Differences in Tibia Shape in Organically Reared Chicken Lines Measured by Means of Geometric Morphometrics


 ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
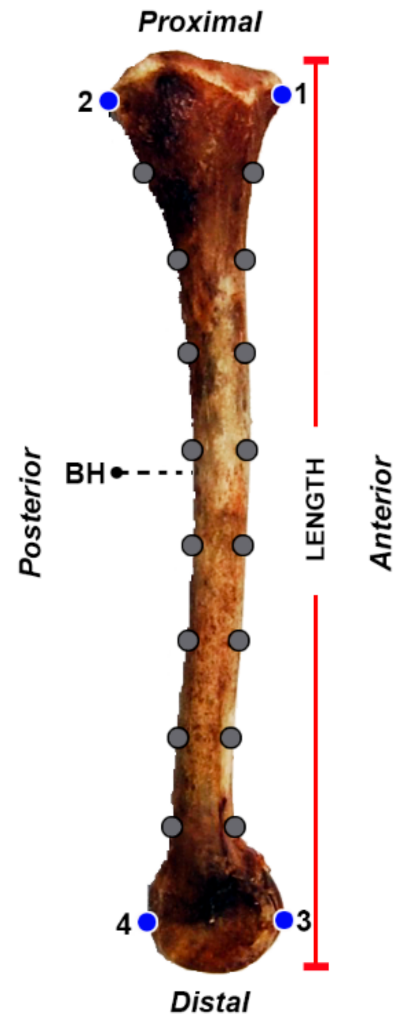
2. Materials and Methods
3. Results and Discussion
3.1. General Results
3.2. Shape Analysis
4. Conclusions
- Tibia shape could represent a reference/starting point for studies on chicken walking behavior. Statistical analysis evidenced a clear and straightforward correlation between the curvature of this bone and the tendency of the animal to spend more time in static activities, such as resting and roosting;
- Degree of curvature of the tibia is positively related to carcass weight and to growth rate. Fast-growing genotypes showed a more pronounced curvature of the tibia;
- Ranger Gold, Rowan Ranger, and RedJA, previously classified as walking poultry lines [20], showed a less curved tibia, similar to that of the reference line (the slow-growing Naked Neck).
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kestin, S.C.; Gordon, S.; Su, G.; Sørensen, P. Relationships in broiler chickens between lameness, liveweight, growth rate and age. Vet. Rec. 2001, 148, 195–197. [Google Scholar] [CrossRef]
- Harash, G.; Richardson, K.C.; Alshamy, Z.; Hünigen, H.; Hafez, H.M.; Plendl, J.; Al Masri, S. Basic morphometry, microcomputed tomography and mechanical evaluation of the tibiotarsal bone of a dual-purpose and a broiler chicken line. PLoS ONE 2020, 15, e0230070. [Google Scholar] [CrossRef]
- Shim, M.Y.; Karnuah, A.B.; Mitchell, A.D.; Anthony, N.B.; Pesti, G.M.; Aggrey, S.E. The effects of growth rate on leg morphology and tibia breaking strength, mineral density, mineral content, and bone ash in broilers. Poult. Sci. 2012, 91, 1790–1795. [Google Scholar] [CrossRef]
- Tickle, P.C.; Paxton, H.; Rankin, J.W.; Hutchinson, J.R.; Codd, J.R. Anatomical and biomechanical traits of broiler chickens across ontogeny. Part I. Anatomy of the musculoskeletal respiratory apparatus and changes in organ size. PeerJ 2014, 2, e432. [Google Scholar] [CrossRef]
- Huang, S.; Kong, A.; Cao, Q.; Tong, Z.; Wang, X. The role of blood vessels in broiler chickens with tibial dyschondroplasia. Poult. Sci. 2019, 98, 6527–6532. [Google Scholar] [CrossRef]
- Kestin, S.C.; Knowles, T.G.; Tinch, A.F.; Gregory, N.G. The prevalence of leg weakness in broiler chickens and its relationship with genotype. Vet. Rec. 1992, 131, 190–194. [Google Scholar] [CrossRef]
- Julian, R.J. Rapid Growth Problems: Ascites and Skeletal Deformities in Broilers. Poult. Sci. 1998, 77, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Duff, S.R.I. Diseases of the musculoskeletal system. In Poultry Diseases; Jordan, F.T.W., Ed.; Balliere Tindall: London, UK, 1990; pp. 254–283. [Google Scholar]
- Bradshaw, R.H.; Kırkden, R.D.; Broom, R.H. A review of the aetiology and pathology of leg weakness in broilers in relation to welfare. Avian Poult. Biol. Rev. 2002, 13, 45–103. [Google Scholar] [CrossRef]
- Corr, S.A.; Mccorquodale, C.C.; Gentle, M.J. Gait Analysis of Poultry. Res. Vet. Sci. 1998, 65, 233–238. [Google Scholar] [CrossRef]
- Guo, Y.; Tang, H.; Wang, X.; Li, W.; Wang, Y.; Yan, F.; Kang, X.; Li, H.R. Clinical assessment of growth performance, bone morphometry, bone quality, and serum indicators in broilers affected by valgus-varus deformity. Poult. Sci. 2019, 98, 4433–4440. [Google Scholar] [CrossRef]
- Hocking, P.M.; Sandercock, D.A.; Wilson, S.; Fleming, R.H. Quantifying genetic (co)variation and effects of genetic selection on tibial bone morphology and quality in 37 lines of broiler, layer and traditional chickens. Br. Poult. Sci. 2009, 50, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.J.; Mills, C.P. Observations on leg deformity in broilers with particular reference to the intertarsal joint. Avian Pathol. 1981, 10, 407–431. [Google Scholar] [CrossRef]
- Vestergaard, K.S.; Sanotra, G.S. Relationships between leg disorders and changes in the behaviour of broiler chickens. Vet. Rec. 1999, 144, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Sherlock, L.; Demmers, T.G.M.; Goodship, A.E.; Mccarthy, I.D.; Wathes, C.M. The relationship between physical activity and leg health in the broiler chicken. Br. Poult. Sci. 2010, 51, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J.; Marcus, L.F. Geometric morphometrics: Reply to M. Corti. Trends Ecol. Evol. 1993, 8, 339. [Google Scholar] [CrossRef]
- Marcus, L.F.; Corti, M.; Loy, A.; Naylor, G.J.P.; Slice, D.E. Advances in Morphometrics; Plenum Press: New York, NY, USA, 1996. [Google Scholar]
- Zelditch, M.L.; Swiderki, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer; Elsevier/Academic Press: San Diego, CA, USA, 2004. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Cartoni Mancinelli, A.; Mattioli, S.; Dal Bosco, A.; Aliberti, A.; Guarino Amato, M.; Castellini, C. Performance, behavior, and welfare status of six different organically reared poultry genotypes. Animals 2020, 10, 550. [Google Scholar] [CrossRef]
- Louton, H.; Keppler, C.; Erhard, M.; van Tuijl, O.; Bachmeier, J.; Damme, K.; Reese, S.; Rauch, E. Animal-based welfare indicators of 4 slow-growing broiler genotypes for the approval in an animal welfare label program. Poult. Sci. 2019, 98, 2326–2337. [Google Scholar] [CrossRef]
- Leterier, C.; Nys, Y. Clinical and anatomical differences in varus and valgus deformities of chick limbs suggest different aetio-pathogenesis. Avian Pathol. 1992, 21, 429–442. [Google Scholar] [CrossRef]
- Rohlf, F.J. TpsDig Version 2.10, Digitalized Landmarks and Outlines; Department of Ecology and Evolution, State University of New York: Stony Book, NY, USA, 2006. [Google Scholar]
- Perez, S.I.; Bernal, V.; Gonzalez, P.N. Differences between sliding semilandmark methods in geometric morphometrics, with an application to human craniofacial and dental variation. J. Anat. 2006, 208, 769–784. [Google Scholar] [CrossRef]
- Pulcini, D.; Russo, T.; Reale, P.; Massa-Gallucci, A.; Brennan, G.; Cataudella, S. Rainbow trout (Oncorhynchus mykiss, Walbaum 1792) develop a more robust body shape under organic rearing. Aquac. Res. 2014, 45, 397–409. [Google Scholar] [CrossRef]
- Gunz, P.; Mitteroecker, P.; Bookstein, F.L. Semilandmarks in three dimensions. In Modern Morphometrics in Physical Anthropology; Slice, D.E., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2005; pp. 73–98. [Google Scholar]
- Gunz, P.; Harvati, K. The Neanderthal ‘chignon’: Variation, integration, and homology. J. Hum. Evol. 2007, 52, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J.; Slice, D.E. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Weiss, S.M.; Kulikowski, C.A. Computer Systems That Learn; Morgan Kaufmann: San Mateo, CA, USA, 1991. [Google Scholar]
- Bookstein, F.L.; Gunz, P.; Mitteroecker, P.; Prossinger, H.; Schaefer, K.; Seidler, H. Cranial integration in Homo: Singular warps analysis of the midsagittal plane in ontogeny and evolution. J. Hum. Evol. 2003, 44, 167–187. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Corti, M. The use of two-block partial least squares to study covariation in shape. Syst. Biol. 2000, 49, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Fadda, C.; Corti, M. Three-dimensional geometric morphometrics of Arvicanthis: Implications for systematics and taxonomy. J. Zool. Syst. Evol. Res. 2001, 39, 235–245. [Google Scholar] [CrossRef]
- Aydin, A. Leg Weaknesses and Lameness Assessment Methods in Broiler Chickens. AAHDS 2018, 1, AAHDS.MS.ID.000506. [Google Scholar] [CrossRef]
- Corr, S.A.; Gentle, M.J.; Mccorquodale, D.; Bennet, D. The effect of morphology on walking ability in the modern broiler: A gait analysis study. Anim. Welf. 2003, 12, 159–171. [Google Scholar]
- Gonzálezcerón, F.; Rekaya, R.; Anthony, N.B.; Aggrey, S.E. Genetic analysis of leg problems and growth in a random mating broiler population. Poult. Sci. 2015, 94, 162–168. [Google Scholar] [CrossRef]
- Kjaer, J.B.; Su, G.; Nielsen, B.L.; Sorensen, P. Foot pad dermatitis and hock burn in broiler chickens and degree of inheritance. Poult. Sci. 2006, 85, 1342–1348. [Google Scholar] [CrossRef]
- Paxton, H.; Tickle, P.J.; Rankin, J.W.; Codd, J.R.; Hutchinson, J.R. Anatomical and biomechanical traits of broiler chickens across ontogeny. Part II. Body segment inertial properties and muscle architecture of the pelvic limb. PeerJ 2014, 2, e473. [Google Scholar] [CrossRef]
- Charuta, A.; Dzierzęcka, M.; Majchrzak, T.; Czerwiński, E.; Cooper, R.G. Computer-generated radiological imagery of the structure of the spongius substance in the postnatal development of the tibiotarsal bones of the Peking domestic duck (Anas platyrhynchos var. domestica). Poult. Sci. 2011, 90, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Kuei, S.C.; Amiel, D.; Gomez, M.A.; Hayes, W.C.; White, F.C.; Akeson, W.H. The effect of prolonged physical training on the properties of long bone: A study of Wolff’s Law. J. Bone Jt. Surg. Am. 1981, 63, 780–787. [Google Scholar] [CrossRef]
- Chen, J.H.; Liu, C.; You, L.; Simmons, C.A. Boning up on Wolff’s Law: Mechanical regulation of the cells that make and maintain bone. J. Biomech. 2010, 43, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Yair, R.; Cahaner, A.; Uni, Z.; Shahar, R. Maternal and genetic effects on broiler bone properties during incubation period. Poult. Sci. 2017, 96, 2301–2311. [Google Scholar] [CrossRef]
- Williams, B.; Solomon, S.; Waddington, D.; Thorp, B.; Farquharson, C. Skeletal development in the meat-type chicken. Br. Poult Sci. 2000, 41, 141–149. [Google Scholar] [CrossRef]
- Williams, B.; Waddington, D.; Murray, D.H.; Farquharson, C. Bone strength during growth: Influence of growth rate on cortical porosity and mineralization. Calcif. Tissue Int. 2004, 74, 236–245. [Google Scholar] [CrossRef]
- Rayner, A.C.; Newberry, R.C.; Vas, J.; Mullan, S. Slow‑growing broilers are healthier and express more behavioural indicators of positive welfare. Sci. Rep. 2020, 10, 15151. [Google Scholar] [CrossRef]
- Cook, M.E. Skeletal deformities and their causes: Introduction. Poult. Sci. 2000, 79, 982–984. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Line | Acronym | Walking Behavior | Growth Rate |
|---|---|---|---|
| Aviagen Ranger Classic | RC | NW | Fast |
| Aviagen Ranger Gold | RG | W | Medium |
| Aviagen Rowan Ranger | RR | W | Slow |
| Hubbard CY Gen5xJA87 | CY | NW | Medium/Fast |
| Hubbard M22xJA87 | M | NW | Medium/Fast |
| Hubbard RedJA | C | W | Slow |
| Hubbard RedJA Naked Neck | NN | W | Slow |
| Acronym | Carcass Weight (g) | Tibia Length (cm) | BH (cm) |
|---|---|---|---|
| RC | 3616.0 ± 54.4 b | 13.4 ± 0.4 b | 1.3 ± 0.2 ab |
| CY | 3487.7 ± 117.6 b | 13.9 ± 1.3 b | 1.3 ± 0.2 ab |
| M | 3213.3 ± 121.2 c | 14.2 ± 0.8 b | 1.4 ± 0.1 a |
| RG | 3179.3 ± 71.7 c | 14.1 ± 0.5 b | 1.2 ± 0.1 bc |
| RR | 2859.33 ± 115.0 c | 13.6 ± 0.8 b | 1.1 ± 0.2 c |
| C | 2930.7 ± 77.2 c | 14.2 ± 0.6 b | 1.1 ± 0.2 c |
| NN | 2431.6 ±61.7 a | 12.0 ± 0.6 a | 0.9 ± 0.1 d |
| True | Allocated to | Total | % | |
|---|---|---|---|---|
| W | NW | |||
| W | 37 | 4 | 41 | 90.2 |
| NW | 10 | 50 | 60 | 83.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulcini, D.; Meo Zilio, D.; Cenci, F.; Castellini, C.; Guarino Amato, M. Differences in Tibia Shape in Organically Reared Chicken Lines Measured by Means of Geometric Morphometrics. Animals 2021, 11, 101. https://doi.org/10.3390/ani11010101
Pulcini D, Meo Zilio D, Cenci F, Castellini C, Guarino Amato M. Differences in Tibia Shape in Organically Reared Chicken Lines Measured by Means of Geometric Morphometrics. Animals. 2021; 11(1):101. https://doi.org/10.3390/ani11010101
Chicago/Turabian StylePulcini, Domitilla, David Meo Zilio, Francesco Cenci, Cesare Castellini, and Monica Guarino Amato. 2021. "Differences in Tibia Shape in Organically Reared Chicken Lines Measured by Means of Geometric Morphometrics" Animals 11, no. 1: 101. https://doi.org/10.3390/ani11010101
APA StylePulcini, D., Meo Zilio, D., Cenci, F., Castellini, C., & Guarino Amato, M. (2021). Differences in Tibia Shape in Organically Reared Chicken Lines Measured by Means of Geometric Morphometrics. Animals, 11(1), 101. https://doi.org/10.3390/ani11010101