1. Introduction
The pullet rearing environment, which constitutes the first 16–18 weeks of life, can have long-term effects on laying hen welfare [
1]. Studies of pullet housing have shown that the rearing environment affects cognition, behavior, as well as bone and muscle development. For example, compared to hens reared in non-enriched cages, aviary-reared hens show improved working memory in the reversal phase of a holeboard task [
2]. Birds reared with access to vertical space use perches and elevated platforms more at 19 weeks of age than birds reared without elevated structures [
3,
4]. Aviary-reared pullets have stronger, more developed radius, humerus, and tibia bones, a larger keel bone, and heavier wing and breast muscles compared to pullets from non-enriched cages at 16 weeks of age [
5]. Importantly, these effects on the skeletal system persist through to the end of the laying period. Aviary-reared hens have stronger bones with greater cross-sectional areas and higher mineral content compared to hens reared in non-enriched cages at 73 weeks of age [
6].
Rearing environments have also been shown to affect keel bone fracture development. Hens housed in enriched colony cages had a higher incidence of keel bone fractures at the end of the laying period if they were reared in non-enriched cages versus aviaries [
7]. The keel bone is crucial for flight and general mobility as it serves as an attachment site for wing muscles [
8]. Birds with fractured keels may experience pain [
9], reduced mobility [
9,
10], and reduced egg production [
11,
12]. Keel bone fractures are suspected to be partly attributed to physical trauma to the bone [
13,
14]. Accordingly, previous research found the number of collisions that a hen experienced corresponded with the risk of keel fracture development [
15,
16]. Using tri-axial accelerometers, the potential causes of such trauma were examined [
17]. The keel-mounted sensors registered acceleration events experienced at the keel by hens as they navigated their enriched colony cage environments. Collisions with cage structures, and specifically the perch, caused most (~80%) of the acceleration events. However, the impact of the rearing environment on collisions was not explored.
The reported influence of rearing on keel bone fracture prevalence [
7] may reflect differences in the birds’ abilities to navigate their adult environments. Therefore, the objective of this study was to investigate whether the early rearing environment (non-enriched cage vs. aviary housing) affects the behavior of hens after they are placed in their layer environment. Specifically, we aimed to determine whether pullet rearing affects the incidence of acceleration events recorded at the keel, and the behaviors associated with these events. We further explored actions and housing structures associated with observed collisions. Considering that the perch was the main structure associated with collisions in a previous study [
17], we also investigated the effect of rearing on perching behavior focusing on perch use and the distribution of birds across the two available perches.
2. Materials and Methods
2.1. Ethical Statement
All procedures were approved by the California Polytechnic State University Animal Care and Use Committee (Protocol #1613).
2.2. Animals
On farm data collection was conducted at the Cal Poly Poultry Center at California Polytechnic State University. A total of 301 pullets were used in this study. The pullets were selected from among 3000 day-old Lohmann LSL-Lite chicks, with infra-red beak trim, that were obtained from Hy-Line North America, LLC (Burley, ID, USA). All pullets were from the same parent flock.
2.3. Pullet Housing and Management
A total of 2000 pullets were reared in a conventional non-enriched cage system (CON; 98 × 69.5 × 41 cm, L × W × H; SALMET S1000 Rearing System, SALMET GmbH & Co. KG, Dietzenbach, Germany). The wire floored cages were stacked in three tiers of 36 cages/tier. All pullets were initially housed in the middle tier at 55–60 pullets/cage with a space allowance of 113.5–123.8 cm2/pullet. Space per bird was increased during wk 6, at which time pullets were sorted among cages in the top and middle tiers. The pullets were redistributed among all three tiers at 12 weeks of age, with 20 pullets/cage at 340.6 cm2/pullet. Water was provided ad libitum via automatic water lines (Ziggity, Middlebury, IN, USA) with 4 nipple drinkers/cage. An automated trough feeder provided hens with ad libitum access to a commercial diet.
A total of 1000 pullets were reared in a multi-tier aviary system (AVI; 16.5 × 4.7 × 2.5 m, L × W × H; SALMET Pedigrow 2 Rearing System, SALMET GmbH & Co. KG, Dietzenbach, Germany). The system contained three elevated tiers at 0.3 m, 1.2 m, and 1.6 m height from the floor. Pullets were enclosed into the middle tier for the first three weeks (564.9 cm2/pullet). After the first three weeks, tier doors were opened and pullets could access the bottom and top tiers of the system, shelves on the outer edge of the middle and top tiers (147 × 18 cm/shelf, L × W), and a scratch area adjacent to the system containing pine wood shavings over a concrete floor (514,000 cm2 scratch area). Considering all sections, shelves, and adjacent scratch area, the total available floor space was 2267 cm2/pullet for the remainder of the rearing period.
Each tier in AVI contained a wire mesh floor, manure belt, automatic water line, automatic feeder line, and square metal perch (2.5 cm diameter) above the feeder. The top tier contained an additional 11 square metal perches (27.7 cm/pullet of linear perch space). Wire mesh ramps were provided to facilitate access from the floor to the middle tier. Water was provided ad libitum with 132 nipple drinkers/tier (Ziggity). AVI chicks were fed the same commercial diet as CON chicks.
For both rearing environments, temperature and artificial lighting were maintained according to the North American Edition of the LSL-Lite Layers Management Guide (Lohmann Tierzucht GmbH, Cuxhaven, Germany).
2.4. Layer Housing and Management
At 19 weeks of age, all birds were moved into their adult housing system—a two-story barn containing 128 enriched colony cages (ECC; 64 cages/floor; 25 ± 2 hens/ECC). Birds were placed in ECC with other birds from the same rearing treatment, and treatments were distributed randomly across each story of the barn. A total of 12 ECC were chosen as focal cages (n = 6 ECC/rearing treatment). Each ECC (360 × 63 × 56 cm, L × W × H; SALMET AGK 3600 Enrichable Colony System, SALMET GmbH & Co.) contained two elevated round metal perches (9 and 25 cm from cage floor) that were each 225 cm in length, as well as an enclosed nest area (51 × 30 × 52 cm, L × W × H) and scratch pad (35 × 13 cm, L × W) over a wire mesh floor. The hens had access to 907.2 cm2/hen of floor space and 18 cm/hen of perch space. Water was provided ad libitum with 8 nipple drinkers/cage (Ziggity). A commercial diet was provided ad libitum in automatic feeders which ran along each cage row. The artificial lighting program started at 12 h/d at initial placement and was subsequently stepped up by 1 h/week to reach 16 h/day.
2.5. Behaviors Associated with Acceleration Events at the Keel
At 21, 35, and 49 weeks, three randomly selected focal hens/ECC (36 total hens/age) were fitted with a custom jacket containing a tri-axial accelerometer with a sensor positioned over the keel bone (Custom Idea Ltd., Shepton Mallet, UK; for more information on jackets and accelerometers, see [
17]). A different set of focal hens was selected at each age. Jackets were fitted to hens during nighttime hours in order to minimize hen disturbance. Each white jacket contained a unique black symbol, which allowed us to individually identify the focal hens during recording. Jackets and accelerometers were worn continuously for 11 d at each age. Hens habituated to the jackets during the first day, and data were recorded for the remaining 10 d. Accelerometers were programmed to record events at the keel with a combined magnitude of acceleration >12 G-Units (G) to avoid the inclusion of minor body movements [
17]. The combined magnitude of acceleration was calculated as the square root of the sum of squared acceleration in three dimensions (X, Y, and Z) [
17]. At the conclusion of each age trial, a log containing date, time, and the maximum combined magnitude of acceleration for each event was generated for each hen. This log was used for video analysis.
ECC were video recorded continuously during each of the three 10 d trials using two video cameras per cage (Veilux 700 Line Day Night IR Bullet Camera, Veilux Inc., Grand Prairie, TX, USA) connected to a digital video recorder with surveillance software (GeoVision Digital Surveillance System version 8.5, GeoVision Inc., Taipei, Taiwan). After the end of each age trial, trained observers used the accelerometer log with the video recordings to identify the behavior associated with each acceleration event (
Table 1) [
17]. Observers were blind to rearing treatment, randomly assigned to video from both rearing treatments, and analyzed all videos using GeoVision Software (GeoVision Digital Surveillance System, ViewLogEx v.8.5.6.0, GeoVision Inc., Tapei, Taiwan). Inter-observer reliability was high (≥90% inter-observer agreement between trainer and each trainee of approximately 20 acceleration events).
2.6. Perching Behavior
Hen perching behavior was evaluated from video on the second and sixth day of each 10-d trial. Using instantaneous scan sampling with 5-min intervals, the number of hens using the low and high perch were observed. It was previously reported that 5-min interval sampling was representative of perching behavior throughout a 15 h day [
18]. A total of five hours of video was reviewed on each day: three hours while the house lights were on (06:00–06:55, 12:00–12:55, 16:00–16:55; “day”) and two hours while lights were off (23:00–23:55, 02:00–02:55; “night”). Preliminary analysis (a review of 24 h of video data for five ECC) found this sampling scheme to be representative of the entire day for perching behavior.
2.7. Data Analysis
For all data analyses, R statistical software (version 3.5.2, R Core Team, Vienna, Austria) [
19] and linear mixed effect models (lme4 package) [
20] were used. Model assumptions were checked for normality of residuals by visually evaluating Q–Q plots, residual vs. fitted values plots, and homoscedasticity of independent variables. As models did not meet one or more normality criteria, generalized linear mixed effect models with a Poisson distribution (lme4 package) [
20] were used to analyze the total number of acceleration events and total number of collisions sustained by each hen across the 10 d at each age. Model assumptions for generalized linear mixed effects models were checked using package DHARMa [
21]. Following a previous study [
17], only acceleration events and collisions associated with accelerations >20 G were included in the statistical analyses. The effect of treatment (factor with 2 levels: CON, AVI), age (factor with 3 levels: 21–22, 35–36, and 49–50 weeks), day/night (factor with 2 levels: day, night), and their interactions (treatment × age, treatment × day/night, age × day/night, treatment × age × day/night) were assessed, with hen ID nested in age nested in ECC as a random effect. The final models were obtained by a stepwise backwards reduction using ANOVA for model comparison with a
p-value of >0.05 as the criterion of exclusion.
Within each 5-min observation, we calculated the average proportion of hens from each ECC that utilized either of the two perches. The numerator was the total number of hens on the low perch and high perch, and the denominator was the total number of hens in the ECC. The data were analyzed using a zero-inflated beta regression model (glmmTMB package) [
22]. The effect of treatment (factor with 2 levels: CON, AVI), age (factor with 3 levels: 21–22, 35–36, and 49–50 weeks), day/night (factor with 2 levels: day, night), and their interactions (treatment × age, treatment × day/night, age × day/night, treatment × age × day/night) were assessed, with day nested in age nested in cage as a random effect.
We also calculated the average proportion of perching hens from each ECC that utilized the high perch during nighttime hours. The numerator was the total number of hens on the high perch, and the denominator was the total number of hens on the low perch and high perch. The same zero-inflated beta regression model described previously was used, excluding the effect of light.
4. Discussion
The objective of this study was to determine whether pullet rearing conditions affect the incidence of acceleration events and collisions at the keel, as well as perching behavior, for ECC housed laying hens. We predicted that AVI hens would sustain fewer acceleration events and fewer collisions than CON hens, and we predicted that AVI hens would perch more overall, and specifically on the high perch, than CON hens. We found that both AVI and CON hens experienced fewer acceleration events at the keel and collided with features of their environment less frequently as they aged. However, AVI hens sustained fewer acceleration events and fewer collisions than CON hens at initial and peak phase of the laying period (21 and 35 weeks of age), suggesting that AVI hens were navigating their laying environment with more ease from an earlier age than CON hens.
Previous research has demonstrated that rearing environments contribute to learning ability, such that birds reared in enriched environments learn faster than birds reared in non-enriched environments. Floor-reared pullets with one week of exposure to outdoor range were faster to learn a Y-maze task compared to floor-reared pullets with no other structural or sensory enrichment [
23]. Similarly, floor-reared pullets with three weeks of sensory enrichment were faster to learn a T-maze task than floor-reared pullets without enrichment [
24]. During the reversal phase of a holeboard task, pullets reared in a multi-tier aviary for 12 weeks were faster to complete the task than pullets reared in a conventional non-enriched cage for 16 weeks [
2]. These previous findings suggest that enriched (AVI) rearing environments improve birds’ abilities to learn compared to non-enriched (CON) rearing environments, which may have contributed to AVI hens experiencing fewer acceleration events and collisions than CON hens at earlier time points. Cognitive testing was not performed in our experiment, but our findings indicate that more cognitive tests and potential brain analyses are needed to assess the effect of rearing on long-term learning in novel environments.
It is worth noting that the magnitude of difference between rearing treatments’ estimated means was low, with CON hens experiencing about 1–2 acceleration events/hen/10 day and about 1 collision/hen/10 day more than AVI hens at initial and peak lay. However, assuming that these numbers remained consistent in the 14 weeks between initial and peak lay, CON hens could have cumulatively experienced up to 10 collisions more per hen than AVI hens between 21 to 35 weeks of age. Previous work found that birds with a higher number of collisions were at an increased risk for sustaining keel bone fractures [
15,
16], and other researchers reported that CON hens have a higher incidence of keel bone fractures than AVI hens at 73 weeks of age in ECC [
7]. Therefore, considering our data with these previous two studies, it is possible that CON hens cumulatively experience more collisions and thus are at a greater risk for injury. Future research should evaluate the effect of rearing on both collisions and keel bone fractures concurrently for further evaluation. As mentioned, the number of collisions experienced across the 10-d trials could be considered low, with estimated means ranging from 0–2 collisions/hen/10 day >20 G across rearing treatments and ages during the day. Our study found fewer collisions on average than previous work using the same accelerometers, which evaluated the number of collisions per hen across two 3-week timepoints/ECC [
17]. Baker et al. [
17] reported an average of 12.0 and 12.6 collisions/hen/3 weeks >20 G during the day (estimated 5.7 and 6.0 collisions/hen/10 d). The hens used in this previous work were floor-reared, older, of a different strain (Hy-line W36), and housed in ECC with a different spatial layout of perches than those in our study. Some or all of these factors could have contributed to the discrepancy in the number of collisions reported in our study and the previous one [
17]. Overall, it is important to consider that acceleration events at the keel are experienced by individuals. The spread of the interquartile range of acceleration event data, particularly at 35 weeks of age (
Figure 2), suggests that some CON individuals experienced more acceleration events than the majority of AVI hens, which could further increase their risk for keel bone fractures during peak lay.
Collisions constituted the main cause of acceleration events experienced at the keel bone for initial and peak lay. This finding confirms previous results obtained using the same accelerometers that collisions constitute a majority of acceleration events >20 G [
17]. However, the number of collisions remained consistent over two age groups (52–60 and 74–83 weeks) in the previous results [
17], whereas the collisions in our study decreased as birds aged. Other researchers have also found hens to experience fewer collisions with increasing age [
14,
25]. The inverse relationship between collisions and age has been attributed to hens being the most active earlier in the laying period and reducing activity over time [
25,
26,
27,
28]. An alternative explanation for our study could be that hens from both rearing treatments were experiencing a novel environment at initial lay. Neither of the rearing environments matched the laying environment in terms of group size, space allowance, and the design and spatial layout of physical structures. A mismatch between the rearing and laying environment is attributed to behavioral challenges when transitioning into the laying environment (e.g., aggression, space perception) [
1]. As a result, having the most acceleration events and collisions at the initial phase of the laying period could also be attributed to adapting to a novel environment.
We descriptively evaluated the objects that birds collided with, as well as the actions that birds performed at the time of collision, to provide further detail into how birds utilized their space. Collisions with either of the perches constituted the greatest proportion of collision objects for both treatments (63.3% of collisions with low perch and high perch for CON hens vs. 57.7% of collisions with low perch and high perch for AVI hens). These findings are in agreement with previous research [
17], which also found perches to be the main objects that birds collided with in ECC (71.5% of collisions with perches). The authors noted that collisions with perches occurred mainly as hens ascended onto them [
17]. Along with slipping while perching, ascending onto perch was also among the top two actions associated with perch collisions by AVI hens in this study. Meanwhile, the top two collision action categories for CON hens were running into an object or a conspecific while locomoting across the floor and slipping on a perch.
We predicted that more AVI hens would be observed perching than CON hens at all ages. If true, this finding would explain why perch ascent may have been a leading source of collisions for AVI hens but not CON hens. Previous research found that pullets reared with access to vertical space utilized perches and elevated platforms more than birds reared without perches [
3,
4]. However, in our study, the total proportion of birds perching was similar across rearing treatments and for most time points. Although CON hens were not raised with perches, they may have perched on the water lines as pullets, offering one possible explanation for the similar perch use, including during the earliest observation time point. It is also possible that the two weeks that the birds spent in the ECC system before the data collection at 21 weeks of age provided the chicks with sufficient time to learn to use the perches. This is particularly likely since the low perch was located only 9 cm from the cage floor and ran the length of the cage, making it easily accessible for the birds.
Daytime perch use was low, with only 1–8% of birds observed perching, regardless of perch height or rearing treatment. A previous study reported a range of 17–30% of birds perching during lights on in groups of 33–50 hens per ECC [
29]. Others reported birds spending 7–25% of their time perching during the day in groups of 14 hens per ECC [
30]. In these studies, daytime perch use was attributed to short bouts of grooming, standing, sleeping, and facilitating access to another resource (e.g., water line). In our study, the perches did not facilitate access to feed or water, so it is possible that they were utilized less during the daytime while birds were active and using other resources in the ECC. On the other hand, laying hens are highly motivated to access perches at night for roosting, which is attributed to an antipredator defense strategy [
31]. At least one-third of birds, regardless of rearing treatment, were observed roosting on perches at night at all three ages in our study. The remaining proportion of birds were commonly observed huddled together in one or two large groups on the floor of the ECC. The proportion of roosting birds in our study is lower than reported in several previous studies of nighttime perching among hens housed in enriched cages (53–72% of hens roosting [
29]; 85–100% of hens roosting [
32]). However, other researchers have reported that the majority of hens in ECC sleep huddled on the floor during lights off as opposed to roosting on perches [
33]. Birds in our study had sufficient perch space for all hens to be perching simultaneously on either perch (at least 15 cm per hen [
32]), but possibly different designs and layout of perches in ECC contributed to the behavioral differences in our study compared to others.
We evaluated perching birds’ preference for the low versus high perch. Given the low daytime perch use observed, our analysis focused on nighttime observations. While both AVI and CON hens utilized perches at night, more AVI hens used the high perch. Previous studies that evaluated the effect of rearing on perch and platform use collected data only at 16 and 19 weeks of age [
3,
4]. Our work demonstrates that rearing influences height preference into mid-lay (49 weeks of age). It is not clear from our study whether the influence of rearing on roost height preference reflects differences in spatial cognition, perception, learning, or physical abilities. It is also unclear whether it was the height or other aspects of perch design that were preferred by the AVI birds, or that may have dissuaded CON birds from mounting it. Unlike the low perch, the high perch in this study was suspended from the cage ceiling and consequently could sway back and forth. This instability of the perch could have impacted the birds’ preference for it.
In addition to perch use, the lighting period, whether lights were on or off, also had an effect on collisions. All birds, regardless of rearing treatment, experienced more collisions during the day versus during the night. Similarly, previous studies found that the majority of collisions took place during lights on, and specifically during the dusk and dawn phases when hens were moving to and from roosting [
17,
25]. In our study, we did not observe a pattern of collisions during dawn and dusk specifically. Instead, the higher prevalence of collisions during lights on could be attributed to hens being generally more active during the day than during the night. In laying hens, more illuminance promotes more physical activity [
34], and, as a result, hens have more opportunities for collisions during lights on. Furthermore, the lights on period was twice as long as the lights off period in our study (16 h vs. 8 h, respectively), so hens also had more total time to sustain collisions during the lights on period as opposed to lights off.
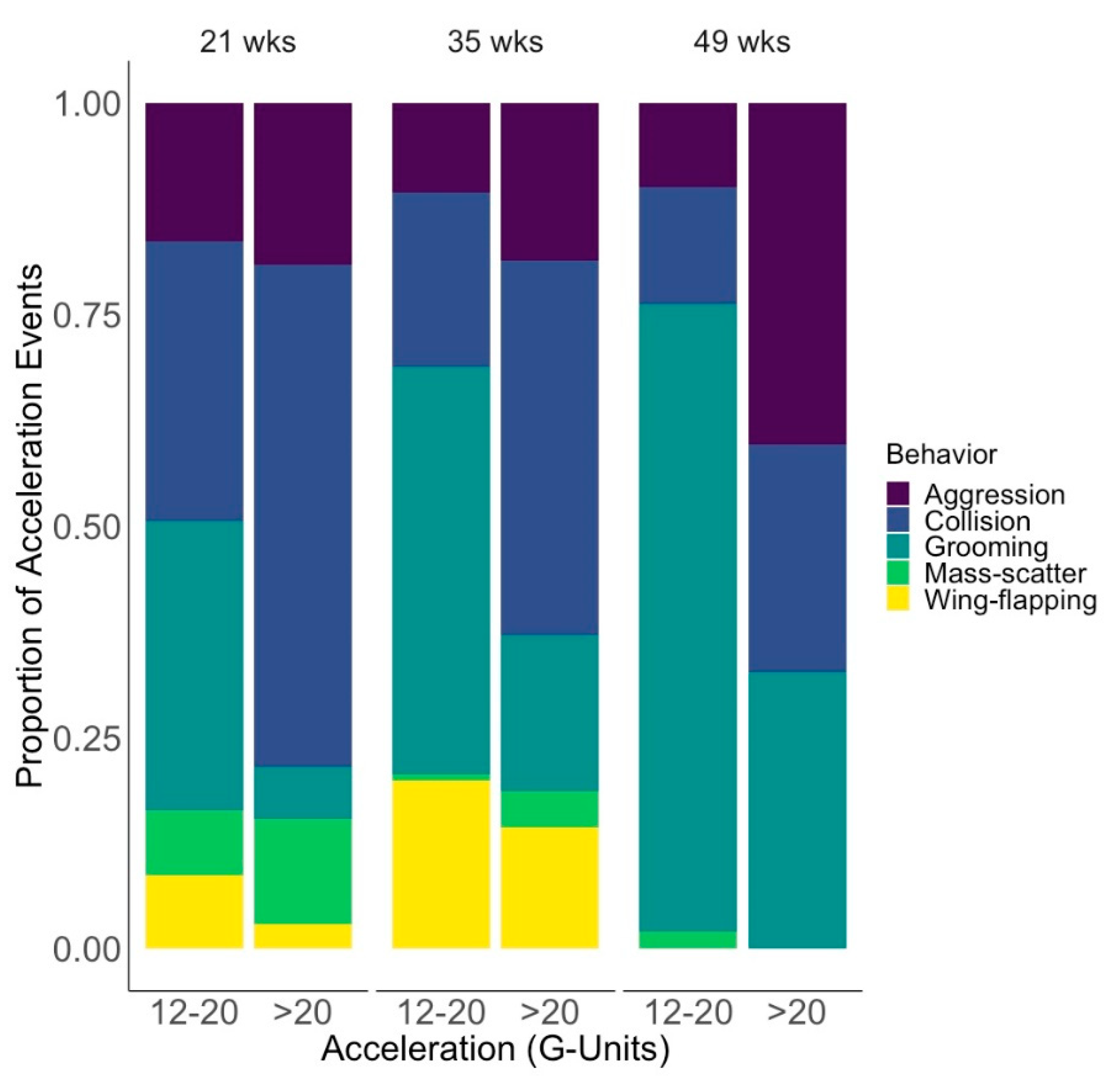
Lastly, we found social interactions to be an important contributing factor to keel acceleration events, such that aggression was the second-most cause of acceleration events in another study and in our study [
17]. Coupled with the fact that a notable collision object category was conspecifics and a notable collision action category was birds being pushed by conspecifics, these findings warrant further investigation into the relationship of social behavior and collisions. In the context of early experiences, it is important to consider that rearing environments differ in the group sizes allowance, availability of space for hiding or retreat, and other aspects that influence social experiences and development of chicks. Research into the interactions between rearing, social behavior, space use, and collisions is needed.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}