1. Introduction
Sardinia, located to the west of Italy in the Mediterranean Sea, is an island characterized by many autochthonous animal species. Among them, there are three horse breeds native to this island that are classified “at risk” by FAO (
http://www.fao.org/dad-is) if considering their local status.
The Giara horse, with its population size of 524 individuals (298 females and 226 males) (FAO 2018; last update: August 2019), is one of the 15 indigenous horse “breeds of limited distribution” in Italy recognized by the AIA, the Italian breeders’ association. The origin of these animals is shrouded in mystery. Even if it is considered an ancient native breed, some scholars claim that they were introduced from the Middle East. In 1540, Sigismondo Arquer was the first to state that there were these wild horses in Sardinia [
1]. In the XVIII century, numerous wild herds of Giara were observed in Sardinia and described in several reports. It takes the name from the Giara plateau, an area located at the Sardinian hinterland of about 45 km
2 between 500 and 600 m above sea level. A mountain chain surrounds the plateau limiting any connections with the underlying valleys throughout time, thus preventing the migration of horses and protecting the biodiversity of this population. The breed is well adapted to dry and harsh environments where it lives in the wild.
The Sarcidano Horse is a rare semi-feral breed originated from the plateau of Sarcidano in the Laconi municipality (province of Oristano). It is also recognized by the AIA (
http://www.aia.it) as “of limited distribution”, with 115 animals (65 females and 50 males) (FAO 2018; last update: March 2020). With the Giara breed, their local status is critical. They are both bred in semi-feral conditions and recorded in the Anagraphic Register (
http://www.anagrafeequidi.it).
The Sardinian Anglo-Arab breed is the result of selected crossbreeding between indigenous Sardinian mares carrying Arabian blood and Thoroughbred and Arab stallions [
2,
3]. It is considered an endangered horse breed currently counting 3005 animals (2871 females and 134 males) (FAO 2018; last update: September 2019). The breed is maintained under controlled breeding conditions and its genealogies have been registered in a dedicated Studbook since 1967. The Sardinian Anglo-Arab is widely used in different equestrian competitions such as flat races and jumping [
2,
4]; for this reason, it was frequently crossbred with Thoroughbreds.
The horse is the typical domestic mammalian species that was selectively bred for specific traits. Few stallions, producing most desirable offspring, were often crossed with local autochthonous mares resulting in a high variation among the maternal lines but a limited variability in the paternal counterpart, both at macro- and micro-geographic levels. This breeding management is reflected in the uniparental markers acquired from female (mitochondrial DNA) and male (Y chromosome) ancestors [
5,
6,
7,
8,
9,
10,
11,
12]. The striking low diversity displayed by the non-recombining region of the Y chromosome (NRY) reflects the small number of stallions that contributed to the establishment of current breeds [
6,
10]. Recently, Wallner and colleagues described the nucleotide variation of modern horse Y chromosomes, resulting in six haplotypes distinct from the Przewalski horse, and traced their distribution using pedigree information [
6,
13].
On the other hand, the female counterpart of modern horses usually manifests a high genetic variability. In particular, the mitochondrial DNA (mtDNA) control regions of Sardinian horse breeds were already analyzed together with other Italian breeds, highlighting a genetic structure for these breeds, who live in geographically isolated contexts [
7,
14]. Previous studies based on shorter mtDNA control-region sequences claimed that Giara and Sarcidano consist of two distinct gene pools without gene flows between them [
15]. The Giara horse was also described as genetically homogeneous and grouping in a unique clade with three ancient breeds (Sorraia, Garrano, and Potoka) [
16].
The aim of this study was to investigate and reconstruct the paternal and maternal phylogenetic histories of three Sardinian horse breeds (Giara, Sarcidano, and Sardinian Anglo-Arab) (
Table 1) by screening for the first time three loci of the NRY (YE3, YE17, and YXX) and by performing a parallel phylogenetic analysis on the mtDNA. This comparative study will shed light on their genetic peculiarities and evaluate the distribution of uniparental markers in present-day Sardinia, thus providing new knowledge for the enhancement of autochthonous genetic resources.
2. Materials and Methods
The analyses were conducted on a dataset of Sardinian horse DNAs previously collected and reported also in Cardinali et al., 2016 [
7] and Giontella et al., 2020 [
14]. All experimental procedures were reviewed and approved by the Animal Research Ethics Committee of the University of Perugia (Prot. N.2017_01).
For the Sardinian Anglo-Arab, genealogical information recorded in the Studbook was considered in order to select maternally unrelated samples belonging to dam lines with at least 100 descendants and deriving from all important and widespread sire lines. For the Giara and Sarcidano breeds, living in semi-feral conditions, sampling was performed during the late summer when the drought forces herds to reach the few available springs located within confined areas. These are equipped with corridors or small capture paddocks for the identification of subjects through a transponder, or any health treatments and biological sampling. In these peculiar circumstances, individuals were randomly sampled taking care to select specimens belonging to different groups in order to reduce the probability of sampling the genealogically closest subjects. Groups mainly consist of: solitary subjects (without herd); herds with a dominant stallion, about 8–10 mares and young foals; herds of only sexually mature young males.
Our complete dataset resulted in a total of 125 DNA samples (39 Giara, 30 Sarcidano, and 56 Sardinian Anglo-Arab) including 34 stallions.
2.1. Y-Chromosome Analysis
Three polymorphic sites of the NRY (YE3, YE17, and YXX) were analyzed for the 34 stallions belonging to the Sardinian breeds (12 Giara, 10 Sarcidano, and 12 Sardinian Anglo-Arab) as reported in Wallner and colleagues [
6] (
Table S1).
Referring to Wallner et al., 2013 [
6], we selected three pairs of oligonucleotides to amplify the polymorphic sites. For the locus YE3, we selected the forward primer (5′-CCCTCTGCTGAGCATCTAGG-3′) used to detect the mutation 10594delT and the reverse primer (5′-GGCTTAGGCCACTGATGGTA-3′) used to analyze the 20 single nucleotide polymorphisms (SNPs) and the indels. For the locus YE17, we amplified through the forward primer 5′-GGCCTAAGTTGTTCGCAGAG-3′ and the reverse 5′-TGACTGGTGGTGTCCAGTGT-3′, while to detect the SNP (Single Nucleotide Polymorphism) G/A in the locus YXX, we amplified by using the forward primer 5′-CCTCCGGCCTTTATGTCTTAG-3′ and the reverse 5′-TTGGGCTGCAGTATACAACG-3′. PCR reactions contained 1X Buffer GoTaq, 2.5 mM of each dNTPs, 0.3 μM of each primer, 0.03 U/μL of GoTaq DNA polymerase (Promega Corporation; Madison, WI, USA), 30 ng of genomic DNA, and H2O to a final volume of 25 μL. PCR amplification was carried out as follows: 95 °C for 2 min, followed by 35 cycles of 95 °C for 30 s, 62 °C for 30 s, 72 °C for 45 s, and then 72 °C for 5 min. The PCR fragments were purified using exonuclease I and alkaline phosphatase (ExoSAP-IT enzymatic system-USB Corporation, Cleveland, OH, USA) and subsequently Sanger-sequenced with the forward primer 5′-GCCAAACTACTCACCAGAAA-3′ for the locus YE3, 5′-GATTACCTCCTGGGACAAC-3′ for the locus YE17, and 5′-TAAAAACCTGTGGAAGGATAA-3′ for the locus YXX. Sequences were, respectively, aligned to the
Equus caballus haplotype HT1 Y chromosome locus YE3 (JX646942.1), locus YE17 (JX646950.1), and locus YXX (JX647030.1) for the haplotype annotation, and their evolutionary relationships were evaluated through a median-joining tree built using Network software v.10.0 [
17].
2.2. Mitochondrial DNA Control-Region Analysis
To provide a more comprehensive overview of the autochthonous horse breeds in Sardinia, the DnaSP 5.1 software [
18] was used to estimate the haplotype variability of the mitochondrial control-region sequences reported in Cardinali et al., 2016 [
7] and Giontella et al., 2020 [
14], belonging to Giara (GI;
N = 39), Sarcidano (SC;
N = 30), and Sardinian Anglo-Arab (AA;
N = 56) (
Table S2). Then, we evaluated the evolutionary relationships among the three breeds through the Network software v.10.0 [
17], by including also the other Giara (
N = 30) and Sarcidano (
N = 23) sequences available from GenBank [
15,
16] (
Table S2).
3. Results
3.1. Y-Chromosome Analysis
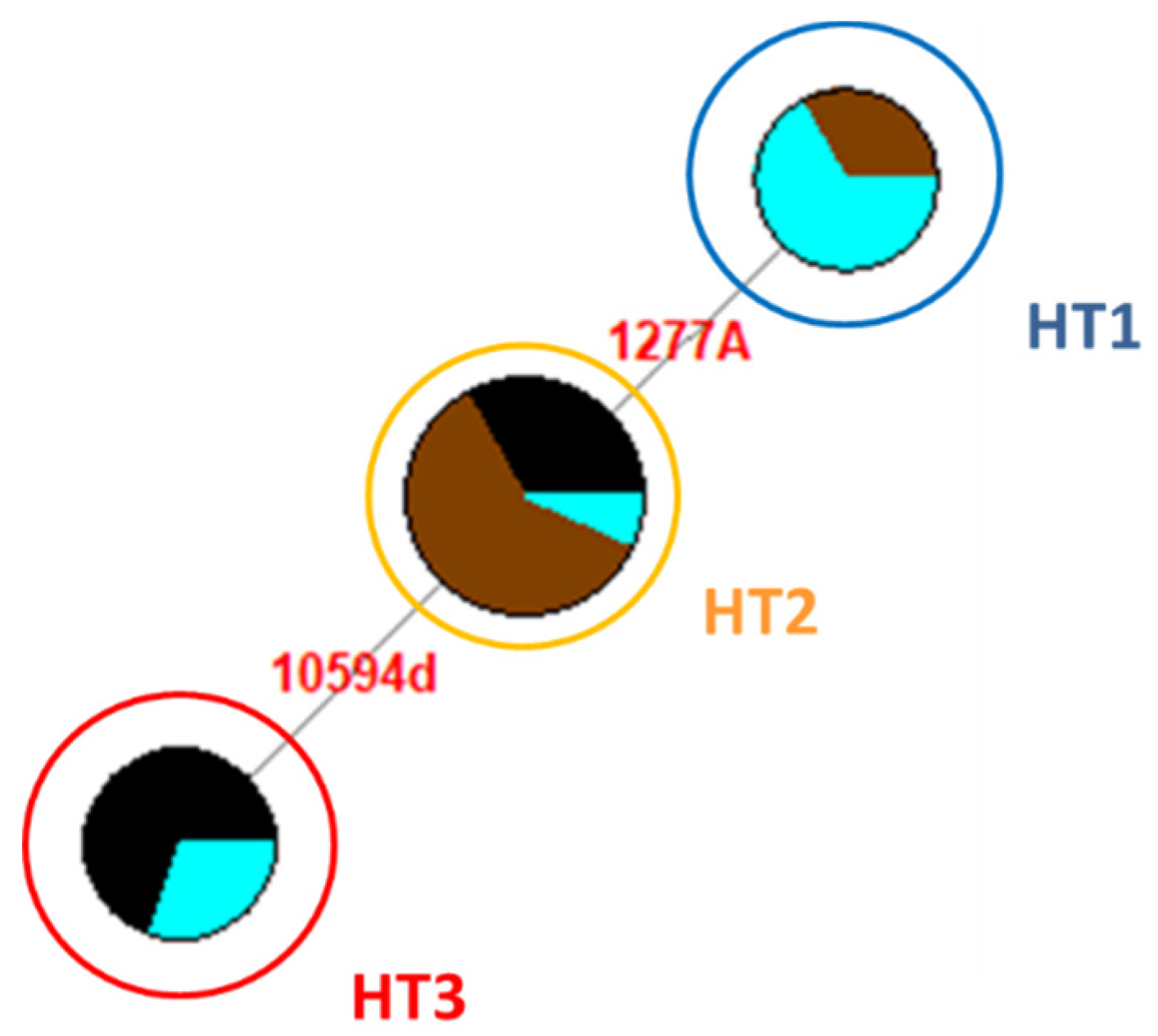
The alignment of the 34 NRY sequences to the published reference sequences (JX646942.1, JX646950.1 and JX647030.1), from the nucleotide position (np) 10,592 to np 11,330 for the locus YE3, from nps 1240 to 1410 for YE17, and from nps 25,342 to 25,480 for YXX, showed three different haplotypes (HT1, HT2, and HT3) [
6] (
Table S1). None of the samples presented mutations at the locus YXX. Considering all three loci, no mutations (HT1) have been identified in eight samples (three Giara and five Sarcidano). The HT2, characterized by a single nucleotide mutation (1277A), was found in 15 samples (nine Giara, one Sarcidano, and five Sardinian Anglo-Arab), while the HT3, differentiated from HT2 by a deletion in np 10,594, was found in 11 stallions (four Sarcidano and seven Sardinian Anglo-Arab) (
Figure 1).
The Sarcidano is the only breed showing all three haplotypes. On the other hand, each breed is characterized by a most represented HT: HT1 in Sarcidano, HT2 in Giara, and HT3 in Sardinian Anglo-Arab (
Figure 2).
3.2. Mitochondrial DNA Control-Region Analysis
The overall alignment of 610 base pairs, from np 15,491 to np 16,100, of the 125 control-region sequences from Cardinali et al., 2016 [
7] and Giontella et al., 2020 [
14] showed a haplotype diversity (Hd) of 0.944, with a total of 38 haplotypes and 49 polymorphic sites (S) detected (
Table 2).
We observed similar values for Giara and Sarcidano in terms of haplotype diversity (0.883 and 0.828, respectively) and number of haplotypes (Nh; nine and eight, respectively), and higher values for Sardinian Anglo-Arab (Hd = 0.969; Nh = 30).
By analyzing a shorter mtDNA control region (from np 15,491 to np 15,740), in order to add another 53 published sequences from Giara and Sarcidano, we observed a total of 47 haplotypes, of which nine were shared between two or more Sardinian breeds while the other 38 were present in only one breed: 11 for Giara, 10 for Sarcidano, and 17 in Sardinian Anglo-Arab (
Figure 3).
All haplotypes were classified in haplogroups, except for three sequences (JF804119, JF804147, and JF804151) from Morelli et al., 2014 [
15], and a total of eight (A, B, E, G, I, L, M, and N) mtDNA lineages were identified, with the highest frequencies for G, L, and I (37%, 24%, and 21%, respectively) (
Table S3). The highest maternal variability was found in Sardinian Anglo-Arab, which showed all eight haplogroups, followed by Giara, with six mtDNA lineages, and then by Sarcidano, presenting only four haplogroups (
Figure 4).
4. Discussion
Even if whole-genome approaches are now opening up new clues on the livestock genetic complexity and admixture, the two uniparental systems are still widely used to solve questions about breed origins and demographic processes. The traceability and characterization of female and male livestock lineages also offer a unique opportunity to conserve genetic resources and to promote and defend local breeds [
19,
20,
21]. This approach is compelling for horses [
9,
22] and even more important for animals that are bred in semi-feral conditions when few genealogical data are available. In this perspective, centers for breed preservation (in situ ed ex situ) have been established for Giara and Sarcidano, where individuals genetically characterized for both parental lines and specifically selected are kept for the management and conservation of local genetic resources.
4.1. Y-Chromosome Analysis
The current Y-chromosomal diversity of Sardinian horse breeds is linked to three haplotypes (HT1-3) that have been previously identified, at relatively high frequencies, across a broad range of horse breeds [
6]. The ancestral HT1 was found in Giara and Sarcidano horses, while HT2, considered a marker of the Neapolitan/Oriental wave, was present in all three Sardinian breeds. Finally, HT3, representing the Thoroughbred wave, was found in Sarcidano and Sardinian Anglo-Arab. Among our samples, the most frequent Y-chromosomal haplotype was HT2 (44%), whose distribution in Central Europe is in agreement with the spreading of Neapolitan and Oriental stallions from the Middle East to Central and Western Europe [
6]. Despite the fact that Sarcidano was the only Sardinian breed presenting all three haplotypes, it had the lowest frequency for HT2 (10%).
The presence of only six haplotypes in European horses and the low microsatellite variability [
6,
23,
24,
25] testify for a decline of the male effective population size and for the consequent decrease of the NRY variability [
13]. The practice of importing stallions (Arab and Thoroughbred) from foreign countries to improve local herds often caused the complete replacement of autochthonous Y chromosomes [
13]. This could be observed also in our dataset, where each breed is characterized by distinctive haplotype frequencies that probably highlight the consequences of three paternal introgressions of imported stallions into local breeds (
Figure 2). Giara shows a high percentage (75%) of HT2, frequent in Central and Eastern Europe, but absent in the breeds from Northern Europe and Iberian Peninsula, thus confirming genetic traces of horses with Middle Eastern origins, as reported by Gratani [
26]. The Sardinian Anglo-Arab shows the highest frequency (58%) for HT3, typical of Thoroughbred (derived from Oriental lineages and responsible for the predominance of this haplotype in modern horses) and distributed across many warm-blood horse breeds. This prevalence confirms that the Sardinian Anglo-Arab is a breed involved in breeding programs to improve the sport aptitude [
27]. The prevalence (50%) of the ancestral haplotype HT1 in Sarcidano confirms its uncertain (perhaps ancient) origins.
4.2. Mitochondrial DNA Control-Region Analysis
The unexpected high value of the overall haplotype diversity in Sardinian breeds is mostly driven by the Sardinian Anglo-Arab that presents 31 of the 38 haplotypes identified in our dataset (
Table 2). The lower number of haplotypes identified in Giara and Sarcidano (Nh = 9 and 8, respectively) is widely shared among these breeds, thus confirming the geographically isolated context of the island, as previously reported [
7,
15], and indicating common maternal local sources.
To include all available data, for a total of 178 horses, we performed the network analysis after sequence trimming to make the mtDNA dataset homogeneous. The analysis revealed a total of 47 haplotypes, with 38 unique mtDNA control-region sequences (
Figure 3). Among the other nine haplotypes, two were shared between Giara and Sarcidano horses, three were present only in Giara and Sardinian Anglo-Arab, and four were shared among the three breeds, testifying for a marginal impact of the latter breed on the original mtDNA gene pool of Giara and Sarcidano.
The haplogroup classification of all Sardinian sequences showed a total of eight horse maternal lineages out of the 17 previously reported [
5] with one main lineage for each breed (
Figure 4): haplogroup G mostly identified in Giara horses (68%), haplogroup I mainly represented in Sarcidano (41%), and haplogroup L, frequent in Sardinian Anglo-Arab (41%) (
Table S3). G and I haplogroups are more common in Asia and the Middle East, respectively [
5]. The percentage of L in Sardinian Anglo-Arab is considerably higher than that recorded in Western Asia (18%) and Continental Europe (31%) [
7].
4.3. Combining Information from Both Uniparental Genetic Systems
The Y-chromosome HT2 and the high number of maternally inherited haplotypes identified in the Sardinian Anglo-Arab breed suggest a sex-bias in the breeding program, where probably stallions from a unique breed (Thoroughbred) have been crossed with mares of different origins, a common breeding procedure used to improve local breeds with Thoroughbred thus creating half-breed individuals. The high mtDNA variability found in the Sardinian Anglo-Arab could be explained by the ancient migratory events that in the past have reached Sardinia and brought mares of different origins (especially old Arabian lines and Part-Arab) [
14].
The semi-feral Sarcidano breed was probably less impacted by these breeding programs and conserved the highest NRY variability and strong traces of the ancestral haplotype HT1. Likewise, on the maternal side, it shows the four most common mtDNA haplogroups in current Italian breeds [
7].
In Giara the high incidence of the mtDNA haplogroup G together with the results obtained from the NRY analysis confirms the Middle Eastern origin of this breed and its subsequent isolation in the island. Contemporary Giara horses are probably descendants of the ancient Phoenicia, in agreement with Gratani [
26] and many non-official records stating that these horses were introduced in Sardinia by Phoenician in the first millennium BCE. Recent studies focusing on the genetic history of human Sardinian populations inferred extensive exchange and continuity between the Phoenician population and broader Sardinia [
28,
29], thus supporting the hypothesis that Phoenician sailors might have brought their horses during their maritime expansions across the Mediterranean Sea and established settlements along the southern shores of Sardinia. The Giara horses are now the only survivors of a race that until the Middle Ages was much more extensive. The isolated environment of the Giara plateau presumably contributed to preservation of the breed’s genetic structure.
5. Conclusions
Sardinia, located to the west of Italy in the Mediterranean Sea, is an island characterized by many autochthonous species. Among them, there are three horse breeds native to this island and classified “at risk”, which were the objective of the present study. The peculiar Y-chromosomal distribution together with the high frequencies of certain mtDNA haplogroups highlights the genetic characteristics of these endangered local breeds.
Each breed showed a distinctive NRY haplotype: the ancestral one (HT1), most represented in Sarcidano; the HT2, marker of the Neapolitan/Oriental wave, above all found in Giara, and HT3, legacy of the Thoroughbred wave, prominent in the Sardinian Anglo-Arab. As for the female counterpart, the haplotype sharing between Giara and Sarcidano supports inferences of common maternal origins and the high incidence of the four most frequent lineages in present-day Italian horse breeds (G, I, L, and M) suggests female inputs from the peninsula. However, Giara shows strong genetic relationships to ancient Eastern Mediterranean sources, corroborating the hypothesis of a common past migration traced back to Phoenicians: both humans and horses could have reached the island from the Eastern Mediterranean area by sea in the first millennium BCE.
The low variability in Giara and Sarcidano confirms the marginal impact of crossbreeding on the indigenous mitochondrial gene pools and the lack of recent gene flow from outside into Sardinia. On the other hand, the high haplotype variability reported for Sardinian Anglo-Arab indicates the presence of multiple mare lineages in the current population.
This study suggests that despite their endangered status, these Sardinian horse breeds preserve significant reservoirs of genetic diversity. Distinctive variants and/or lineages have been identified when evaluating the two uniparental genetic markers. Therefore, targeted conservation efforts that take into account both female and male lines of descent are required to safeguard the extant genetic variability and to establish improvements in the selection programs.
Supplementary Materials
The following are available online at
https://www.mdpi.com/2076-2615/10/9/1544/s1, Table S1: List of 34 male horses here analyzed and their Y-chromosomal haplotypes, Table S2: List of 178 mtDNA control regions belonging to the Sardinian horse breeds included in the analyses, Table S3: Haplogroup frequencies (%) distribution for each breed based on 175 mtDNA control regions.
Author Contributions
Conceptualization, A.G., I.C., A.A., M.S., and H.L.; formal analysis, A.G. and I.C.; data curation, C.P. and G.P.B.; writing—original draft preparation, A.G., I.C., A.A., M.S., and H.L.; writing—review and editing, I.C., C.P., R.C., G.P.B., and M.S.; supervision, A.A., M.S., and H.L.; project administration, A.G., M.S., and R.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
A special word of thanks is due to AGRIS Sardegna, the former Istituto Incremento Ippico della Sardegna, Sardinian Regional Department for Agriculture and Agro-Pastoral Reform, C.R.P. Sardinian Regional Programming Centre and ANACAAD, the National Association of Anglo-Arab and part-bred Anglo-Arab Horse. A.A. received support from the Italian Ministry of Education, University and Research (MIUR) for the Dipartimenti di Eccellenza Program (2018–2022).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Arquer, S. Sardiniae Brevis Historia et Descripitio; CUEC: Cagliari, Italia, 2008. [Google Scholar]
- Gratani, L. L’Anglo-Arabo Sardo; Equestri: Milano, Italy, 1988. [Google Scholar]
- Giontella, A.; Sarti, F.M.; Biggio, G.P.; Giovannini, S.; Cherchi, R.; Pieramati, C.; Silvestrelli, M. Genetic Parameters and Inbreeding Effect of Morphological Traits in Sardinian Anglo Arab Horse. Animals 2020, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Giontella, A.; Sarti, F.M.; Biggio, G.P.; Giovannini, S.; Cherchi, R.; Silvestrelli, M.; Pieramati, C. Elo Method and Race Traits: A New Integrated System for Sport Horse Genetic Evaluation. Animals 2020, 10, 1145. [Google Scholar] [CrossRef] [PubMed]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Hooshiar Kashani, B.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallner, B.; Vogl, C.; Shukla, P.; Burgstaller, J.P.; Druml, T.; Brem, G. Identification of genetic variation on the horse Y chromosome and the tracing of male founder lineages in modern breeds. PLoS ONE 2013, 8, e60015. [Google Scholar] [CrossRef] [Green Version]
- Cardinali, I.; Lancioni, H.; Giontella, A.; Capodiferro, M.R.; Capomaccio, S.; Buttazzoni, L.; Biggio, G.P.; Cherchi, R.; Albertini, E.; Olivieri, A.; et al. An Overview of Ten Italian Horse Breeds through Mitochondrial DNA. PLoS ONE 2016, 11, e0153004. [Google Scholar] [CrossRef] [Green Version]
- Kakoi, H.; Kikuchi, M.; Tozaki, T.; Hirota, K.I.; Nagata, S.I.; Hobo, S.; Takasu, M. Distribution of Y chromosomal haplotypes in Japanese native horse populations. J. Equine Sci. 2018, 29, 39–42. [Google Scholar] [CrossRef]
- Khaudov, A.D.; Duduev, A.S.; Kokov, Z.A.; Amshokov, K.K.; Zhekamukhov, M.K.; Zaitsev, A.M.; Reissmann, M. Genetic analysis of maternal and paternal lineages in Kabardian horses by uniparental molecular markers. Open Vet. J. 2018, 8, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Wutke, S.; Sandoval-Castellanos, E.; Benecke, N.; Döhle, H.J.; Friederich, S.; Gonzalez, J.; Hofreiter, M.; Lõugas, L.; Magnell, O.; Malaspinas, A.S.; et al. Decline of genetic diversity in ancient domestic stallions in Europe. Sci. Adv. 2018, 4, eaap9691. [Google Scholar] [CrossRef] [Green Version]
- Lancioni, H.; Cardinali, I.; Giontella, A.; Antognoni, M.T.; Miglio, A. Mitochondrial DNA variation in the Italian Heavy Draught Horse. PeerJ 2020, 15, e8996. [Google Scholar] [CrossRef]
- Giontella, A.; Cardinali, I.; Lancioni, H.; Giovannini, S.; Pieramati, C.; Silvestrelli, M.; Sarti, F.M. Mitochondrial DNA Survey Reveals the Lack of Accuracy in Maremmano Horse Studbook Records. Animals 2020, 10, 839. [Google Scholar] [CrossRef]
- Wallner, B.; Palmieri, N.; Vogl, C.; Rigler, D.; Bozlak, E.; Druml, T.; Jagannathan, V.; Leeb, T.; Fries, R.; Tetens, J.; et al. Chromosome Uncovers the Recent Oriental Origin of Modern Stallions. Curr. Biol. 2017, 27, 2029–2035.E5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giontella, A.; Sarti, F.M.; Cardinali, I.; Giovannini, S.; Cherchi, R.; Lancioni, H.; Silvestrelli, M.; Pieramati, C. Genetic Variability and Population Structure in the Sardinian Anglo-Arab Horse. Animals 2020, 10, 1018. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Useli, A.; Sanna, D.; Barbato, M.; Contu, D.; Pala, M.; Cancedda, M.; Francalacci, P. Mitochondrial DNA lineages of Italian Giara and Sarcidano horses. Genet. Mol. Res. 2014, 13, 8241–8257. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, M.C.; Strillacci, M.G.; Valiati, P.; Bighignoli, B.; Cancedda, M.; Zanotti, M. Mitochondrial D-loop sequence variation among Italian horse breeds. Genet. Sel. Evol. 2004, 36, 663–672. [Google Scholar] [CrossRef] [Green Version]
- Network Software. Available online: https://www.fluxus-engineering.com/ (accessed on 17 March 2020).
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Lenstra, J.A.; Groeneveld, L.F.; Eding, H.; Kantanen, J.; Williams, J.L.; Taberlet, P.; Nicolazzi, E.L.; Sölkner, J.; Simianer, H.; Ciani, E.; et al. Molecular tools and analytical approaches for the characterization of farm animal genetic diversity. Anim. Genet. 2012, 43, 483–502. [Google Scholar] [CrossRef] [PubMed]
- Kanthaswamy, S. Review: Domestic animal forensic genetics–biological evidence, genetic markers, analytical approaches and challenges. Anim. Genet. 2015, 46, 473–484. [Google Scholar] [CrossRef]
- Di Lorenzo, P.; Lancioni, H.; Ceccobelli, S.; Curcio, L.; Panella, F.; Lasagna, E. Uniparental genetic systems: A male and a female perspective in the domestic cattle origin and evolution. Electron. J. Biotechnol. 2016, 23, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Bodo, I.; Alderson, L.; Langlois, B. (Eds.) Conservation Genetics of Endangered Horse Breeds, 1st ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2005. [Google Scholar]
- Kreutzmann, N.; Brem, G.; Wallner, B. The domestic horse harbours Y-chromosomal microsatellite polymorphism only on two widely distributed male lineages. Anim. Genet. 2014, 45, 460. [Google Scholar] [CrossRef]
- Han, H.; Zhang, Q.; Gao, K.; Yue, X.; Zhang, T.; Dang, R.; Lan, X.; Chen, H.; Lei, C. Y-single nucleotide polymorphisms diversity in Chinese indigenous horse. Asian Australas J. Anim. Sci. 2015, 28, 1066–1074. [Google Scholar] [CrossRef]
- Han, H.; Wallner, B.; Rigler, D.; MacHugh, D.E.; Manglai, D.; Hill, E.W. Chinese Mongolian horses may retain early domestic male genetic lineages yet to be discovered. Anim. Genet. 2019, 50, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Gratani, L. Il Cavallo della Giara, Istituto di Incremento Ippico della Sardegna; RAS: Cagliari, Italy, 1980. [Google Scholar]
- Giontella, A.; Pieramati, C.; Silvestrelli, M.; Sarti, F.M. Analysis of founders and performance test effects on an autochthonous horse population through pedigree analysis: Structure, genetic variability and inbreeding. Animal 2019, 13, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Matisoo-Smith, E.; Gosling, A.L.; Platt, D.; Kardailsky, O.; Prost, S.; Cameron-Christie, S.; Collins, C.J.; Boocock, J.; Kurumilian, Y.; Guirguis, M.; et al. Ancient mitogenomes of Phoenicians from Sardinia and Lebanon: A story of settlement, integration, and female mobility. PLoS ONE 2018, 13, e0190169. [Google Scholar] [CrossRef] [PubMed]
- Marcus, J.H.; Posth, C.; Ringbauer, H.; Lai, L.; Skeates, R.; Sidore, C.; Beckett, J.; Furtwängler, A.; Olivieri, A.; Chiang, C.W.K.; et al. Genetic history from the Middle Neolithic to present on the Mediterranean island of Sardinia. Nat. Commun. 2020, 11, 939. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}