
Altered Hypoxia-Induced and Heat Shock Protein Immunostaining in Secondary Hair Follicles Associated with Changes in Altitude and Temperature in Tibetan Cashmere Goats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tibet Cashmere Goats in Rikaze Steppe and Huan County Steppe
2.2. Skin Sample Collection
2.3. Wax Section and Ultrathin Section Analysis
2.4. Immunohistochemical and Immunofluorescence Analysis
2.5. Measurements and Statistical Analysis
3. Results
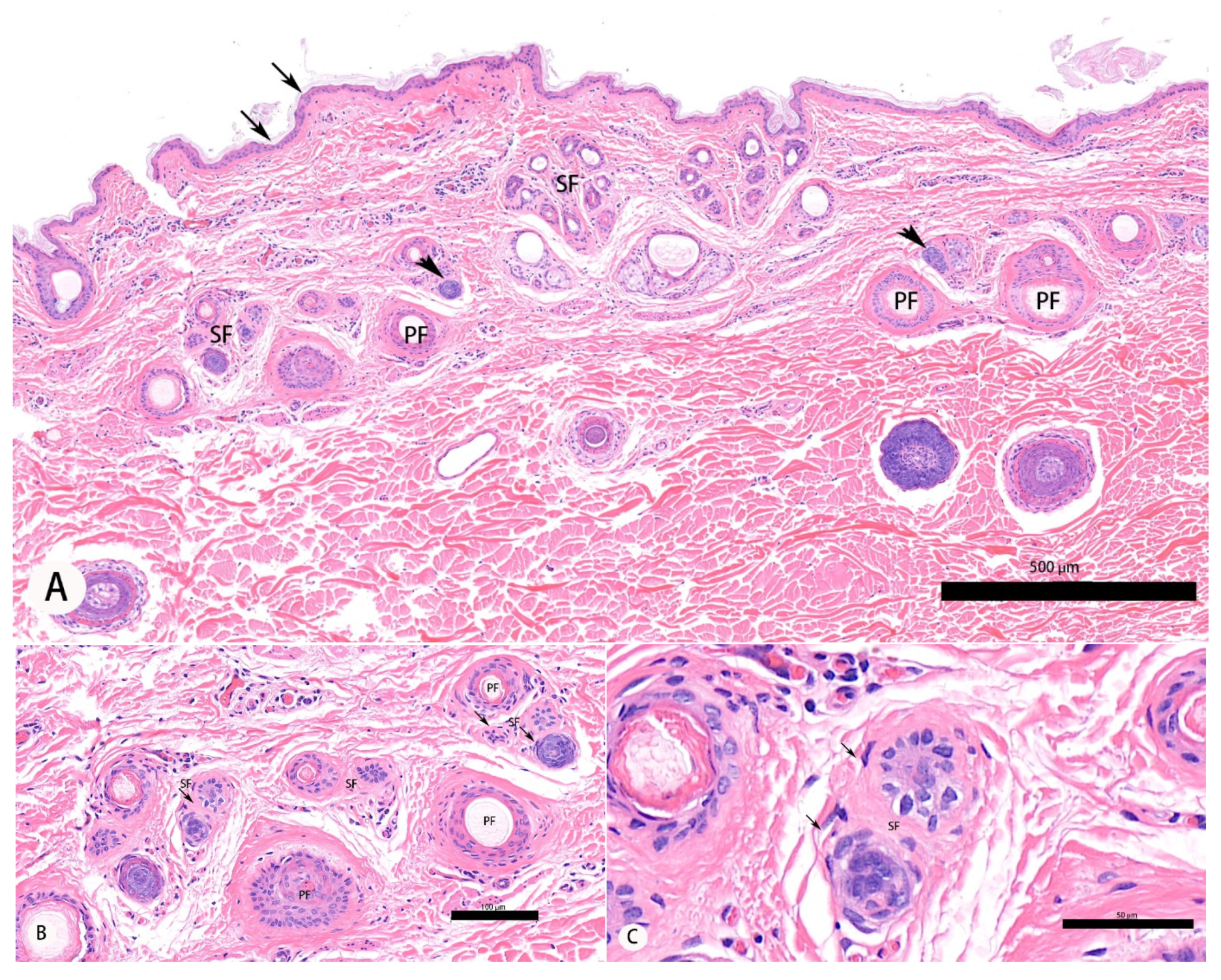
3.1. Morphological Analysis of Secondary Hair Follicle and Histomorphological Evaluations
3.2. Immunohistochemical Detection of HIF-1a, HIF-2a, HIF-3a, HSP27, and HOXC13 in Secondary Hair Follicles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Bai, J.Y.; Zhang, Q.; Li, J.Q.; Dao, E.J.; Jia, X. Estimates of genetic parameters and genetic trends for production traits of Inner Mongolian White Cashmere goat. Asian-Australas J. Anim. Sci. 2006, 19, 13–18. [Google Scholar] [CrossRef]
- FAOSTAT. Final 2012 Data and Preliminary 2013 Data for Live Animals. Available online: http://faostat.fao.org/site/573/DesktopDefault.Aspx?PageID=573#ancor (accessed on 1 January 2015).
- Wang, Y.; Wang, J.; Zi, X.D.; Huatai, C.R.; Ouyang, X.; Liu, L.S. Genetic diversity of Tibetan goats of Plateau type using microsatellite markers. Arch. Anim. Breed. 2011, 54, 188–197. [Google Scholar] [CrossRef]
- Zhang, C.J.; Wang, Y.; Lu, F.S. Study on electrocardiogram of Qinghai native goat. Progress Vet. Med. 2004, 4, 021. [Google Scholar]
- Simonson, T.S.; Yang, Y.Z.; Hu, C.D.; Yun, H.X.; Qin, G.; Witherspoon, D.J.; Bai, Z.Z.; Lorenzo, F.R.; Xing, J.C.; Jorde, L.B.; et al. Genetic evidence for high altitude adaptation in Tibet. Science 2010, 329, 72–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, R.L.; Cai, Q.; Shen, Y.Y.; San, A.; Ma, L.; Zhang, Y.; Yi, X.; Chen, Y.; Yang, L.F.; Huang, Y.; et al. Draft genome sequence of the Tibetan antelope. Nat. Commun. 2013, 4, 1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Yu, S.; Hu, J.; Cui, Y.; Liu, P. Changes in the anatomic microscopic structure and the expression of HIF-1_ and VEGF of the yak heart with aging and hypoxia. PLoS ONE 2016, 11, e0149947. [Google Scholar]
- Wang, Y.; Luo, M.R.; Lin, Y.Q.; Zhu, J.J. Comparative analysis of tissue expression and methylation reveal the crucial hypoxia genes in hypoxia resistant animals. Can. J. Anim. Sci. 2018, 98, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Alexander, A.; Jensen, R. Gross cardiac changes in cattle with high mountain (brisket) disease and in experimental cattle maintained at high altitudes. Am. J. Vet. Res. 1959, 20, 680–689. [Google Scholar]
- Recavarren, S.; Arias-Stella, J. Right ventricular hypertrophy in people born and living at high altitudes. Br. Heart J. 1964, 26, 806–812. [Google Scholar] [CrossRef] [Green Version]
- Ge, R.L.; Mo, V.Y.; Januzzi, J.L.; Jin, G.; Yang, Y.; Han, S.; Wood, M.J.; Levine, B.D. B-type natriuretic peptide, vascular endothelial growth factor, endothelin-1, and nitric oxide synthase in chronic mountain sickness. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1427–H1433. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.; Shimoda, L.A.; Iyer, N.V.; Huso, D.L.; Sun, X.; McWilliams, R.; Beaty, T.; Sham, J.S.K.; Wiener, C.M.; Sylvester, J.T.; et al. Impaired physiological responses to chronic hypoxia in mice partially deficient for hypoxia-inducible factor 1alpha. J. Clin. Investig. 1999, 103, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Tipoe, G.L.; Fung, M.L. Expression of HIF-1_lpha, VEGF and VEGF receptors in the carotid body of chronically hypoxic rat. Respir. Physiol. Neurobiol. 2003, 138, 143–154. [Google Scholar] [CrossRef]
- Maynard, M.A.; Evans, A.J.; Shi, W.; Kim, W.Y.; Liu, F.F.; Ohh, M. Dominant-negative HIF-3 alpha 4 suppresses VHL-null renal cell carcinoma progression. Cell Cycle 2007, 6, 2810–2816. [Google Scholar] [CrossRef] [Green Version]
- He, Y.Y.; Munday, S.J.; Perrott, M.; Wang, G.; Liu, X. Association of age with the expression of hypoxia-inducible factors HIF-1a, HIF-2a, HIF-3a and VEGF in lung and heart of Tibetan Sheep. Animals 2019, 9, 673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Fan, Z.; Han, E.; Hou, R.; Zhang, L.; Galaverni, M.; Huang, J.; Liu, H.; Silva, P.; Li, P.; et al. Hypoxia adaptations in the grey wolf (Canis lupus chanco) from Qinghai-Tibet Plateau. PLoS Genet. 2014, 10, e1004466. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.F.; Wang, X.R.; Yang, Y.W.; Lin, H. Hypoxia upregulates hypoxia inducible factor (HIF)-1a expression in lung epithelial cells: Characterization and comparison with HIF-1a. Cell Res. 2006, 16, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, J.N.; Lambert, H.; Hickey, E.; Weber, L.A.; Landry, J. Modulation of cellular thermoresistance and actin filaments stability accompanies phosphorylation-induced changes in the oligomeric structure of heat shock protein 27. Mol. Cell. Biol. 1995, 15, 05–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adly, M.A.; Assaf, H.A.; Hussein, M.R. Expression of the heat shock protein-27 in the adult human scalp skin and hair follicle: Hair cycle-dependent changes. J. Am. Acad. Dermatol. 2006, 54, 811–817. [Google Scholar] [CrossRef]
- Jantschitsch, C.; Kindas-Mugge, I.; Metze, D.; Amann, G.; Micksche, M.; Trautinger, F. Expression of the small heat shock protein HSP 27 in developing human skin. Br. J. Dermatol. 1998, 139, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Du, P.; Wang, H.F.; He, Y.Y. HOCX13 and HSP27 expression in skin and the periodic growth of secondary fibre hair follicles from Longdong cashmere goats raised in different production system. Anat. Rec. 2018, 301, 742–752. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.H.; Yan, Z.W.; Zhang, W.G.; Li, J.Q. HOXC13 and the development of hair follicle. Hereditas 2010, 32, 656–662. [Google Scholar]
- Wu, J.H.; Zhang, Y.J.; Zhang, J.X.; Chang, Z.L.; Li, J.Q.; Yan, Z.W.; Zhang, W.G. HOXC13/b-catenin correlation with hair follicle activity in cashmere goat. J. Integr. Agric. 2012, 11, 1159–1166. [Google Scholar]
- Wu, J.H.; Zhang, W.G.; Li, J.Q.; Yin, J.; Zhang, Y.J. HOXC13 expression pattern in cashmere goat skin during hair follicle development. Agric. Sci. China 2009, 8, 491–496. [Google Scholar] [CrossRef]
- He, Y.Y.; Luo, Y.Z.; Cheng, L.X.; Wang, J.Q.; Liu, X.; Li, S.B.; Cui, Y. Determination of secondary hair follicle characteristics, density, activity, and Hoxc13 expression pattern of Hexi cashmere goats breed. Anat. Record 2015, 298, 1796–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panaretto, B.A. Effect of light and cyclic activity of wool hair follicle and possible relationships to changes in the pelage of other mammals. In Physiological and Environmental Limitations to Wool Growth; Black, J.L., Reis, P.J., Eds.; University of New England Publishing Unit: Armidale, Australia, 1979; pp. 327–336. [Google Scholar]
- Yusuf, N.; Nasti, T.; Ahmad, I.; Chowdhury, S.; Mohiuddin, H.; Xu, H.; Athar, M.; Timares, L.; Elmets, C. In Vivo Suppression of Heat Shock Protein (HSP)27 and HSP70 Accelerates DMBA-Induced Skin Carcinogenesis by Inducing Antigenic Unresponsiveness to the Initiating Carcinogenic Chemical. J. Immunol. 2015, 194, 4796–4803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, S.D. Heat shock proteins and the skin. Clin. Exp. Dermatol. 2002, 27, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Trautinger, F.; Trautinger, I.; Kindas-Mugge, I.; Metze, D.; Luger, T.A. Human keratinocytes in vivo and in vitro constitutively express the 72-kD heat shock protein. J. Investig. Dermatol. 1993, 101, 334–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, N.; McArdle, A.; Guerin, D.; Tasker, H.; Wareing, P.; Foster, C.S.; Jackson, M.J.; Rhodes, L.E. Hyperthermia to normal human skin in vivo upregulates heat shock proteins 27, 60, 72i and 90. J. Cutan. Pathol. 2000, 27, 176–182. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Liu, X.; De, J.; Kang, S.; Munday, J.S. Altered Hypoxia-Induced and Heat Shock Protein Immunostaining in Secondary Hair Follicles Associated with Changes in Altitude and Temperature in Tibetan Cashmere Goats. Animals 2021, 11, 2798. https://doi.org/10.3390/ani11102798
He Y, Liu X, De J, Kang S, Munday JS. Altered Hypoxia-Induced and Heat Shock Protein Immunostaining in Secondary Hair Follicles Associated with Changes in Altitude and Temperature in Tibetan Cashmere Goats. Animals. 2021; 11(10):2798. https://doi.org/10.3390/ani11102798
Chicago/Turabian StyleHe, Yanyu, Xiu Liu, Jie De, Saihong Kang, and John S. Munday. 2021. "Altered Hypoxia-Induced and Heat Shock Protein Immunostaining in Secondary Hair Follicles Associated with Changes in Altitude and Temperature in Tibetan Cashmere Goats" Animals 11, no. 10: 2798. https://doi.org/10.3390/ani11102798