1. Introduction
Oxidative damage is a critical problem in the poultry industry. It occurs when exogenously- and/or endogenously-produced Reactive Oxygen Species (ROS) exceed the antioxidant capacity of cells and extracellular spaces, causing the disruption of the normal cellular function by influencing the gene expression and signal transduction [
1]. World poultry populations are mainly located in tropical and subtropical regions, where heat stress is a main concern, as it negatively influences the antioxidant status, which is reflected by the increased serum lipid peroxidation and reduced plasma concentrations of antioxidants [
2]. These stress factors affect the health status of poultry and the safety of poultry products, while they adversely influence the intestinal oxidative status and disrupt the normal function of enterocytes, causing an abnormality in nutrient absorption and diseases [
3]. As a consequence of suppressed nutrient absorption, the production indexes (such as feed efficiency and survival rate) are impaired, leading to heavy economic losses to the poultry industry [
4].
Tight Junctions (TJs) are intercellular junctional complexes that maintain epithelial cells adherent to each other and guarantee the paracellular transportation of nutrients, forming a barrier between the lumen and host to prevent bacterial translocation [
3]. The disruption of TJ impaired the normal intestinal function causing leaky gut that compromises the absorption of luminal substances into the bloodstream and could lead to chronic inflammation with impairment of animal health and growth performances [
5]. Furthermore, the increased intestinal permeability could induce bacterial translocation, and a systemic bacterial infection might also occur [
6]. Therefore, protecting the intestinal tract from excessive oxidative damage and maintaining the integrity of the TJ could be major factors that positively influence the performance of birds in the intensive poultry industry [
4,
7].
Agricultural by-products, such as Wheat Bran (WB), contain Non-Starch Polysaccharides (NSPs) that act as anti-nutritional compounds in monogastric animals, which tend to inhibit digestibility and promote pathogen proliferation in the gastrointestinal tract, and eventually lead to gut inflammation and worsen the performance of the animals [
8,
9]. Solid-State Fermentation (SSF) could represent a valuable method to use agricultural by-products as substrates for NSP-degrading microorganisms, converting them into alternative feed ingredients [
10,
11]. In addition, filamentous fungi were reported as being a suitable inoculant for SSF due to their ability to withstand low humidity environments [
11,
12]. WB, a solid-state that is fermented by the
Trichoderma sp., has been shown to exhibit antioxidant properties and to improve the nutrition value of WB, which could potentially be a low-cost feedstuff candidate [
13,
14]. Solid-state WB fermented by
Antrodia cinnamomea was found to be suitable for producing bioactive compounds, such as phenolics, triterpenoids, and polysaccharides, as well as for growth promotion and its antioxidative and anti-inflammatory effects in broiler feeding trials [
11,
15]. According to these studies, the use of fungal SSF in agricultural by-products is viable for producing functional feedstuffs that contain bioactive compounds.
The
Laetiporus sp. is a fungal species with medical properties, and it was traditionally used by Europeans to cure gastric cancer, rheumatism, pyretic diseases, and coughs [
16]. In our previous studies, we showed that
L. sulphureus fermented WB and potentially enhanced the growth performance of broilers by modifying their intestinal microflora and their immune status [
17]. Furthermore, submerged mycelial cultures of
L. sulphureus produce functional polysaccharides [
18,
19], as well as mycophenolic acids [
20], and are able to dampen the excessive immune response of the selected cells without causing cytotoxicity [
21,
22]. Petrović et al. [
23] reported that the aqueous extracts of wild
L. sulphureus exert 1, 1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging activity, which correlates with its total phenolic content. Our previous study also indicated the potential of
L. sulphureus to produce various bioactive compounds, such as crude phenolics, crude triterpenoids, polysaccharides, and ergosterol. The same study also showed that ethanol extracts of
L. sulphureus fermented WB significantly attenuated the DNA damage induced by 2,2′-Azobis (2-amidinopropane) dihydrochloride (AAPH) in peripheral blood mononuclear cells of chickens, while exerting an in vitro antioxidant effect, including DPPH radical scavenging activity and reducing power [
24]. However, the effects of
L. sulphureus on the antioxidative status and TJ modulation in the gastrointestinal tracts of broilers has rarely been studied.
In order to evaluate the in vivo antioxidant properties of
L. sulphureus solid-state fermented product (FL) in broiler chickens, nuclear factor erythroid 2–related factor 2
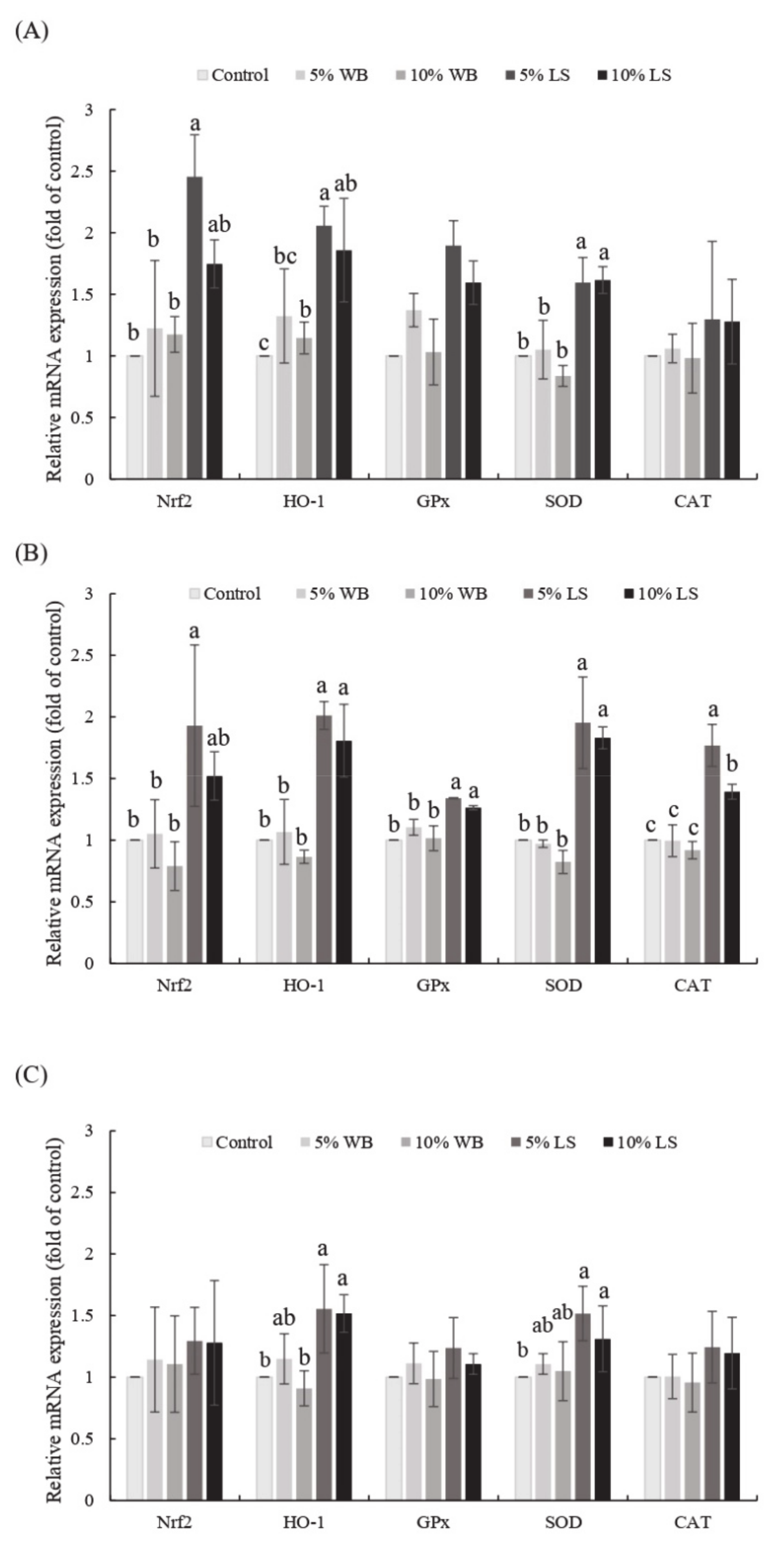
(Nrf2) and its downstream pathway was evaluated. Nrf2 is a redox-sensitive transcription factor that could be triggered by ROS, leading to the production of detoxification and antioxidant enzymes such as heme oxygenase-1 (HO-1), superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) [
2]. Therefore, this study was performed in order to evaluate if the in vitro antioxidant activity of FL could be used in vivo to improve the antioxidant status of broiler chickens and further investigated the effect of FL on broilers and the interaction between the antioxidation capacity and the expression of intestinal TJ mRNA.
4. Discussion
In order to evaluate the in vivo antioxidant properties of FL in broiler chickens, we focused on
Nrf2 and its downstream pathway. In this study, the expression of
Nrf2,
HO-1, and
SOD mRNAs in FL supplemented groups were up-regulated in the liver and jejunum, compared to those in the control group, which indicates the antioxidant role of FL in broilers. Nrf2 is a redox-sensitive transcription factor that is localized in the cytoplasm and binds with Kelch-like ECH-associated protein 1 (Keap 1), an actin-binding protein, under normal conditions [
32]. Upon activation by ROS, Nrf2 dissociates from Keap 1, interacts with the antioxidant response elements, and regulates the expression of downstream antioxidant genes to activate the antioxidant and detoxifying effects [
2]. The induction of
HO-1 is an important cellular process for dealing with oxidative stress by degrading the intracellular levels of pro-oxidant heme and by producing biliverdin (a precursor of bilirubin) [
33]. The elevated antioxidative status of FL supplemented groups could be due to the bioactive phenolic compounds within FL. The phenolic compounds derived from fungus have been reported to activate the Nrf2-Keap1 pathway in a suppressed
Nrf2 diabetic rat model [
34]. Furthermore,
A. cinnamomea, a brown-rot fungus that exerted medicinal effects similar to those of
L. sulphureus, reported the enhanced expression of
Nrf2,
HO-1, and
SOD mRNAs in the liver of 35-day-old chickens fed upon its fermented feedstuff, as well as a further increase in SOD activity in the serum [
11,
15]. Therefore, it can be validated that FL, as a medicinal fungus fermented products, may have also improved the antioxidative status of broilers via a similar mechanism as reported by above mentioned previous studies.
Non-enzymatic and enzymatic antioxidant systems are two strategies that are used in cells to inhibit the potential ROS toxicity. Enzymatic antioxidants are endogenously synthesized and regulated, which is a crucial indicator for evaluating the oxidative status of animal tissues [
11,
15]. SOD is an endogenous antioxidant enzyme that catalyzes the dismutation of O
2- to H
2O
2 and O
2. In our study, the elevated SOD activity in the 5% FL supplemented group indicated an increase in the extracellular antioxidant enzymatic activity. Similarly, Lin et al. [
35] observed that the dietary supplementation of mulberry leaves improved the serum SOD levels and showed that these outcomes were due to the presence of abundant phenolic compounds in the tested product, which was in common with FL that consisted of phenolic compounds that exerted antioxidant ability. The development of oxidative injury could be indicated by the serum concentration of MDA because it is one of the end products of lipid peroxidation [
36]. In this study, the FL-supplemented groups had a higher SOD activity and a lower MDA concentration. The elevated expression of serum SOD and
HO-1 mRNA enhanced the capacity of broilers to catalyze the harmful radicals and encounter potential oxidative damage, which further led to the reduction of MDA in the serum, which represents a total decline in the lipid oxidation in animals. Likewise, Lee et al. [
11] reported that the
A. cinnamomea-fermented product supported the antioxidant status of broiler chickens by improving the SOD activities. Furthermore, the enzyme powder fermented by
Trichoderma pseudokoningii was also found to exert an antioxidant effect on broiler chickens, which increased the serum SOD activities and reduced the MDA concentration [
14]. Interestingly, FL seemed to have a less-pronounced effect on the ileum, which might be due to the direct absorbance of a simple-structured polyphenol in the jejunum, while the remaining complicated polyphenols are more likely to be fermented in the hindgut by cecal microbiota and to exert their bio-functional effects [
37].
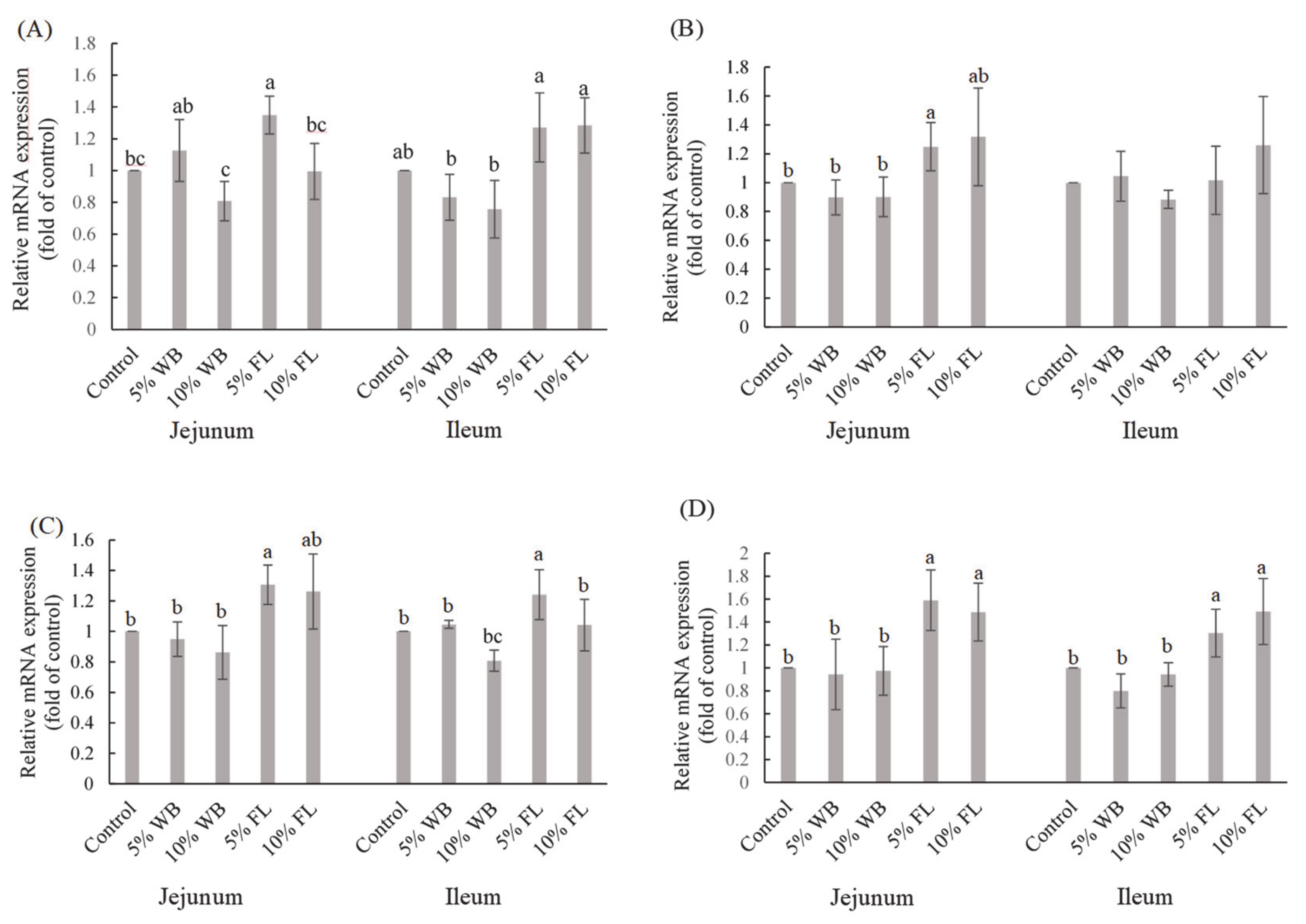
According to the results of this study, the increased mRNA expression of transmembrane proteins claudin-1 (CLDN-1), occludin (OCLN), and mRNA of peripheral membrane protein zonula occluden-1 (ZO-1) may be caused by the enhancement of the antioxidative status in the jejunum of the FL-supplemented groups. By improving the function of TJ and eliminating the deleterious effects of oxidative stress, the increase in
MUC-2 mRNA expression could lead to the improved health condition of intestinal goblet cells. The integrity of epithelial cells and the normal function of the intestine in broiler chickens is maintained by the TJs, which consist of several crucial elements, including occludin, claudins, and ZOs.
OCLN translates to occludin that forms the TJs, and its sealing property is involved in the hurdle functions of the epithelial barrier [
38]. Claudins (translated from
CLDN) are another family of integral membrane proteins that collaborate with occludin to maintain the integrity of the TJs [
5]. ZO-1 and ZO-2 bind directly to the COOH terminus of the intracellular domain of occludin, which contributes to the normal structure of the epithelial barrier function [
6]. In addition, the
MUC-2 gene is expressed by goblet cells, which produce a mucus layer that helps in blocking pathogens invasion along with TJs [
39]. The TJs could be disturbed when animals encounter heat stress and oxidative damage due to excessive free radicals, which could lead to leaky gut and onset of inflammatory response with poor animal health and impaired growth performances as a consequence. Moreover,
MUC-2 deficiency in mice causes spontaneous inflammation and allows the colonization of unusual commensal bacteria [
40]. The interaction of the antioxidant status and TJ integrity has also been proven in several studies by using various animals and cell models. Chen et al. [
9] reported that riboflavin deprivation decreased the antioxidant enzyme activities, such as SOD, GPx, and glutathione reductase activities in young grass carp, which further reduced the expression of TJ mRNA,
OCLN, and
ZO-1. Zhao et al. [
41] reported that the
MUC-2 function could be interrupted by bisphenol A, which induces mitochondrial dysfunction and oxidative stress. Exogenous antioxidants, including polyphenols and polysaccharides, help to scavenge the excessive free radicals and prevent oxidative damage to the intestinal cells and proteins [
42]. In an H
2O
2-induced oxidative damaged Caco-2 cell model, a phytogenic called red-osier dogwood, which contained abundant phenolic compounds, was introduced to enhance the expression of
HO-1,
SOD,
GPx genes, and the Nrf2 protein, while improving the cell-integrity by increasing the expression of ZO-1 and claudin-3 that were damaged by H
2O
2. In addition, Sun et al. [
43] demonstrated that the essential oil cinnamaldehyde exerted an antioxidant activity and elevated the protein expression of claudin-4, occluding, and ZO-1. Chitosan oligosaccharide, an antioxidant and an immunomodulatory substance, has been proven to improve
OCLN mRNA expression in the jejunum of broilers, while an increase in antioxidant enzymes and reduced levels of proinflammatory cytokine IL-6 were also observed [
44]. Quercetin, an antioxidant flavonoid, was demonstrated to increase
MUC-2 gene expression in human intestinal goblet cell-like LS174T [
45]. Therefore, the improvement of the
MUC-2 status in broilers supplemented with FL could result in the elevation of TJ integrity.
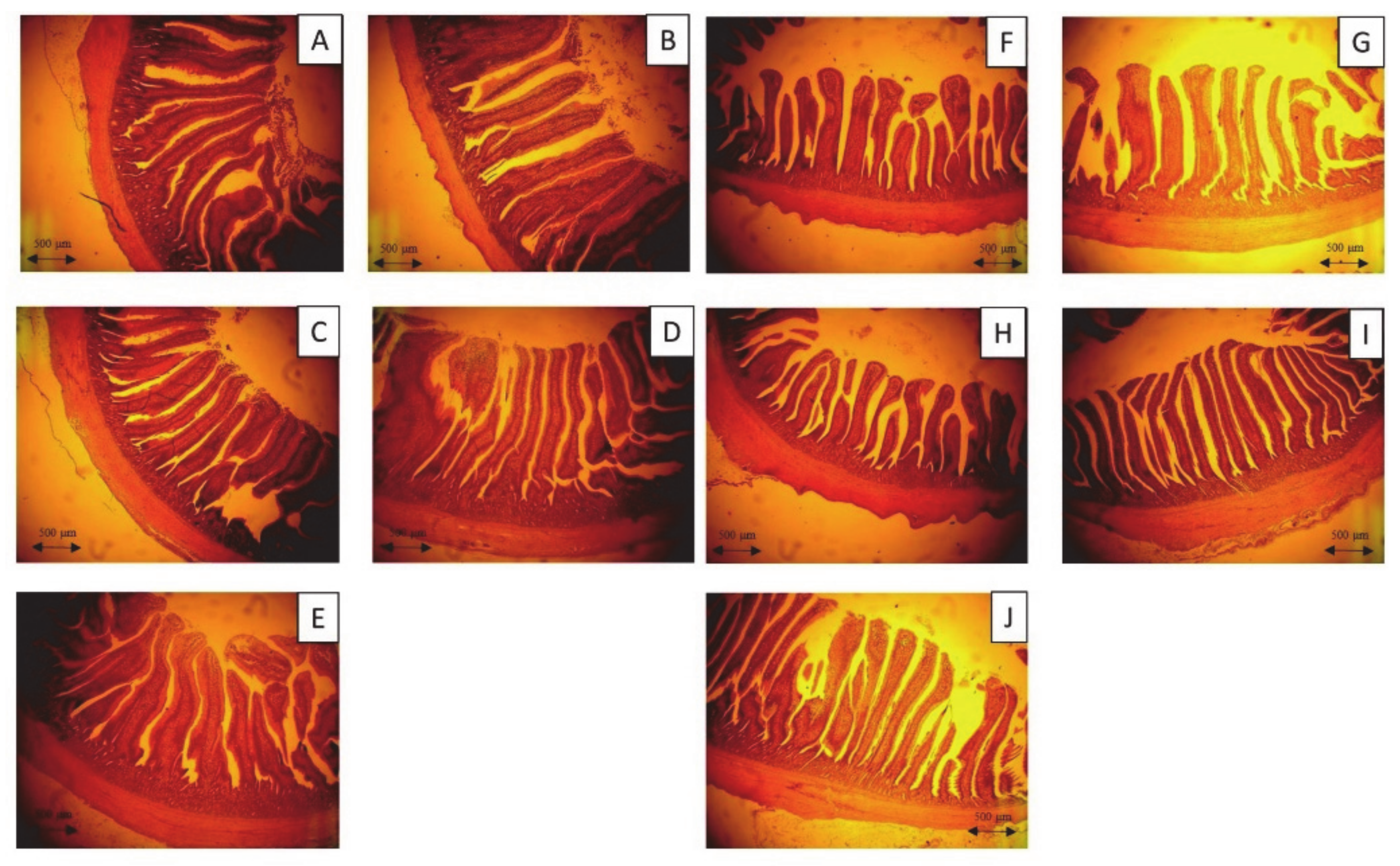
The villus height and crypt depth, with respect to the morphologies of the jejunum and ileum of broilers, are always used as an indicator of intestinal health. The villus height represents nutrient absorption efficiency, and a low crypt depth is favored, due to the reduced intestinal cell turnover rate, to save energy and achieve a better growth performance [
46,
47]. In our previous study, lignocellulose enzymes (laccase and xylanase), which are capable of degrading the deleterious effects of NSP-rich WB, were produced during
L. sulphureus fermentation [
17]. The improved morphologies of the jejunum and ileum in broilers could be due to the lignocellulose in FL. Similar results were also demonstrated by Chu et al. [
13], who observed a positive impact of
Trichoderma fermented wheat bran on the intestinal morphology of broilers, while Lin et al. [
14] showed that the
T. pseudokoningii fermented enzyme powder that contains NSPase improved the intestinal morphology of broilers. Furthermore, FL was found to be rich in phenolic compounds and capable of exerting an antioxidant effect, thus preventing oxidative stress. In the lumen of the intestine, stressors, including pathogens and pro-oxidants, can cause dynamic changes in the intestinal mucosa due to the close proximity of the mucosal surface and the intestinal contents [
1]. According to the outcomes of TJ and intestinal morphology, the antioxidant effect of FL could potentially protect the intestinal mucosa, as well as the integrity of epithelial cells, from oxidative damage. Similar results were shown by Viveros et al. [
48], who suggested that the dietary grape pomace concentrate (60 g/kg in feed) that are rich in polyphenols could improve the ratio of villus height and crypt depth (villus height: crypt depth) and increased the feed efficiency of broilers. Likewise, Lai et al. [
8] reported that fermentation of soybean hulls containing phenolic-rich residues of
Pleurotus eryngii stalk increased the villus height/crypt depth in the ileum. In this study, FL improved the villi:crypt ratio in the ileum compared to the control and WB groups, while higher villi and less shallow crypts were discovered in both jejunum and ileum of 5% FL group compared to the WB and control groups. These improvements in intestinal morphology could be possibly caused by the antioxidant effect along with the potential existence of NSP enzymes.






{kind=link}
{kind=link}
{kind=link}