Friendship or Competition? Symmetry in Social Play within the Two Packs of German Shepherd Puppies

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Subjects of the Study
2.2. Data Collection, Behavioural Observations, and Analysis
2.3. Play Observations
2.4. Testing of the 50:50 Ratio
3. Results
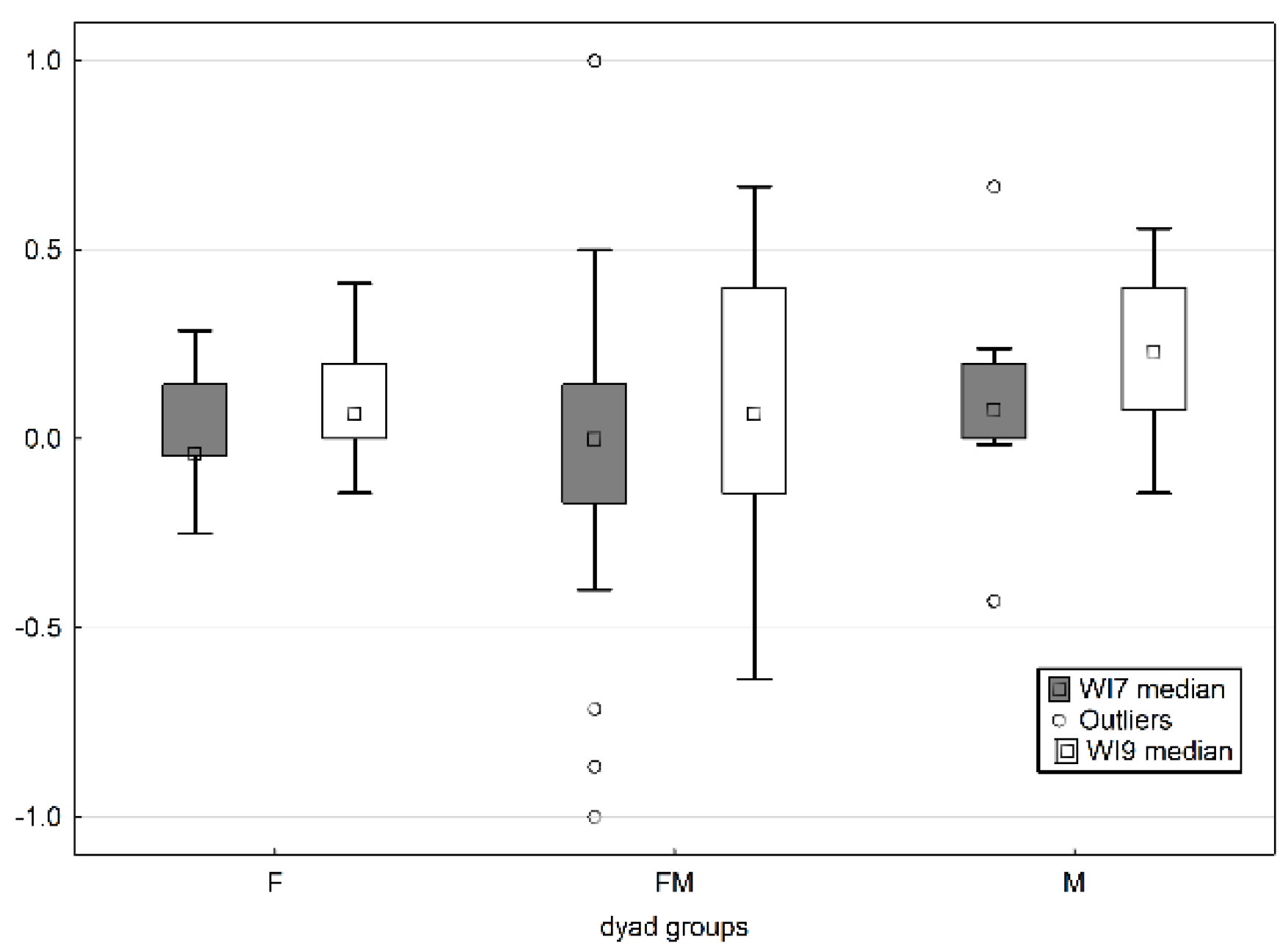
3.1. Play Interactions within the Litter in Relation to the Sex of Puppies
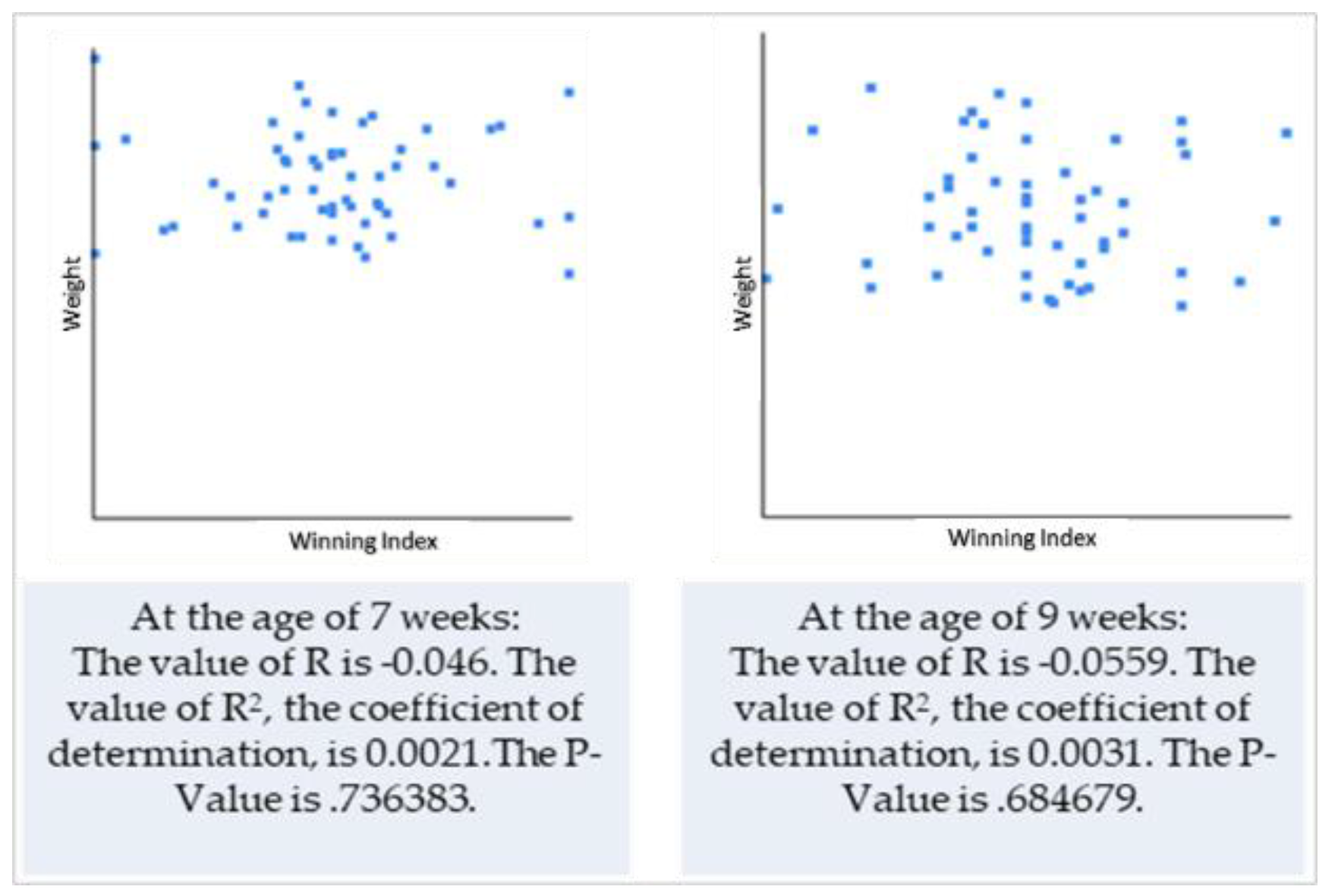
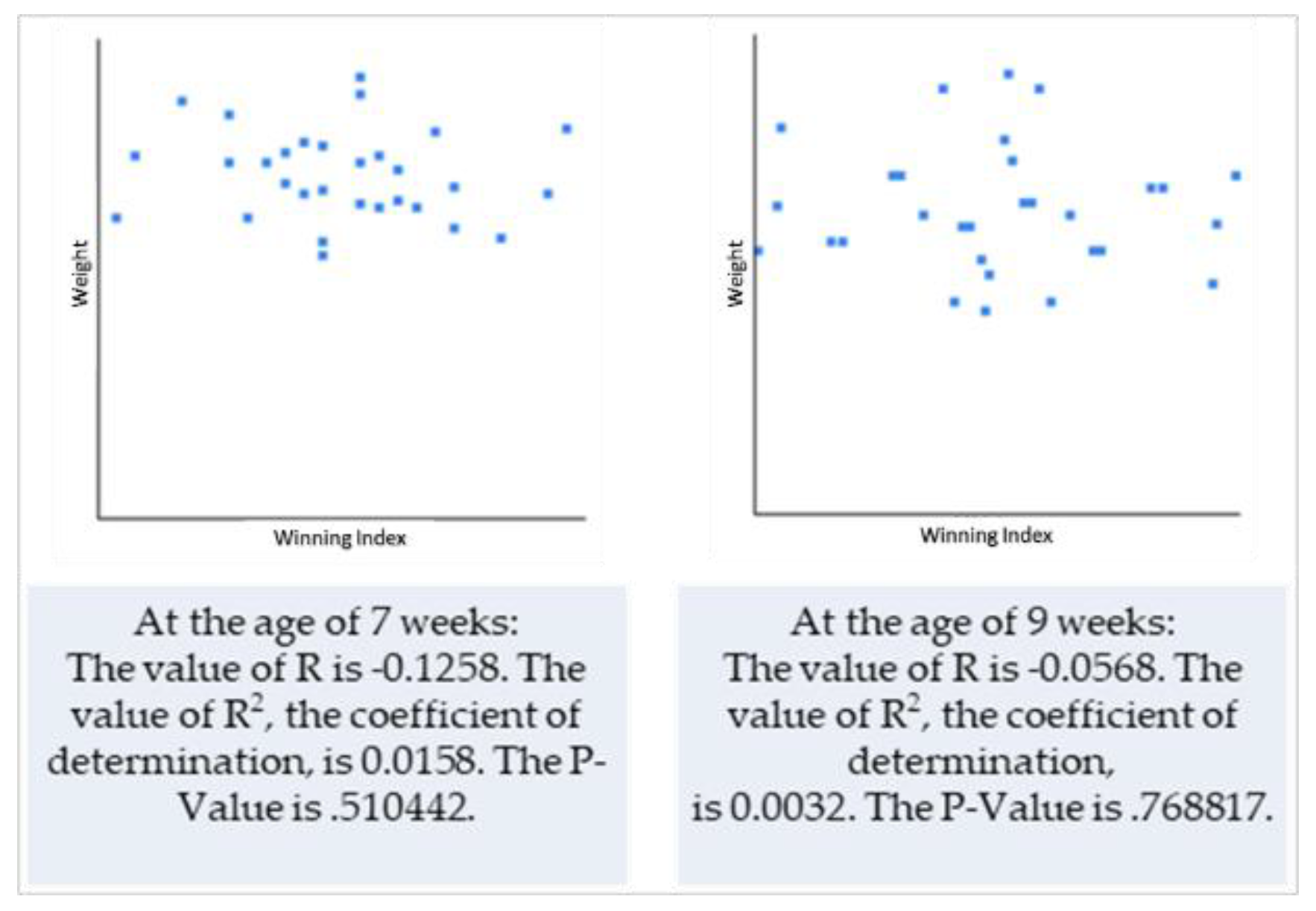
3.2. Dyadic Play Interactions within the Litter in Relation to Size (Weight vs. Chest Circumference) of Puppies
3.3. Testing the 50:50 Rule
4. Discussion
4.1. Play Interactions within the Litter in Relation to the Sex of Puppies
4.2. Dyadic Play Interactions within the Litter in Relation to Size (Weight vs. Chest Circumference) of Puppies
4.3. Testing the 50:50 Rule
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bekoff, M.; Byers, J.A. A critical reanalysis of the ontogeny of mammalian social and locomotor play. An ethological hornet’s nest. In Behavioral Development; The Bielefeld Interdisciplinary Project; Immelmann, K., Barlow, G.W., Petrinovich, L., Main, M., Eds.; Cambridge University Press: Cambridge, MA, USA, 1981; pp. 296–337. [Google Scholar]
- Fagen, R.M. Animal Play Behavior; Oxford University Press: Oxford, UK, 1981; p. 684. [Google Scholar]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits, 1st ed.; A Bradford Book; MIT Press: Cambridge, MA, USA, 2006; p. 501. [Google Scholar]
- Bekoff, M. Social play and play-soliciting by infant canids. Am. Zool. 1974, 14, 323–340. [Google Scholar] [CrossRef]
- Ward, C.; Bauer, E.B.; Smuts, B.B. Partner preferences and asymmetries in social play among domestic dogs, Canis lupus familiaris, littermates. Anim. Behav. 2008, 76, 1187–1199. [Google Scholar] [CrossRef]
- Aloff, B. Canine Body Language. A Photographic Guide Interpreting the Native Language of Domestic Dog; Dogwise Publishing: Wenatchee, WA, USA, 2005; p. 157. [Google Scholar]
- Handelman, B. Canine Behaviour: A Photo Illustrated Handbook; Dogwise Publishing: Wenatchee, WA, USA, 2008; p. 347. [Google Scholar]
- Špinka, M.; Newberry, R.C.; Bekoff, M. Mammalian play: Training for the unexpected. Q Rev. Biol. 2001, 76, 141–168. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, L.J.M.J. The neurobiology of social reward: Insight from social play behaviour in adolescent rats. Biol. Psychiatry 2011, 69, 103S. [Google Scholar]
- Horowitz, A.; Hecht, J. Examining dog-human play: The characteristics, effect, and vocalizations of a unique interspecific interaction. Anim. Cogn. 2016, 19, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Sommerville, R.; O’Connor, E.A.; Asher, L. Why dogs play? A review of the function of play in the domestic dog. App. Anim. Behav. Sci. 2017, 197, 1–8. [Google Scholar] [CrossRef]
- Rooney, N.J.; Bradshaw, J.W.S.; Robinson, I.H. A comparison of dog-dog and dog-human play behaviour. App. Anim. Behav. Sci. 2000, 66, 235–248. [Google Scholar] [CrossRef]
- Scott, J.P.; Fuller, J.L. Dog Behaviour: The Genetic Basis; The University of Chicago Press: Chicago, IL, USA, 1965; p. 506. [Google Scholar]
- Scott, J.P.; Marston, M.V. Critical periods affecting the development of normal and mal-adjustive social behaviour of puppies. Pedagog. Semin. J. Genet. Psychol. 1950, 77, 26–60. [Google Scholar] [CrossRef]
- Byosiere, S.-E.; Espinoza, J.; Marshall-Pescini, S.; Smuts, B.; Range, F. Investigating the function of play bows in dog and wolf puppies (Canis lupus familiaris, Canis lupus occidentalis). PLoS ONE 2016, 11, e0168570. [Google Scholar] [CrossRef]
- Pellis, S.M.; Pellis, V.C. The Playful Brain: Venturing to the Limits of Neuroscience; Oneworld Press: Oxford, UK, 2009; p. 288. [Google Scholar]
- Coppinger, R.P.; Smith, C.K. A model for understanding the evolution of mammalian behaviour. In Current Mammalogy; Genoways, H., Ed.; Plenum Press: New York, NY, USA, 1990; pp. 335–374. [Google Scholar]
- Biben, M. Squirrel monkey play-fighting: Making the case for a cognitive training function for play. In Animal Play: Evolutionary, Comparative and Ecological Perspectives; Bekoff, M., Byers, J.A., Eds.; Cambridge University Press: Cambridge, MA, USA, 1998; pp. 217–244. [Google Scholar]
- Bayers, J.A. Biological effects of Locomotor Play: Getting into shape, or something more specific? In Animal Play: Evolutionary, Comparative and Ecological Perspectives; Bekoff, M., Byers, J.A., Eds.; Cambridge University Press: Cambridge, MA, USA, 1998; pp. 205–220. [Google Scholar]
- Zahavi, A. The testing a bond. Anim. Behav. 1977, 25, 246–247. [Google Scholar] [CrossRef]
- Bekoff, M.; Allen, C. Intentional communication and social play: How and why animals negotiate and agree to play. In Animal Play: Evolutionary, Comparative and Ecological Perspectives; Bekoff, M., Byers, J.A., Eds.; Cambridge University Press: Cambridge, MA, USA, 1998; pp. 97–114. [Google Scholar]
- Bekoff, M. Social play behaviour: Cooperation, fairness, trust and evolution of morality. J. Conscious. Stud. 2001, 8, 81–90. [Google Scholar]
- Pozis-Francois, O.; Zahavi, A.; Zahavi, A. Social play in Arabian Babblers. Behaviour 2004, 141, 425–450. [Google Scholar]
- Bekoff, M. Social play behaviour. Bioscience 1984, 84, 228–233. [Google Scholar] [CrossRef]
- Dehasse, J. Sensory, emotional and social development of the young dog. Bull. Vet. Clin. Ethol. 1994, 2, 6–29. [Google Scholar]
- Case, L.P. The Dog: Its Behaviour, Nutrition and Health, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2005; p. 496. [Google Scholar]
- Bekoff, M. The Development of Social Interaction, Play, and Metacommunication in Mammals: An Ethological Perspective. Q. Rev. Biol. 1972, 47, 412–434. [Google Scholar] [CrossRef]
- Meyer, S.; Weber, J.-M. Ontogeny of Dominance in Free-living Red Foxes. Ethology 1996, 102, 1008–1019. [Google Scholar] [CrossRef]
- Fuller, J.L.; Fox, M.W. The behaviour of dogs. In The Behaviour of Domestic Animals, 2nd ed.; Hafez, E.S.E., Ed.; Williams & Wilkins: Baltimore, MD, USA, 1969; p. 438. [Google Scholar]
- McBride, A. The human-dog relationship. In The Waltham Book of Human-Animal Interactions: Benefits and Responsibilities of Pet Ownership; Robinson, I., Ed.; Pergamon Press: New York, NY, USA, 1995; pp. 99–112. [Google Scholar]
- Altmann, S.A. Social behaviour of anthropoid primates: Analysis of recent concepts. In Roots of Behavior; Bliss, E.L., Ed.; Harper: New York, NY, USA, 1962; pp. 277–285. [Google Scholar]
- Aldis, O. Play Fighting; Academic Press: New York, NY, USA, 1975; p. 319. [Google Scholar]
- Bekoff, M. Wild justice and fair play: Cooperation, forgiveness and morality in animals. Biol. Philos. 2004, 19, 489–520. [Google Scholar] [CrossRef]
- Owens, N.W. Social play behaviour in free-living baboons, Papio anubis. Anim. Behav. 1975, 23, 387–408. [Google Scholar] [CrossRef]
- Bauer, E.B.; Smuts, B.B. Cooperation and Competition during Dyadic Play in Domestic Dogs, Canis familiaris. Anim. Behav. 2007, 73, 489–499. [Google Scholar] [CrossRef]
- Essler, J.L.; Cafazzo, S.; Marshall-Pescini, S.; Virányi, Z.; Kotrschal, K.; Range, F. Play behaviour in Wolves: Using the ‘50:50’ Rule to test for Egalitarian Play Styles. PLoS ONE 2016, 11, e0154150. [Google Scholar] [CrossRef]
- Lund, J.D.; Vestergaard, K.S. Development of social behaviour in four litters of dogs (Canis familiaris). Acta Vet. Scand. 1998, 39, 183–193. [Google Scholar] [PubMed]
- Pal, S.K. Play behaviour during early ontogeny in free-ranging dogs (Canis familiaris). Appl. Anim. Behav. Sci. 2010, 126, 140–153. [Google Scholar] [CrossRef]
- Bonanni, R.; Cafazzo, S.; Abis, A.; Barillari, E.; Valsecchi, P.; Natoli, E. Age-graded dominance hierarchies and social tolerance in packs of free-ranging dogs. Behav. Ecol. 2017, 28, 1004–1020. [Google Scholar] [CrossRef]
- Range, F.; Virányi, Z. Tracking the evolutionary origins of dog-human cooperation: The “Canine Cooperation Hypothesis”. Front. Psychol. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Mech, L.D. The Wolf: Ecology and Behaviour of Endangered Species; Natural History Press: New York, NY, USA, 1970; p. 385. [Google Scholar]
- Mech, L.D.; Boitani, L. Wolves: Behavior, Ecology and Conservation, 2nd ed.; University Chicago Press: Chicago, IL, USA, 2003; p. 448. [Google Scholar]
- Lord, K.; Feinstein, M.; Smith, B.; Coppinger, R. Variation in reproductive traits of members of the genus Canis with the special attention to the domestic dog (Canis familiaris). Behav. Processes 2013, 92, 131–142. [Google Scholar] [CrossRef]
- Serpell, J. The Domestic Dog: Its Evolution, Behaviour and Interactions with People; Cambridge University Press: Cambridge, UK, 1995; p. 284. [Google Scholar]
- Wright, J.C. The development of social structure during the primary socialization period in German shepherds. Dev. Psychobiol. 1980, 13, 17–24. [Google Scholar] [CrossRef]
- van der Borg, J.A.M.; Schilder, M.B.H.; Vinke, C.M.; de Vries, H. Dominance in Domestic Dogs: A Quantitative Analysis of Its Behavioural Measures. PLoS ONE 2015, 10, e0133978. [Google Scholar] [CrossRef]
- Trisko, R.K.; Smuts, B.B. Dominance relationships in a group of domestic dogs (Canis lupus familiaris). Behaviour 2015, 152, 677–704. [Google Scholar] [CrossRef]
- Feddersen-Petersen, D. The ontogeny of social play and agonistic behaviour in selected canine species. Bonn. Zool. Beitr. 1991, 42, 97–114. [Google Scholar]
- Feddersen-Petersen, D. Social behaviour of dogs and related canids. In The Behavioural Biology of Dogs; Jensen, P., Ed.; Cromwell Press: Trowbridge, UK, 2007; pp. 105–119. [Google Scholar]
- Pierantoni, L.; Albertini, M.; Pirrone, F. Prevalence of owner-reported behaviours in dogs separated from the litter at two different ages. Vet. Rec. 2011, 169, 468–473. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Litter | Birth | Dog ID | Sex | Weight (kg) | Chest Circumference (cm) | ||
|---|---|---|---|---|---|---|---|
| (month) | 7 weeks | 9 weeks | 7 weeks | 9 weeks | |||
| 1 | April | A | Female | 4.1 | 5.6 | 38 | 45 |
| B | Female | 4.9 | 7.0 | 39 | 43 | ||
| C | Male | 4.5 | 5.4 | 39 | 46 | ||
| D | Male | 5.2 | 7.1 | 42 | 47 | ||
| E | Female | 3.8 | 5.0 | 36 | 40 | ||
| F | Male | 4.2 | 5.1 | 37 | 40 | ||
| G | Male | 4.3 | 6.4 | 39 | 42 | ||
| H | Female | 4.5 | 6.7 | 38 | 46 | ||
| 2 | March | I | Female | 5.5 | 6.0 | 47 | 49 |
| J | Female | 5.2 | 5.1 | 47 | 47 | ||
| K | Male | 5.9 | 5.3 | 45 | 47 | ||
| L | Male | 6.7 | 7.5 | 50 | 51 | ||
| M | Female | 5.7 | 5.3 | 42 | 46 | ||
| N | Male | 5.4 | 5.8 | 45 | 48 | ||
| Behaviour (Attacks/Pursuits) | Definition |
|---|---|
| Forced down | W uses sufficient physical force to cause L to drop partially or completely to the ground; force may be applied with a bite, push/tackle, body slam or some other forceful movement. |
| Chin over | W places chin over L’s back, usually right behind the neck or near L’s shoulders, W’s chin may or may not touch L but W’s chin must be at or near a 90-degree angle in relation to the plane of L’s spine. |
| Mount | W rears up to place forelegs on L’s back in a front lateral or rear mount position, differentiated from push/tackles by W’s rounded spine position during the mount and distinct pause in the stable mount position, which may or may not be accompanied by thrusting. |
| Over | W sits on, stands over, or lies over L at least 25% of W’s torso over L torso. |
| Overs during downs | W stands over or lies on L with at least 50% of W’s torso over L’s torso (or vice versa: 50% of L’s torso is under W’s torso), or W sits and exerts weight directly on L’s head or torso with a distinct pause in the sitting position. |
| Paw on | W stands up on its hind legs and puts front legs on R’s shoulders, usually silent and with open mouth, individuals can bite each other. |
| Push/tackle | W rears up and places one or both front paws on L with forceful contact; forelimbs may or may not be wrapped around L’s body in a tackle grasp (if interaction results in a down, it is coded as forced down, instead of independent push/tackle). |
| Number of Interactions in Dyads | ||||||||
|---|---|---|---|---|---|---|---|---|
| Litter | Week | Total | Male vs. Male | Male vs. Female | Female vs. Female | |||
| pack | number | % | number | % | number | % | ||
| 1 | 7 | 339 | 61 | 18.00% | 210 | 61.94% | 68 | 20.05% |
| 9 | 433 | 75 | 17.30% | 261 | 60.27% | 97 | 22.40% | |
| 2 | 7 | 948 | 175 | 18.40% | 564 | 59.49% | 209 | 22.05% |
| 9 | 822 | 168 | 20.44% | 481 | 58.52% | 173 | 21.05% | |
| Both puppy Packs | Sex | Observed (No.) | Expected (No.) | Difference | x2 Value | p-Value | alpha < 0.05 |
| All interactions (n = 2542) | Same | 926 | 997 | −71.00 | 8.545 | 0.00347 | * significant |
| Opposite | 1616 | 1545 | 71.00 | ||||
| Puppy pack 1 | Sex | Observed (%) | Expected (%) | Difference | x2 value | p-value | alpha < 0.05 |
| Interaction of females | Same | 26 | 43 | −17.00 | 11.791 | 0.0006 | * significant |
| Opposite | 74 | 57 | 17.00 | ||||
| Interaction of males | Same | 22 | 43 | −21.00 | 17.993 | 0.00002 | * significant |
| Opposite | 78 | 57 | 21.00 | ||||
| Puppy pack 2 | Sex | Observed (%) | Expected (%) | Difference | x2 value | p-value | alpha < 0.05 |
| Interaction of females | Same | 27 | 40 | −13.00 | 7.042 | 0.00796 | * significant |
| Opposite | 73 | 60 | 13.00 | ||||
| Interaction of males | Same | 25 | 40 | −15.00 | 9.375 | 0.0022 | * significant |
| Opposite | 75 | 60 | 15.00 |
| Puppy Pack 1—Winning Indexes for Dyadic Play Interactions | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 7th Week of age | |||||||||||
| Puppy | A♀ | B♀ | C♂ | D♂ | E♀ | F♂ | G♂ | H♀ | WI (Mean) | Median | SD |
| A♀ | * | 0.167 | −0.23 | 1 | 0 | 0.2 | −0.06 | 0.29 | 0.1952857 | 0.167 | 0.3961341 |
| B♀ | −0.167 | * | 0 | −0.19 | 0.14 | −0.71 | 0.14 | −0.04 | −0.1181429 | −0.04 | 0.2918861 |
| C♂ | 0.23 | 0 | * | 0.67 | −0.13 | 0.2 | −0.43 | 0.5 | 0.1485714 | 0.2 | 0.3739621 |
| D♂ | −1 | 0.19 | −0.67 | * | 1 | 0.11 | 0 | −0.4 | −0.11 | 0 | 0.6547773 |
| E♀ | 0 | −0.14 | 0.13 | −1 | * | −1 | −0.87 | −0.25 | −0.4471429 | −0.25 | 0.4926701 |
| F♂ | −0.2 | 0.71 | −0.2 | −0.11 | 1 | * | 0.08 | −0.08 | 0.1714286 | −0.08 | 0.4836124 |
| G♂ | 0.06 | −0.14 | 0.43 | 0 | 0.87 | −0.08 | * | 0.27 | 0.2014286 | 0.06 | 0.3559695 |
| H♀ | −0.29 | 0.04 | −0.5 | 0.4 | 0.25 | 0.08 | −0.27 | * | −0.0414286 | 0.04 | 0.322874 |
| 9th Week of age | |||||||||||
| Puppy | A♀ | B♀ | C♂ | D♂ | E♀ | F♂ | G♂ | H♀ | WI (Mean) | Median | SD |
| A♀ | * | 0 | −0.25 | 0.67 | 0.2 | 0 | 0.1 | 0.41 | 0.1614286 | 0.1 | 0.3019618 |
| B♀ | 0 | * | 0.11 | −0.14 | 0.07 | 0 | 0.2 | 0 | 0.0342857 | 0 | 0.1067485 |
| C♂ | 0.25 | −0.11 | * | 0.4 | −0.18 | 0.25 | −0.14 | 0.4 | 0.1242857 | 0.25 | 0.2585122 |
| D♂ | −0.67 | 0.14 | −0.4 | * | 0.4 | 0.06 | 0.08 | 0 | −0.0557143 | 0.06 | 0.3598081 |
| E♀ | −0.2 | −0.07 | 0.18 | −0.4 | * | −0.64 | −0.55 | −0.14 | −0.26 | −0.2 | 0.2874601 |
| F♂ | 0 | 0 | −0.25 | - | 0.64 | * | 0.23 | −0.16 | 0.0766667 | 0 | 0.32104 |
| G♂ | −0.1 | −0.2 | 0.14 | −0.08 | 0.55 | −0.23 | * | 0.14 | 0.0314286 | −0.08 | 0.2720557 |
| H♀ | −0.41 | 0 | 0.4 | 0 | 0.14 | 0.16 | −0.14 | * | 0.0214286 | 0 | 0.2548482 |
| Puppy Pack 2—Winning Indexes for Dyadic Play Interactions | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 7th Week of age | |||||||||
| Puppy | I♀ | J♀ | K♂ | L♂ | M♀ | N♂ | WI (Mean) | Median | SD |
| I♀ | * | −0.04 | −0.06 | −0.17 | −0.08 | −0.06 | −0.082 | −0.06 | 0.0511859 |
| J♀ | 0.04 | * | 0.1 | 0.02 | −0.04 | 0.02 | 0.028 | 0.02 | 0.0501996 |
| K♂ | 0.06 | −0.1 | * | 0.24 | 0.12 | 0.02 | 0.068 | 0.06 | 0.1253794 |
| L♂ | 0.17 | −0.02 | −0.24 | * | 0.12 | −0.02 | 0.002 | −0.02 | 0.1594365 |
| M♀ | 0.08 | 0.04 | −0.12 | −0.12 | * | 0.22 | 0.02 | 0.04 | 0.1442221 |
| N♂ | 0.06 | −0.02 | −0.02 | 0.02 | −0.22 | * | −0.036 | −0.02 | 0.108074 |
| 9th Week of age | |||||||||
| Puppy | I♀ | J♀ | K♂ | L♂ | M♀ | N♂ | WI (Mean) | Median | SD |
| I♀ | * | −0.04 | 0.25 | 0.02 | 0.27 | 0.57 | 0.214 | 0.25 | 0.241516 |
| J♀ | 0.04 | * | 0.07 | 0.03 | 0.09 | 0.62 | 0.17 | 0.07 | 0.2526856 |
| K♂ | −0.25 | −0.07 | * | 0.11 | −0.19 | 0.43 | 0.006 | −0.07 | 0.2740073 |
| L♂ | −0.02 | −0.03 | −0.11 | * | 0.14 | 0.56 | 0.108 | −0.02 | 0.2684586 |
| M♀ | −0.27 | −0.09 | 0.19 | −0.14 | * | 0.4 | 0.018 | −0.09 | 0.2716063 |
| N♂ | −0.57 | −0.62 | −0.43 | −0.56 | −0.4 | * | −0.516 | −0.56 | 0.095551 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kottferová, J.; Skurková, L.; Mesarčová, L.; Lešková, L.; Demeová, A.; Jakuba, T. Friendship or Competition? Symmetry in Social Play within the Two Packs of German Shepherd Puppies. Animals 2020, 10, 1627. https://doi.org/10.3390/ani10091627
Kottferová J, Skurková L, Mesarčová L, Lešková L, Demeová A, Jakuba T. Friendship or Competition? Symmetry in Social Play within the Two Packs of German Shepherd Puppies. Animals. 2020; 10(9):1627. https://doi.org/10.3390/ani10091627
Chicago/Turabian StyleKottferová, Jana, Lenka Skurková, Lýdia Mesarčová, Lenka Lešková, Alena Demeová, and Tomáš Jakuba. 2020. "Friendship or Competition? Symmetry in Social Play within the Two Packs of German Shepherd Puppies" Animals 10, no. 9: 1627. https://doi.org/10.3390/ani10091627
APA StyleKottferová, J., Skurková, L., Mesarčová, L., Lešková, L., Demeová, A., & Jakuba, T. (2020). Friendship or Competition? Symmetry in Social Play within the Two Packs of German Shepherd Puppies. Animals, 10(9), 1627. https://doi.org/10.3390/ani10091627