Effects of Different Dietary Vegetable Lipid Sources on Health Status in Nile Tilapia (Oreochromis niloticus): Haematological Indices, Immune Response Parameters and Plasma Proteome


 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Diet Formulation and Fish Culture
2.2. Fish Sampling and Blood Collection
2.3. Haematological and Immune Assays
2.4. Plasma Proteome Analysis
2.4.1. Protein Preparation, Labelling and Two-Dimension Gel Electrophoresis
2.4.2. Gel Image Acquisition and Analysis and Protein Identification by MALDI-TOF/TOF
2.5. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
2.5.1. Molecular Cloning of Genes
2.5.2. qRT-PCR Analysis Gene Expression
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost. Rev. Fish Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- De Pablo, M.A.; Álvarez De Cienfuegos, G.A. Modulatory effects of dietary lipids on immune system functions. Immunol. Cell Biol 2000, 78, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Arts, M.T.; Kohler, C.C. Health and condition in fish: The influence of lipids on membrane competency and immune response. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M.J., Eds.; Springer: New York, NY, USA, 2009; pp. 237–256. ISBN 978-0-387-89366-2. [Google Scholar]
- Dennis, E.A.; Norris, P.C. Eicosanoid Storm in Infection and Inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildirim-Aksoy, M.; Lim, C.; Davis, D.A.; Shelby, R.; Klesius, P.H. Influence of Dietary Lipid Sources on the Growth Performance, Immune Response and Resistance of Nile Tilapia, Oreochromis niloticus, to Streptococcus iniae Challenge. J. Appl. Aquac. 2007, 19, 29–49. [Google Scholar] [CrossRef]
- Lim, C.; Yildirim-Aksoy, M.; Li, M.H.; Welker, T.L.; Klesius, P.H. Influence of dietary levels of lipid and vitamin E on growth and resistance of Nile tilapia to Streptococcus iniae challenge. Aquaculture 2009, 298, 76–82. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V. Mechanisms of Oxidative Processes in Meat and Toxicity Induced by Postprandial Degradation Products: A Review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 96–123. [Google Scholar] [CrossRef]
- Miller, G.J. Effects of diet composition on coagulation pathways. Am. J. Clin. Nutr. 1998, 67, 542S–545S. [Google Scholar] [CrossRef] [Green Version]
- Rakocy, J.E. Cultured Aquatic Species Information Programme. Oreochromis niloticus. Cultured Aquatic Species Information Programme; FAO Fisheries and Aquaculture Department: Rome, Italy, 18 February 2005; [Cited 27 June 2020]. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2018 - Meeting the sustainable development goals. Rome. Licence: CC BY-NC-SA 3.0 IGO.
- Lim, C.; Yildirim-Aksoy, M.; Klesius, P.H. Lipid, Fatty Acid Requirements of Tilapia. N. Am. J. Aquac. 2011, 73, 188–193. [Google Scholar] [CrossRef]
- Olsen, R.E.; Henderson, R.J.; McAndrew, B.J. The conversion of linoleic acid and linolenic acid to longer chain polyunsaturated fatty acids by Tilapia (Oreochromis) nilotica in vivo. Fish Physiol. Biochem. 1990, 8, 261–270. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Tanomman, S.; Ketudat-Cairns, M.; Jangprai, A.; Boonanuntanasarn, S. Characterization of fatty acid delta-6 desaturase gene in Nile tilapia and heterogenous expression in Saccharomyces cerevisiae. Comp. Biochem. Physiol. Part B 2013, 166, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Torrecillas, S.; Benítez-Dorta, V.; Caballero, M.J.; Izquierdo, M.S.; Zamorano, M.J. Effects of dietary vegetable oils on the expression of eicosanoid receptors genes in Senegalese sole (Solea senegalensis) intestine. Aquaculture Rep. 2019, 15, 100201. [Google Scholar] [CrossRef]
- Turkmen, S.; Perera, E.; Zamorano, M.J.; Simó-Mirabet, P.; Xu, H.; Pérez-S#xE1;nchez, J.; Izquierdo, M. Effects of Dietary Lipid Composition and Fatty Acid Desaturase 2 Expression in Broodstock Gilthead Sea Bream on Lipid Metabolism-Related Genes and Methylation of the fads2 Gene Promoter in Their Offspring. Int. J. Mol. Sci. 2019, 20, 6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, N.; Silva, T.S.; Wulff, T.; Schrama, D.; Dias, J.P.; Rodrigues, P.M.L.; Conceicao, L.E.C. Nutritional mitigation of winter thermal stress in gilthead seabream: Associated metabolic pathways and potential indicators of nutritional state. J. Proteomics 2016, 142, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Boonanuntanasarn, S.; Nakharuthai, C.; Schrama, D.; Duangkaewa, R.; Rodrigues, P.M. Effects of dietary lipid sources on hepatic nutritive contents, fatty acid composition and proteome of Nile tilapia (Oreochromis niloticus). J. Proteomics 2019, 192, 208–222. [Google Scholar] [CrossRef]
- Schrama, D.; Cerqueira, M.; Raposo, C.S.; da Costa, A.M.R.; Wulff, T.; Gonçalves, A.; Camacho, C.; Colen, R.; Fonseca, F.; Rodrigues, P.M. Dietary Creatine Supplementation in Gilthead Seabream (Sparus aurata): Comparative Proteomics Analysis on Fish Allergens, Muscle Quality, and Liver. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- AOAC (Ed.) AOAC Official methods of Analysis. In Association of Official Analytical Chemists, 14th ed.; AOAC: Arlington, VA, USA, 1990. [Google Scholar]
- Tiengtam, N.; Khempaka, S.; Paengkoum, P.; Booanuntanasarn, S. Effects of inulin and Jerusalem artichoke (Helianthus tuberosus) as prebiotic ingredients in the diet of juvenile Nile tilapia (Oreochromis niloticus). Anim. Feed Sci. Technol. 2016, 207, 120–129. [Google Scholar] [CrossRef]
- Pitaksong, T.; Kupittayanant, P.; Boonanuntanasarn, S. The effects of vitamins C and E on the growth, tissue accumulation and prophylactic response to thermal and acidic stress of hybrid catfish. Aquac Nutr 2013, 19, 148–162. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Anderson, D.P. Nonspecific defence mechanisms assay in fish. II. Potential killing activity of neutrophils and macrophages, lysozyme activity in serum and organs and total immunoglobulin (T- Ig) levels in serum. In Fish Diseases Diagnosis and Prevention’s Methods; FAO-Project GCP/INT/526/JPN; IFI: Olsztyn, Poland, 1993; pp. 105–112. [Google Scholar]
- Siwicki, A.K.; Studnicka, M. The phagocytic activity of neutrophils and serum lysozyme activity in experimentally infected carp, Cyprinus carpio L. J. Fish Biol. 1987, 31, 57–60. [Google Scholar] [CrossRef]
- Milla, S.; Mathieu, C.; Wang, N.; Lambert, S.; Nadzialek, S.; Massart, S.; Henrotte, E.; Douxfils, J.; Mélard, C.; Mandiki, S.N.M.; et al. Spleen immune status is affected after acute handling stress but not regulated by cortisol in Eurasian perch, Perca fluviatilis. Fish Shellfish Immunol. 2010, 28, 931–941. [Google Scholar] [CrossRef]
- Ricci, S.; Patoia, L.; Berrettini, M.; Binaglia, L.; Bucaneve, G.; Vecchini, A.; Carloni, I.; Carloni, I.; Agostini, L.; Parise, P. Fatty acid pattern of red blood cell membranes and risk of ischemic brain infarction: A case-control study. Stroke 1987, 18, 575–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougherty, R.M.; Galli, C.; Ferro-Luzzi, A.; Iacono, J.M. Lipid and phospholipid fatty acid composition of plasma, red blood cells, and platelets and how they are affected by dietary lipids: A study of normal subjects from Italy, Finland, and the USA. Am. J. Clin. Nutr. 1987, 45, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Revskij, D.; Haubold, S.; Viergutz, T.; Kröger-Koch, C.; Tuchscherer, A.; Kienberger, H.; Rychlik, M.; Tröscher, A.; Hammon, H.M.; Schuberth, H.J.; et al. Dietary Fatty Acids Affect Red Blood Cell Membrane Composition and Red Blood Cell ATP Release in Dairy Cows. Int. J. Mol. Sci. 2019, 20, 2769. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.W.; Costa, D.V.d.; Leal, C.A.G.; Figueiredo, H.C.P.; Rosa, P.V. Dietary oil sources on the innate immunity and resistance of Nile tilapia, Oreochromis niloticus, to Streptococcus agalactiae challenge. J. World Aquacult. Soc. 2015, 46, 252–262. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T. The innate and adaptive immune system of fish. In Infectious Disease in Aquaculture; Austin, B., Ed.; Woodhead Publishing: Cambridge, UK, 2012; pp. 3–68. [Google Scholar]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of Omega-3 Fatty Acids on Immune Cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef] [Green Version]
- Ayisi, C.L.; Zhao, J.; Wu, J.W. Replacement of fish oil with palm oil: Effects on growth performance, innate immune response, antioxidant capacity and disease resistance in Nile tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0196100. [Google Scholar] [CrossRef] [Green Version]
- Tan, P.; Dong, X.; Xu, H.; Mai, K.; Ai, Q. Dietary Vegetable Oil Suppressed Non-Specific Immunity and Liver Antioxidant Capacity but Induced Inflammatory Response in Japanese Sea Bass (Lateolabrax Japonicus). Fish Shellfish Immunol. 2017, 63, 139–146. [Google Scholar] [CrossRef]
- Borth, W. Alpha 2-macroglobulin, a multifunctional binding protein with targeting characteristics. FASEB J. 1992, 6, 3345–3353. [Google Scholar] [CrossRef]
- Rehman, A.A.; Ahsan, H.; Kahn, F.H. Alpha-2-Macroglobulin: A physiological guardian. J. Cell. Physiol. 2013, 228, 1665–1675. [Google Scholar] [CrossRef]
- Borth, W.; Luger, T.A. Identification of Alpha 2-macroglobulin as a Cytokine Binding Plasma Protein. Binding of interleukin-1 Beta to “F” Alpha 2-macroglobulin. J. Biol. Chem. 1989, 264, 5818–5825. [Google Scholar]
- do Nascimento, O.V.; Lima-Neto, A.B.M.; van Tilburg, M.F.; de Oliveira Monteiro-Moreira, A.C.; Lobo, M.D.P.; Rondina, D.; Fernandes, V.O.; Montenegro, A.P.D.R.; Montenegro, R.M. Proteomic analysis to identify candidate biomarkers associated with type 1 diabetes. Diabetes Metab Syndr Obes. 2018, 11, 289–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.; Yang, Z.; Cheng, Y.; Wang, J.; Zhou, J. Effects of the complete replacement of fish oil with linseed oil on growth, fatty acid composition, and protein expression in the Chinese mitten crab (Eriocheir sinensis). Proteome Sci. 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Wypasek, E.; Klukowska, A.; Zdziarska, J.; Zawilska, K.; Treliński, J.; Iwaniec, T.; Mital, A.; Pietrys, D.; Sydor, W.; Neerman-Arbez, M.; et al. Genetic and clinical characterization of congenital fibrinogen disorders in Polish patients: Identification of three novel fibrinogen gamma chain mutations. Thromb. Res. 2019, 182, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Lew, J.L.; Huang, L.; Yu, J.; Zhang, T.; Hrywna, Y.; Thompson, J.R.; de Pedro, N.; Blevins, R.A.; Pelaez, F.; et al. Human kininogen gene is transactivated by the farnesoid X receptor. J. Biol. Chem. 2003, 278, 28765–28770. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Wollaston-Hayden, E.E.; Markowski, T.W.; Higgins, L.A.; Mashek, D.G. Quantitative analysis of the murine lipid droplet-associated proteome during diet-induced hepatic steatosis. J. Lipid Res. 2015, 56, 2260–2272. [Google Scholar] [CrossRef] [Green Version]
- Blank, M.; Shoenfeld, Y. Histidine-rich glycoprotein modulation of immune/autoimmune, vascular, and coagulation systems. Clin. Rev. Allergy Immunol. 2008, 34, 307–331. [Google Scholar] [CrossRef]
- Priebatsch, K.M.; Poon, I.K.H.; Patel, K.K.; Kvansakul, M.; Hulett, M.D. Divalent metal binding by histidine-rich glycoprotein differentially regulates higher order oligomerisation and proteolytic processing. FEBS Lett. 2017, 591, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Hahl, P.; Hunt, R.; Bjes, E.S.; Skaff, A.; Keightley, A.; Smith, A. Identification of oxidative modifications of hemopexin and their predicted physiological relevance. J. Biol. Chem. 2017, 292, 13658–13671. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Sammy, F.; Yang, H.; Thundivalappil, S.; Hellman, J.; Tracey, K.J.; Warren, H.S. Identification of hemopexin as an anti-inflammatory factor that inhibits synergy of hemoglobin with HMGB1 in sterile and infectious inflammation. J. Immunol. 2012, 189, 2017–2022. [Google Scholar] [CrossRef] [Green Version]
- Montecinos, L.; Eskew, J.D.; Smith, A. What Is Next in This “Age” of Heme-Driven Pathology and Protection by Hemopexin? An Update and Links with Iron. Pharmaceuticals 2019, 12, 144. [Google Scholar] [CrossRef] [Green Version]
- Serebrenik, Y.V.; Hellerschmied, D.; Toure, M.; López-Giráldez, F.; Brookner, D.; Crews, C.M. Targeted protein unfolding uncovers a Golgi-specific transcriptional stress response. Mol. Biol. Cell 2018, 29, 1284–1298. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhou, L.; Zhang, H.; Chen, R.; Zhang, Y.; Li, L.; Lu, J.Y.; Jiang, H.; Liu, D.; Qi, S. Regulation of hepatic lipogenesis by the zinc finger protein Zbtb20. Nat. Commun. 2017, 8, 14824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, S.; Hess, M.A.; Ormonde-Hanson, P.; Malandro, J.; Hu, H.; Chen, M.; Kehrer, R.; Frodsham, M.; Schumacher, C.; Beluch, M.; et al. A broad role for the zincfinger protein ZNF202 in human lipid metabolism. J. Biol. Chem. 2000, 275, 15685–15690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertomeu, T.; Coulombe-Huntington, J.; Chatr-aryamontri, A.; Bourdages, K.G.; Coyaud, E.; Raught, B.; Xia, Y.; Tyersa, M. A High-Resolution Genome-Wide CRISPR/Cas9 Viability Screen Reveals Structural Features and Contextual Diversity of the Human Cell-Essential Proteome. Mol. Cell. Biol. 2018, 38, e00302-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Burdette, J.E.; Wang, J.P. Integrative network analysis of TCGA data for ovarian cancer. BMC Syst. Biol. 2014, 8, 1338. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Hayashi, T.; Fiches, G.; Xu, Q.; Li, M.; Que, J.; Liu, S.; Zhang, W.; Qi, J.; Santoso, N.; et al. Diversified Application of Barcoded PLATO (PLATO-BC) Platform for Identification of Protein Interactions. Genom. Proteomics Bioinform. 2019, 17, 319–331. [Google Scholar] [CrossRef]
- Maitre, J.L.; Heisenberg, C.P. Three Functions of Cadherins in Cell Adhesion Review. Curr. Biol. 2013, 23, R626–R633. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, D.B.; Philippova, M.P.; Tkachuk, V.A. Structure and Functions of Classical Cadherins. Biochemistry 2001, 66, 1174–1186. [Google Scholar] [CrossRef]
- Jiang, W.G.; Hiscox, S.; Horrobin, D.F.; Hallett, M.B.; Mansel, R.E.; Puntis, M.C. Expression of Catenins in Human Cancer Cells and Its Regulation by n-6 Polyunsaturated Fatty Acids. Anticancer Res. 1995, 15, 2569–2573. [Google Scholar]
- Zou, Z.; Bellenger, S.; Massey, K.A.; Nicolaou, A.; Geissler, A.; Bidu, C.; Bonnotte, B.; Pierre, A.S.; Minville-Walz, M.; Rialland, M.; et al. Inhibition of the HER2 pathway n-3 polyunsaturated fatty acids prevents breast cancer infat-1 transgenic mice. J. Lipid Res. 2013, 54, 3453–3463. [Google Scholar] [CrossRef] [Green Version]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA Abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, S.; Pal, R. Integrated analysis of transcriptomic and proteomic data. Curr. Genom. 2013, 14, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
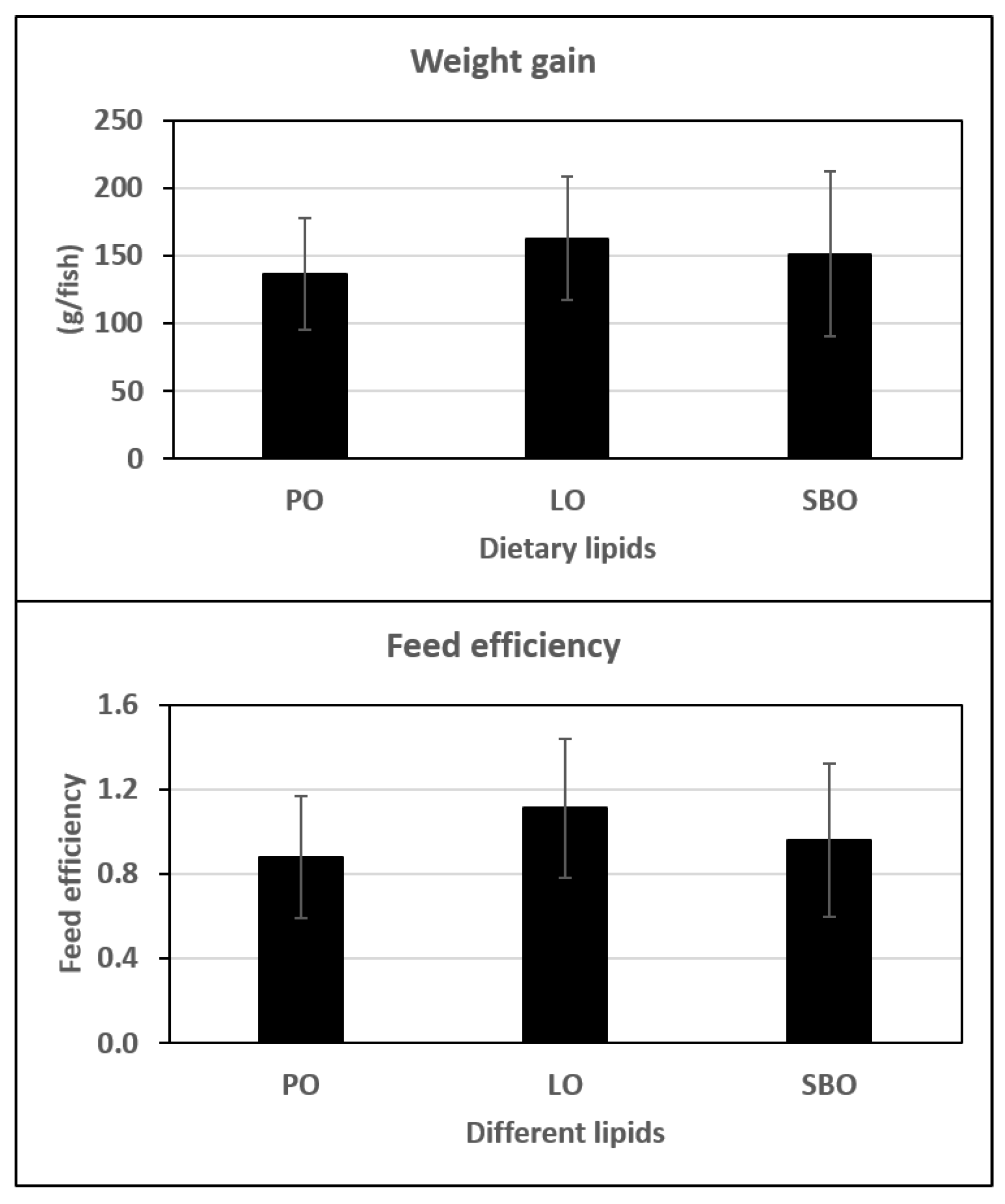



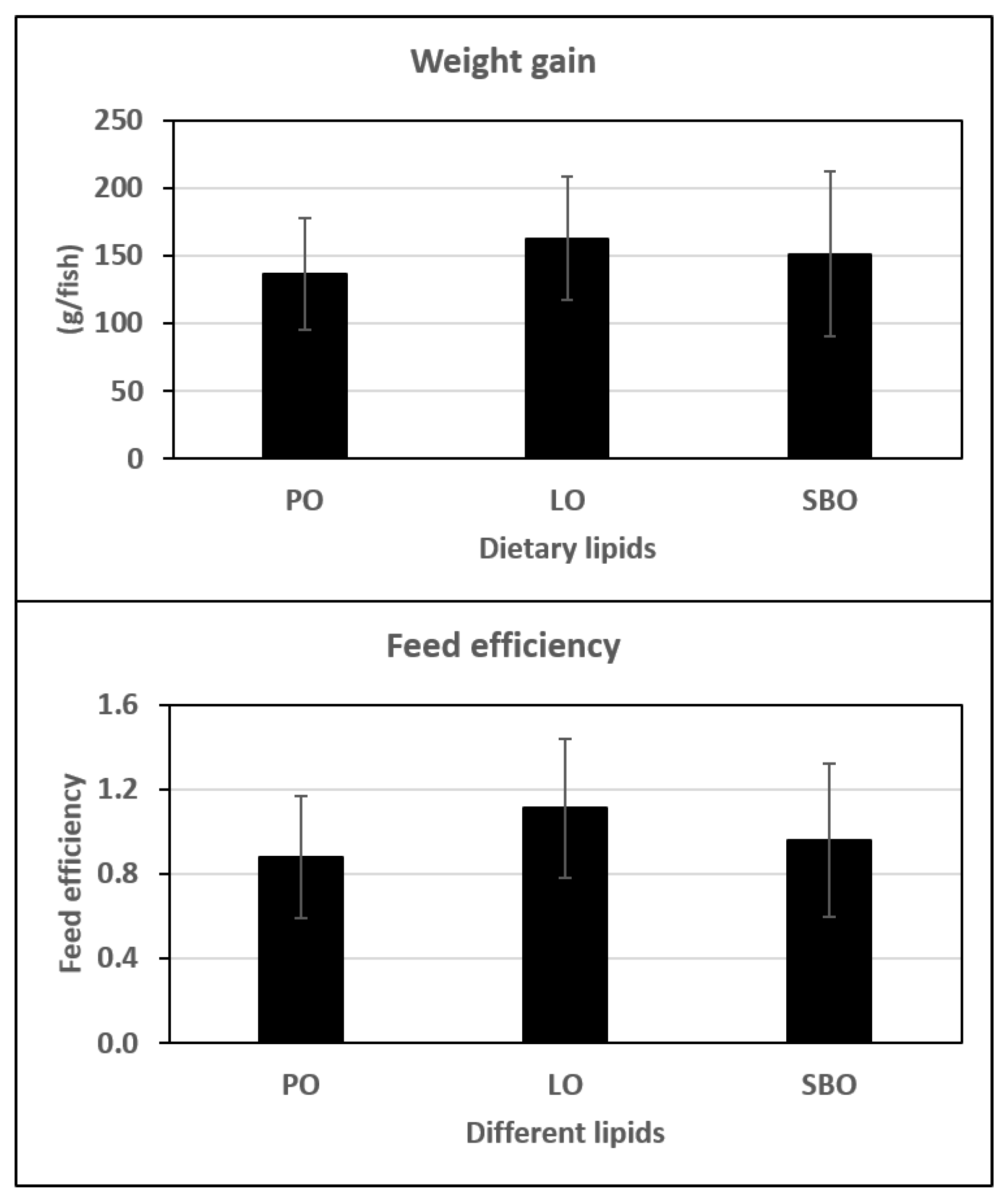{kind=link}
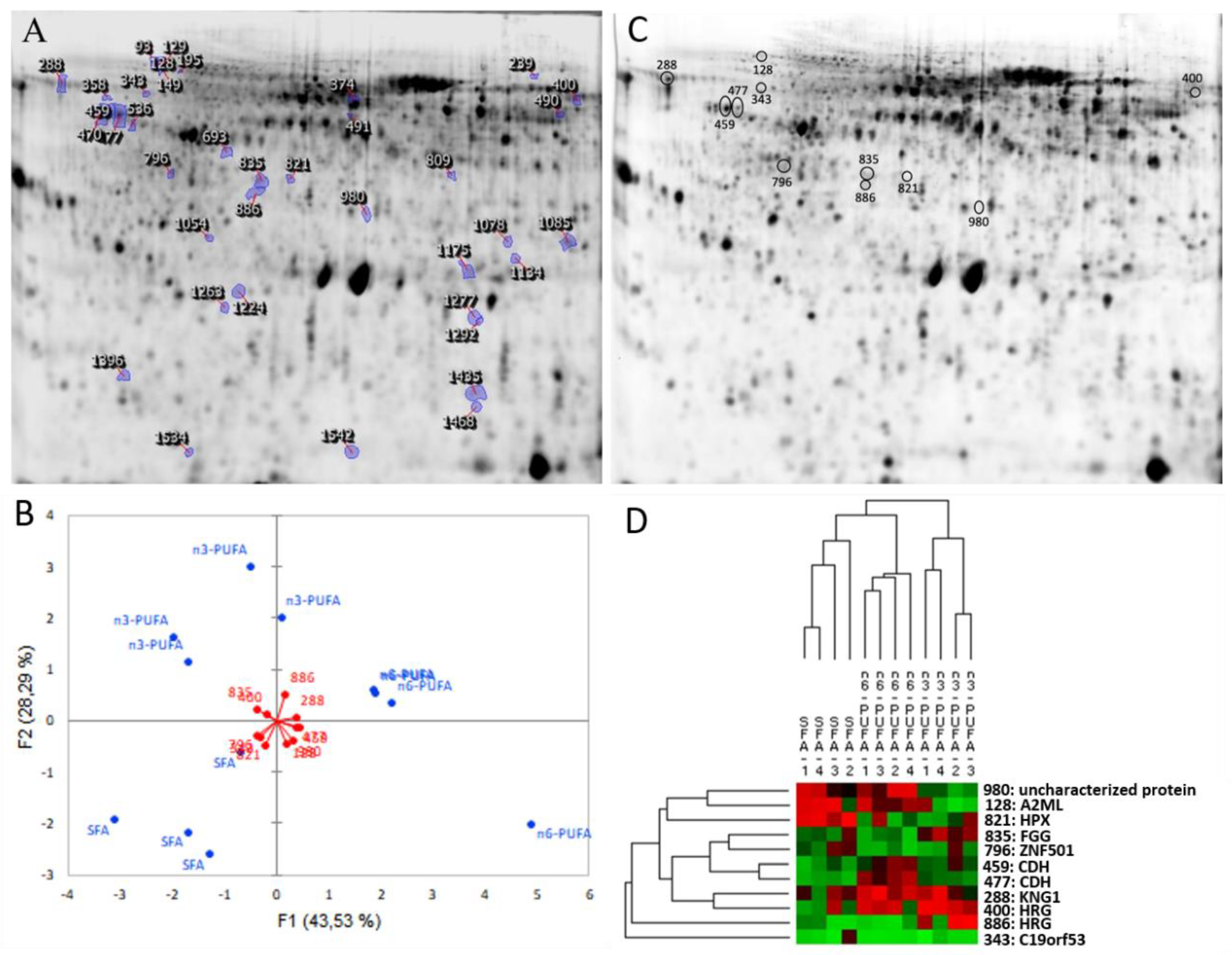{kind=link}
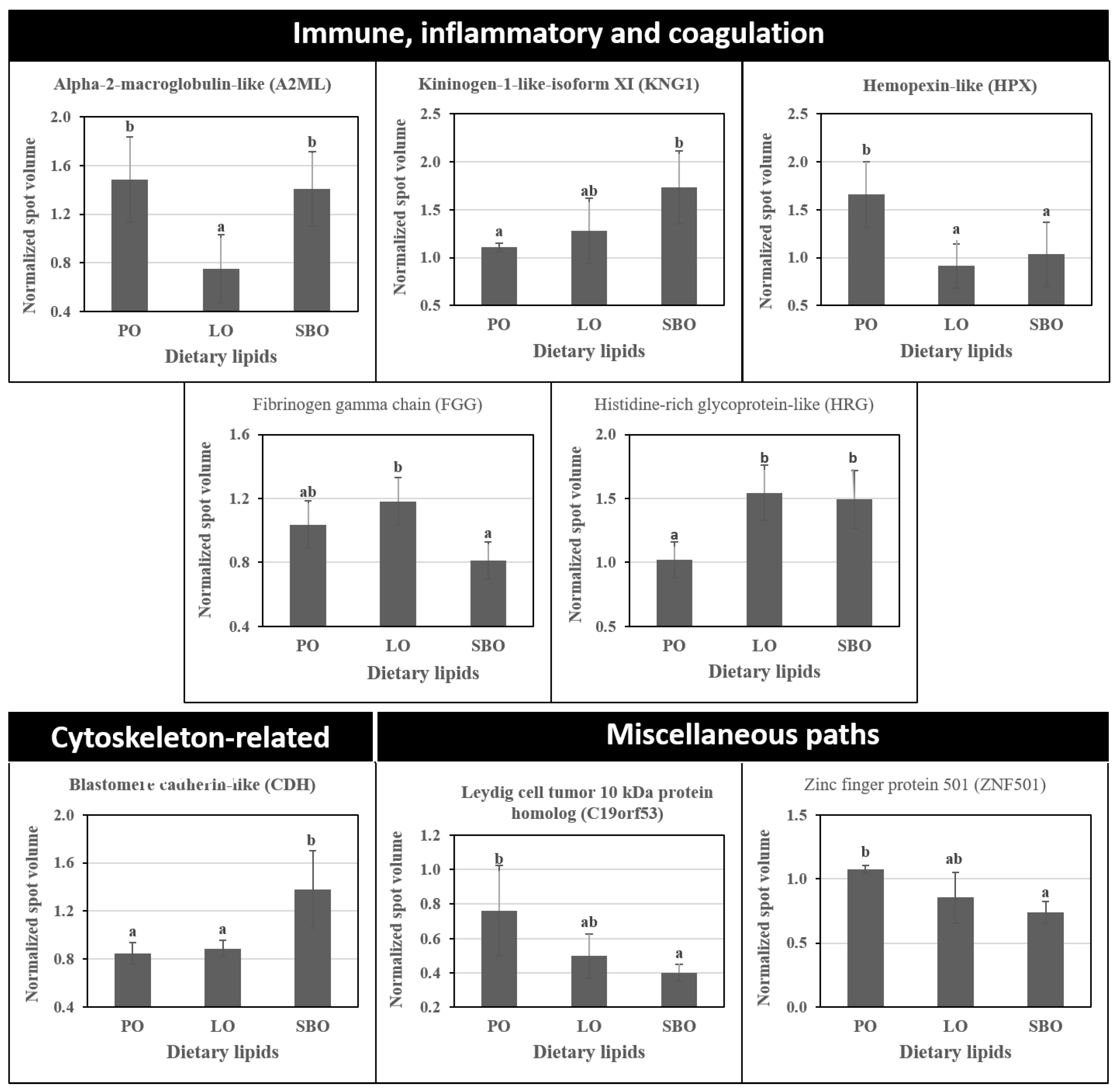{kind=link}
| Ingredient (%) | Diet | ||
|---|---|---|---|
| PO | LO | SBO | |
| Casein | 32.0 | 32.0 | 32.0 |
| Gelatin | 6.0 | 6.0 | 6.0 |
| Palm oil | 10.0 | - | - |
| Linseed oil | - | 10.0 | - |
| Soybean oil | - | - | 10.0 |
| Dextrin | 30.0 | 30.0 | 30.0 |
| Dicalcium phosphate | 1.0 | 1.0 | 1.0 |
| Premix 1 | 4.0 | 4.0 | 4.0 |
| Carboxymethyl cellulose | 2.0 | 2.0 | 2.0 |
| Alpha-cellulose | 15.0 | 15.0 | 15.0 |
| Proximate composition (%) | |||
| Dry matter | 81.3 | 81.5 | 81.8 |
| Crude protein | 28.3 | 28.3 | 28.3 |
| Crude lipid | 10.7 | 10.4 | 10.6 |
| Crude fibre | 2.7 | 2.8 | 2.8 |
| Crude Ash | 3.2 | 3.3 | 3.3 |
| Nitrogen-free extract 2 | 36.4 | 36.7 | 36.8 |
| Gene Transcript Name (Abbreviation) | GenBank Accession Number | 5′ to 3′ Nucleotide Sequences1 | Annealing Temperature (°C) | Expected Amplicon Size (bp) |
|---|---|---|---|---|
| CDH | XM_005465470.2 | 5′-AGGCCAAAGATGATGATGAGCCAG-3′ 5′-TCCCGCCATATCTGCTGCTTGTAC-3′ | 58 | 200 |
| FGG | XP_013129545.1 | 5′-GTTTCTACTTCGGTGGAGATGCAG-3′ 5′-GCAGCATGACATCTGTTCATCCAC-3′ | 55 | 109 |
| HPX | XP_003440941.1 | 5′-CCTCAAGGAGAGTGCATTACCGAC-3′ 5′-GAACCTGGTGAAGTTGTGTCCATG-3′ | 60 | 198 |
| A2ML | XP_005461528.1 | 5′-GGAAGAGGACCAGGAAACACTTGG-3′ 5′-ACACCACCCTTCATTCTGTTGTTG-3′ | 55 | 187 |
| KNG1 | XP_005464291.1 | 5′-TGCCACAGCTACATCTTCACCCTC-3′ 5′-GGCTTCAAAGCTTTCCAGGGCTTG-3′ | 55 | 162 |
| HRG | XP_005475829.1 | 5′-GATGGTGATGCCACCATAACCA-3′ 5′-CGTCCCACTTCCTTCAAGATGTAG-3′ | 58 | 202 |
| ZNF501 | XM_005477455.2 | 5′-CTGAAGAGACACATGGCACTTCAC-3′ 5′-CCTGTGTGAACTCTGATGTGCGTC-3′ | 58 | 198 |
| C19orf53 | XM_003455758.4 | 5′-CGAAGAAAGGAGGGAGGATCATTG-3′ 5′-TTAATCACACTCAGCGGCTTGTGG-3′ | 55 | 157 |
| Parameters | Diet | ||
|---|---|---|---|
| PO | LO | SBO | |
| Haematological parameters | |||
| RBC1 (cells × 106 µL−1) | 1.7 ± 0.2 | 2.3 ± 0.3 | 2.0 ± 0.4 |
| Haemoglobin (g dL−1) | 8.9 ± 0.5 | 9.7 ± 1.3 | 8.8 ± 0.6 |
| Haematocrit (%) | 29.1 ± 0.5 | 31.6 ± 3.9 | 29.1 ± 1.3 |
| Immune parameters | |||
| Lysozyme activity (µg mL−1) | 11.32 ± 1.16 | 12.34 ± 0.73 | 11.73 ± 0.60 |
| Total immunoglobulin (mg L−1) | 2.89 ± 0.31 | 2.57 ± 0.13 | 2.58 ± 0.34 |
| ACH502 (units mL−1) | 94.82 ± 11.19 a | 128.20 ± 17.37 b | 105.02 ± 26.92 ab |
| Spot No. | Protein [Species] | Entry Name (Uniprot) | Mascot Score | GI Number a | Mwt/pIt b (Mwe/pIe c) | Sequence Coverage (%) | Number of Matched Peptides | Best Peptide Match: Sequence |
|---|---|---|---|---|---|---|---|---|
| Immune, inflammatory and coagulation | ||||||||
| 128 | Alpha-2-macroglobulin-like [O. niloticus] | A2ML | 78 | gi|542250738 | 74,969/5.64 (73,451/4.8) | 18 | 5 | MSIFTKMEQK |
| 288 | Kininogen-1-like isoform X1 [O. niloticus] | KNG1 | 122 | gi|542257849 | 41,185/5.80 (66,931/4.4) | 44 | 5 | AVHSAVDKFNER |
| 821 | Hemopexin-like [O. niloticus] | HPX | 84 | gi|348506792 | 50,278/5.91 (37,438/5.3) | 4 | 2 | IRDVDLSATPR |
| 835 | Fibrinogen gamma chain [O. niloticus] | FGG | 83 | gi|908456888 | 42,405/4.80 (38,318/5.2) | 35 | 3 | SSDPDVHFK |
| 886 400 | Histidine-rich glycoprotein-Like [O. niloticus] | HRG d | 117 94 | gi|542204170 [gi|908518607] | 48,193/6.64 (36,015/5.2) [39,519/6.52] [(62,423/6.2)] | 33 31 | 6 4 | FRLHEVQGNSVEQVDGGCNVK K.AVHHINEYHDHGYK.F |
| Cytoskeleton-related proteins | ||||||||
| 477 459 | Blastomere cadherin-like [O. niloticus] | CDH d | 120 102 | gi|542261060 | 64,731/4.67 (55147/4.6) | 35 10 | 7 3 | KLFYSISGPGADLPPVDR [R.VFPDVFFTENNR.G] |
| Miscellaneous paths | ||||||||
| 343 | Leydig cell tumour 10 kDa protein homolog [Ictalurus punctatus] | C19orf53 | 343 | gi|1042336086 | 10,905/11.63 (63,891/4.7) | 30 | 4 | KQLNKPK |
| 796 | Zinc finger protein 501 [Sinocyclocheilus rhinocerous] | ZNF501 | 54 | gi|1025318457 | 44,576/8.99 (40,768/4.9) | 12 | 4 | VKTEFIK |
| 980 | uncharacterized protein KIAA2012-like isoform X1 [Lepisosteus oculatus] | 51 | gi|973180948 | 91,103/5.93 (33,331/5.5) | 7 | 6 | R.TLRDLTGAILAYGNK.Q | |
| Target Genes | Diet | ||
|---|---|---|---|
| PO | LO | SBO | |
| Liver | |||
| a2ml | 0.76 ± 0.05 | 0.73 ± 0.04 | 0.69 ± 0.04 |
| kng1 | 0.72 ± 0.06 b | 0.64 ± 0.03 a | 0.60 ± 0.06 a |
| hpx | 0.59 ± 0.08 | 0.50 ± 0.01 | 0.53 ± 0.07 |
| fgg | 0.73 ± 0.04 | 0.71 ± 0.03 | 0.70 ± 0.02 |
| hrg | 0.22 ± 0.03 b | 0.16 ± 0.04 ab | 0.12 ± 0.06 a |
| cdh | 0.33 ± 0.02 b | 0.30 ± 0.00 a | 0.30 ± 0.01 a |
| c19orf53 1 | 0.41 ± 0.04 | 0.38 ± 0.02 | 0.37 ± 0.01 |
| znf501 1 | 0.60 ± 0.02 b | 0.56 ± 0.02 a | 0.56 ± 0.00 a |
| Muscle | |||
| a2ml | 0.48 ± 0.14 | 0.46 ± 0.14 | 0.53 ± 0.05 |
| kng1 | 0.42 ± 0.03 a | 0.49 ± 0.07 b | 0.51 ± 0.04 b |
| hpx | 0.20 ± 0.12 a | 0.27 ± 0.11 ab | 0.37 ± 0.04 b |
| fgg | 0.35 ± 0.03 | 0.41 ± 0.10 | 0.44 ± 0.04 |
| cdh | 0.23 ± 0.14 | 0.24 ± 0.08 | 0.28 ± 0.03 |
| c19orf53 | 0.57 ± 0.03 | 0.58 ± 0.06 | 0.53 ± 0.02 |
| znf501 | 0.38 ± 0.05 a | 0.47 ± 0.09 ab | 0.49 ± 0.04 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakharuthai, C.; Rodrigues, P.M.; Schrama, D.; Kumkhong, S.; Boonanuntanasarn, S. Effects of Different Dietary Vegetable Lipid Sources on Health Status in Nile Tilapia (Oreochromis niloticus): Haematological Indices, Immune Response Parameters and Plasma Proteome. Animals 2020, 10, 1377. https://doi.org/10.3390/ani10081377
Nakharuthai C, Rodrigues PM, Schrama D, Kumkhong S, Boonanuntanasarn S. Effects of Different Dietary Vegetable Lipid Sources on Health Status in Nile Tilapia (Oreochromis niloticus): Haematological Indices, Immune Response Parameters and Plasma Proteome. Animals. 2020; 10(8):1377. https://doi.org/10.3390/ani10081377
Chicago/Turabian StyleNakharuthai, Chatsirin, Pedro M. Rodrigues, Denise Schrama, Suksan Kumkhong, and Surintorn Boonanuntanasarn. 2020. "Effects of Different Dietary Vegetable Lipid Sources on Health Status in Nile Tilapia (Oreochromis niloticus): Haematological Indices, Immune Response Parameters and Plasma Proteome" Animals 10, no. 8: 1377. https://doi.org/10.3390/ani10081377
APA StyleNakharuthai, C., Rodrigues, P. M., Schrama, D., Kumkhong, S., & Boonanuntanasarn, S. (2020). Effects of Different Dietary Vegetable Lipid Sources on Health Status in Nile Tilapia (Oreochromis niloticus): Haematological Indices, Immune Response Parameters and Plasma Proteome. Animals, 10(8), 1377. https://doi.org/10.3390/ani10081377