.0 ± 8.4 abc | 79.6 ± 4.4 a | 74.0 ± 4.2 ab | 51.4 ± 5.5 d | 57.3 ± 4.8 cd | 62.2 ± 4.4 bcd |
| DUR (h) | 7.5 ± 5.9 b | 18.9 ± 4.5 ab | 29.5 ± 4.6 a | 25.2 ± 5.3 a | 18.9 ± 4.5 ab | 28.4 ± 5.0 a |
| OR (n) | 0.86 ± 0.3 d | 1.2 ± 0.2 cd | 1.2 ± 0.2 cd | 1.6 ± 0.2 bc | 2.3 ± 0.2 a | 2.1 ± 0.2 ab |
| PREG (%) | 3/8 b (38) | 9/14 ab (64) | 4/13 b (31) | 6/10 ab (60) | 5/14 b (36) | 10/11 a (91) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuñiga-Garcia, S.; Meza-Herrera, C.A.; Mendoza-Cortina, A.; Otal-Salaverri, J.; Perez-Marin, C.; Lopez-Flores, N.M.; Carrillo, E.; Calderon-Leyva, G.; Gutierrez-Guzman, U.N.; Veliz-Deras, F.G. Effect of Social Rank upon Estrus Induction and Some Reproductive Outcomes in Anestrus Goats Treated With Progesterone + eCG. Animals 2020, 10, 1125. https://doi.org/10.3390/ani10071125
Zuñiga-Garcia S, Meza-Herrera CA, Mendoza-Cortina A, Otal-Salaverri J, Perez-Marin C, Lopez-Flores NM, Carrillo E, Calderon-Leyva G, Gutierrez-Guzman UN, Veliz-Deras FG. Effect of Social Rank upon Estrus Induction and Some Reproductive Outcomes in Anestrus Goats Treated With Progesterone + eCG. Animals. 2020; 10(7):1125. https://doi.org/10.3390/ani10071125
Chicago/Turabian StyleZuñiga-Garcia, Santiago, Cesar A. Meza-Herrera, Adela Mendoza-Cortina, Julio Otal-Salaverri, Carlos Perez-Marin, Noé M. Lopez-Flores, Evaristo Carrillo, Guadalupe Calderon-Leyva, Ulises N. Gutierrez-Guzman, and Francisco G. Veliz-Deras. 2020. "Effect of Social Rank upon Estrus Induction and Some Reproductive Outcomes in Anestrus Goats Treated With Progesterone + eCG" Animals 10, no. 7: 1125. https://doi.org/10.3390/ani10071125