A Deletion Downstream of the CHCHD7 Gene Is Associated with Growth Traits in Sheep

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Samples and Data Collection
2.3. Isolation and Use of Sheep Genomic DNA
2.4. Primer Design, PCR Amplification, and Genotyping
2.5. Linkage Disequilibrium Analysis with Two Deletions in the PLAG1 Gene
2.6. Statistical Analyses
3. Results
3.1. Genotyping of 8-bp Deletion Downstream of the CHCHD7 Gene
3.2. Genetic Parameter Analysis of the 8-bp Deletion Downstream of the Sheep CHCHD7 Gene
3.3. Analysis of the Association between the 8-bp Deletion and Growth Traits in Tan Sheep
3.4. Linkage Disequilibrium Analysis between the 8-bp Deletion Downstream of the CHCHD7 Gene and Two Deletions in the PLAG1 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, J.; Erdenee, S.; Zhang, S.; Wei, Z.; Zhang, M.; Jin, Y.; Wu, H.; Chen, H.; Sun, X.; Xu, H.; et al. Genetic effects of PRNP gene insertion/deletion (indel) on phenotypic traits in sheep. Prion 2018, 12, 42–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liu, S.; Yuan, T.; Sun, X. Genetic effects of FTO gene insertion/deletion (InDel) on fat-tail measurements and growth traits in Tong sheep. Anim. Biotechnol. 2019, 23, 1–11. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, S.; Li, J.; Wang, X.; Peng, K.; Lan, X.; Pan, C. Development of a touch-down multiplex PCR method for simultaneously rapidly detecting three novel insertion/deletions (indels) within one gene: An example for goat GHR gene. Anim. Biotechnol. 2019, 30, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Jiang, E.; Yan, H.; Zhu, H.; Chen, H.; Liu, J.; Qu, L.; Pan, C.; Lan, X. InDels within caprine IGF2BP1 intron 2 and the 3’-untranslated regions are associated with goat growth traits. Anim. Genet. 2020, 51, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, X.; Ma, L.; Xu, H.; Cao, X.; Luo, R.; Chen, H.; Sun, X.; Cai, Y.; Lan, X. Detection of a new 20-bp insertion/deletion (indel) within sheep PRND gene using mathematical expectation (ME) method. Prion 2017, 11, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Erdenee, S.; Li, J.; Kang, Z.; Xu, H.; Zang, R.; Cao, X.; Yang, J.; Cai, Y.; Lan, X. Sheep zinc finger proteins 395 (ZNF395): Insertion/deletion variations, associations with growth traits, and mRNA expression. Anim. Biotechnol. 2019, 31, 237–244. [Google Scholar] [CrossRef]
- Nishimura, S.; Watanabe, T.; Mizoshita, K.; Tatsuda, K.; Fujita, T.; Watanabe, N.; Sugimoto, Y.; Takasuga, A. Genome-wide association study identified three major QTL for carcass weight including the PLAG1-CHCHD7 QTN for stature in Japanese Black cattle. BMC Genet. 2012, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, I.A.; Khatkar, M.S.; Thomson, P.C.; Raadsma, H.W. Composite selection signals can localize the trait specific genomic regions in multi-breed populations of cattle and sheep. BMC Genet. 2014, 15, 34. [Google Scholar] [CrossRef] [Green Version]
- Lettre, G.; Jackson, A.U.; Gieger, C.; Schumacher, F.R.; Berndt, S.I.; Sanna, S.; Eyheramendy, S.; Voight, B.F.; Butler, J.L.; Guiducci, C.; et al. Identification of ten loci associated with height highlights new biological pathways in human growth. Nat. Geneti. 2008, 40, 584–591. [Google Scholar] [CrossRef]
- Taye, M.; Yoon, J.; Dessie, T.; Cho, S.; Oh, S.J.; Lee, H.K.; Kim, H. Deciphering signature of selection affecting beef quality traits in Angus cattle. Genes Genom. 2018, 40, 63–75. [Google Scholar] [CrossRef]
- Jiao, S.; Maltecca, C.; Gray, K.A.; Cassady, J.P. Feed intake, average daily gain, feed efficiency, and real-time ultrasound traits in Duroc pigs: II. Genome-wide association. J. Anim. Sci. 2014, 92, 2846–2860. [Google Scholar] [CrossRef] [PubMed]
- Asp, J.; Persson, F.; Kost-Alimova, M.; Stenman, G. CHCHD7-PLAG1 and TCEA1-PLAG1 gene fusions resulting from cryptic, intrachromosomal 8q rearrangements in pleomorphic salivary gland adenomas. Genes Chromosom. Cancer 2006, 45, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Fink, T.; Tiplady, K.; Lopdell, T.; Johnson, T.; Snell, R.G.; Spelman, R.J.; Davis, S.R.; Littlejohn, M.D. Functional confirmation of PLAG1 as the candidate causative gene underlying major pleiotropic effects on body weight and milk characteristics. Sci. Rep. 2017, 7, 44793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signer-Hasler, H.; Burren, A.; Ammann, P.; Drögemüller, C.; Flury, C. Runs of homozygosity and signatures of selection: A comparison among eight local Swiss sheep breeds. Anim. Genet. 2019, 50, 512–525. [Google Scholar] [CrossRef]
- Wu, H.; Pan, Y.; Zhang, Q.; Cao, Y.; Li, J.; Chen, H.; Cai, Y.; Sun, X.; Lan, X. Insertion/deletion (InDel) variations in sheep PLAG1 gene locating in growth-related major QTL are associated with adult body weight and morphometric traits. Small Rumin. Res. 2019, 178, 63–69. [Google Scholar] [CrossRef]
- Utsunomiya, Y.T.; do Carmo, A.S.; Carvalheiro, R.; Neves, H.H.; Matos, M.C.; Zavarez, L.B.; Pérez O’Brien, A.M.; Sölkner, J.; McEwan, J.C.; Cole, J.B.; et al. Genome-wide association study for birth weight in Nellore cattle points to previously described orthologous genes affecting human and bovine height. BMC Genet. 2013, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Liu, N.; Zhang, X.; Zhang, Y.; Qu, L.; Yan, H.; Lan, X.; Dong, W.; Pan, C. A 20-bp insertion/deletion (indel) polymorphism within the CDC25A gene and its associations with growth traits in goat. Arch. Anim. Breed. 2019, 62, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.D.; He, S.; Zhu, Y.J.; Cao, X.; Luo, R.Y.; Cai, Y.; Xu, H.W.; Sun, X.Z. A novel 29 bp insertion/deletion (indel) variant of the LHX3 gene and its influence on growth traits in four sheep breeds of various fecundity. Arch. Anim. Breed. 2017, 60, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Müllenbach, R.; Lagoda, P.J.; Welter, C. An efficient salt-chloroform extraction of DNA from blood and tissues. Trends Genet. 1989, 5, 391. [Google Scholar] [PubMed]
- Li, Z.Q.; Zhang, Z.; He, Z.D.; Tang, W.; Li, T.; Zeng, Z.; He, L.; Shi, Y.Y. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: Update of the SHEsis. Cell Res. 2009, 19, 519–523. [Google Scholar] [CrossRef]
- Shi, T.; Peng, W.; Yan, J.; Cai, H.; Lan, X.; Lei, C.; Bai, Y.; Chen, H. A novel 17 bp indel in the SMAD3 gene alters transcription level, contributing to phenotypic traits in Chinese cattle. Arch. Anim. Breed. 2016, 59, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Shi, J.; Luo, Y.Q.; Sun, S.Y.; Pu, M. Genetic characterization of the gypsy moth from China (Lepidoptera, Lymantriidae) using inter simple sequence repeats markers. PLoS ONE 2013, 8, e73017. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Bi, Y.; Wang, R.; Pan, C.; Chen, H.; Lan, X.; Qu, L. Detection of a 4 bp Mutation in the 3’UTR Region of Goat Sox9 Gene and Its Effect on the Growth Traits. Animals 2020, 10, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khani, K.; Abdolmohammadi, A.; Foroutanifar, S.; Zebarjadi, A. Assessment of polymorphisms in mysostatin gene and their allele substitution effects showed weak association with growth traits in Iranian Markhoz goats. J. Agric. Sci. 2017, 155, 519–526. [Google Scholar] [CrossRef]
- Ren, F.; Yu, S.; Chen, R.; Lv, X.; Pan, C. Identification of a novel 12-bp insertion/deletion (indel) of iPS-related Oct4 gene and its association with reproductive traits in male piglets. Anim. Reprod Sci. 2017, 178, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, C.M.; Chan, K.; Lau, Y.F. A PCR artifact: Generation of heteroduplexes. Am. J. Hum. Genet. 1989, 45, 337–339. [Google Scholar] [PubMed]
- Song, Y.; Xu, L.; Chen, Y.; Zhang, L.; Gao, H.; Zhu, B.; Niu, H.; Zhang, W.; Xia, J.; Gao, X.; et al. Genome-Wide Association Study Reveals the PLAG1 Gene for Knuckle, Biceps and Shank Weight in Simmental Beef Cattle. PLoS ONE 2016, 11, e0168316. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, I.A.; Khatkar, M.S.; Thomson, P.C.; Raadsma, H.W. Composite selection signals for complex traits exemplified through bovine stature using multibreed cohorts of European and African Bos taurus. G3 Genes Genomes Genet. 2015, 5, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Cheruiyot, E.K.; Bett, R.C.; Amimo, J.O.; Zhang, Y.; Mrode, R.; Mujibi, F.D.N. Signatures of selection in admixed dairy cattle in Tanzania. Front Genet. 2018, 9, 607. [Google Scholar] [CrossRef] [Green Version]
- Van Laere, A.S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Abi Habib, W.; Brioude, F.; Edouard, T.; Bennett, J.T.; Lienhardt-Roussie, A.; Tixier, F.; Salem, J.; Yuen, T.; Azzi, S.; Le Bouc, Y.; et al. Genetic disruption of the oncogenic HMGA2-PLAG1-IGF2 pathway causes fetal growth restriction. Genet. Med. 2018, 20, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visscher, P.M.; Goddard, M.E. Cattle gain stature. Nat. Genet. 2011, 43, 397–398. [Google Scholar] [CrossRef] [PubMed]
- Hiroshi, H.; Kouji, S.; Toshio, W.; Akihiro, K.; Kazunori, M.; Yoshikazu, S.; Akiko, T. Comparison of the effects explained by variations in the bovine PLAG1 and NCAPG genes on daily body weight gain, linear skeletal measurements and carcass traits in Japanese Black steers from a progeny testing program. Anim. Sci. J. 2013, 84, 529–534. [Google Scholar] [CrossRef]
- Latifa, K.; Haruko, T.; Li, L.; Tom, D.; Juan, A.; Denis, B.; Nadine, C.; Stephen, R.; Frédéric, F.; Bernard, G.; et al. Variants modulating the expression of a chromosome domain encompassing PLAG1 influence bovine stature. Nat. Genet. 2011, 43, 405–413. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; Guo, Y.; She, S.; Wang, B.; Jiang, Y.; Bai, Y.; Song, X.; Li, L.; Shi, L.; et al. Screening of deletion variants within the goat PRDM6 gene and its effects on growth traits. Animals 2020, 10, 208. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Li, M.; Cheng, H.; Fan, W.; Yuan, Z.; Gao, Q.; Xu, Y.; Guo, Z.; Zhang, Y.; Hu, J.; et al. An intercross population study reveals genes associated with body size and plumage color in ducks. Nat. Commun. 2018, 9, 2648. [Google Scholar] [CrossRef] [Green Version]

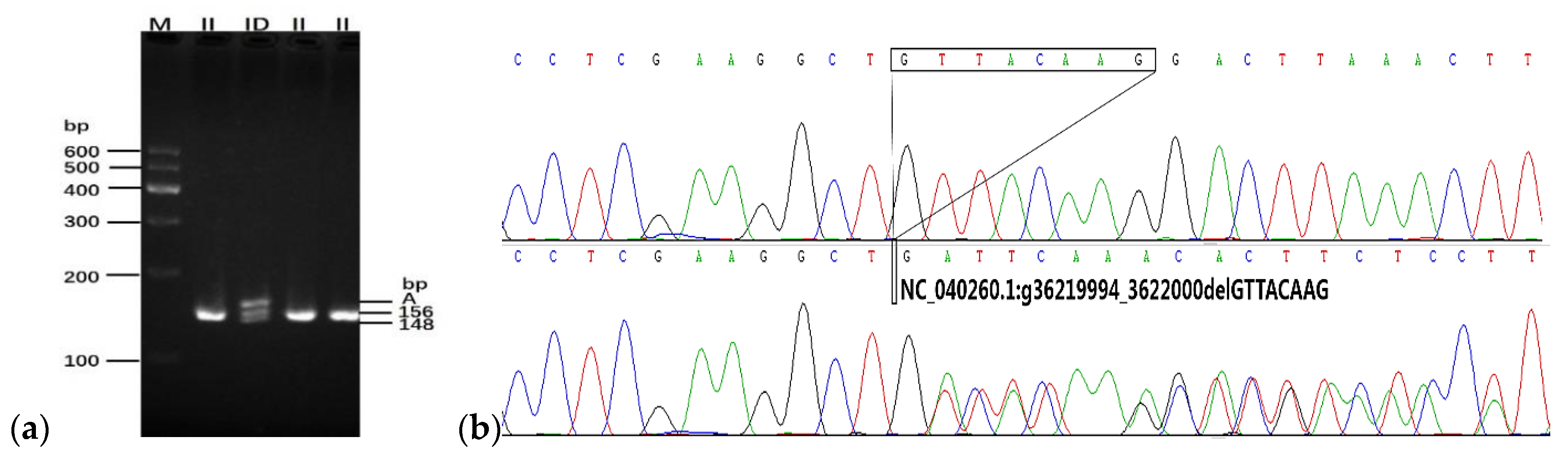


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeds | Sizes | Genotypic Frequencies | Allelic Frequencies | Population Parameters | |||||
|---|---|---|---|---|---|---|---|---|---|
| Wildtype | Deletion | Wildtype | Deletion | Ho | He | Ne | MAF | ||
| TS | 907 | 0.986 (n = 894) | 0.014 (n = 13) | 0.993 | 0.007 | 0.986 | 0.014 | 1.014 | 0.007 |
| LXBH | 629 | 0.978 (n = 615) | 0.022 (n = 14) | 0.989 | 0.011 | 0.978 | 0.022 | 1.023 | 0.011 |
| STHS | 190 | 0.984 (n = 187) | 0.016 (n = 3) | 0.992 | 0.008 | 0.984 | 0.016 | 1.016 | 0.008 |
| LFTS | 49 | 0.980 (n = 48) | 0.020 (n = 1) | 0.990 | 0.010 | 0.980 | 0.020 | 1.021 | 0.010 |
| HS | 201 | 1.000 (n = 201) | 0 (n = 0) | 1.000 | 0 | 1.000 | 0 | 1.000 | 0 |
| SS | 48 | 1.000 (n = 48) | 0 (n = 0) | 1.000 | 0 | 1.000 | 0 | 1.000 | 0 |
| AUW | 326 | 1.000 (n = 326) | 0 (n = 0) | 1.000 | 0 | 1.000 | 0 | 1.000 | 0 |
| Gender | Traits | Genotypes (LSM ± SE) | p-Values | |
|---|---|---|---|---|
| Wildtype | Deletion Type | |||
| Rams | BW (kg) | 33.71 ± 0.11 (n = 449) | 33.03 ± 0.58 (n = 9) | 0.256 |
| BH (cm) | 65.04 ± 0.15 (n = 449) | 62.67 ± 1.86 (n = 9) | 0.237 | |
| BL (cm) | 64.85 a ± 0.21 (n = 449) | 61.22 b ± 1.70 (n = 9) | 0.032 | |
| HHC (cm) | 62.52 ± 0.14 (n = 449) | 60.56 ± 1.89 (n = 9) | 0.287 | |
| PG (cm) | 78.78 ± 0.19 (n = 449) | 78.33 ± 1.00 (n = 9) | 0.717 | |
| CD (cm) | 26.54 a ± 0.11 (n = 449) | 24.66 b ± 0.68 (n = 9) | 0.015 | |
| CW (cm) | 20.74 ± 0.09 (n = 449) | 20.56 ± 0.60 (n = 9) | 0.649 | |
| CC (cm) | 8.14 ± 0.03 (n = 449) | 8.33 ± 0.14 (n = 9) | 0.171 | |
| Ewes | BW (kg) | 33.33 ± 0.10 (n = 445) | 33.48 ± 0.42 (n = 4) | 0.900 |
| BH (cm) | 63.74 ± 0.15 (n = 445) | 61.75 ± 1.11 (n = 4) | 0.164 | |
| BL (cm) | 65.12 ± 0.22 (n = 445) | 64.00 ± 1.35 (n = 4) | 0.493 | |
| HHC (cm) | 61.94 ± 0.14 (n = 445) | 61.00 ± 0.91 (n = 4) | 0.525 | |
| PG (cm) | 79.83 ± 0.18 (n = 445) | 79.25 ± 2.10 (n = 4) | 0.649 | |
| CD (cm) | 26.42 ± 0.10 (n = 445) | 25.50 ± 0.65 (n = 4) | 0.316 | |
| CW (cm) | 21.01 b ± 0.09 (n = 445) | 22.75 a ± 0.48 (n = 4) | 0.047 | |
| CC (cm) | 8.03 ± 0.03 (n = 445) | 8.00 ± 0.46 (n = 4) | 0.937 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Li, H.; Wang, Z.; Abudureyimu, A.; Yang, J.; Cao, X.; Lan, X.; Zang, R.; Cai, Y. A Deletion Downstream of the CHCHD7 Gene Is Associated with Growth Traits in Sheep. Animals 2020, 10, 1472. https://doi.org/10.3390/ani10091472
Xu H, Li H, Wang Z, Abudureyimu A, Yang J, Cao X, Lan X, Zang R, Cai Y. A Deletion Downstream of the CHCHD7 Gene Is Associated with Growth Traits in Sheep. Animals. 2020; 10(9):1472. https://doi.org/10.3390/ani10091472
Chicago/Turabian StyleXu, Hongwei, Haixia Li, Zhen Wang, Ayimuguli Abudureyimu, Jutian Yang, Xin Cao, Xianyong Lan, Rongxin Zang, and Yong Cai. 2020. "A Deletion Downstream of the CHCHD7 Gene Is Associated with Growth Traits in Sheep" Animals 10, no. 9: 1472. https://doi.org/10.3390/ani10091472