
Sexual Development in Non-Human Parasitic Apicomplexa: Just Biology or Targets for Control?



Abstract
:Simple Summary
Abstract
1. Introduction
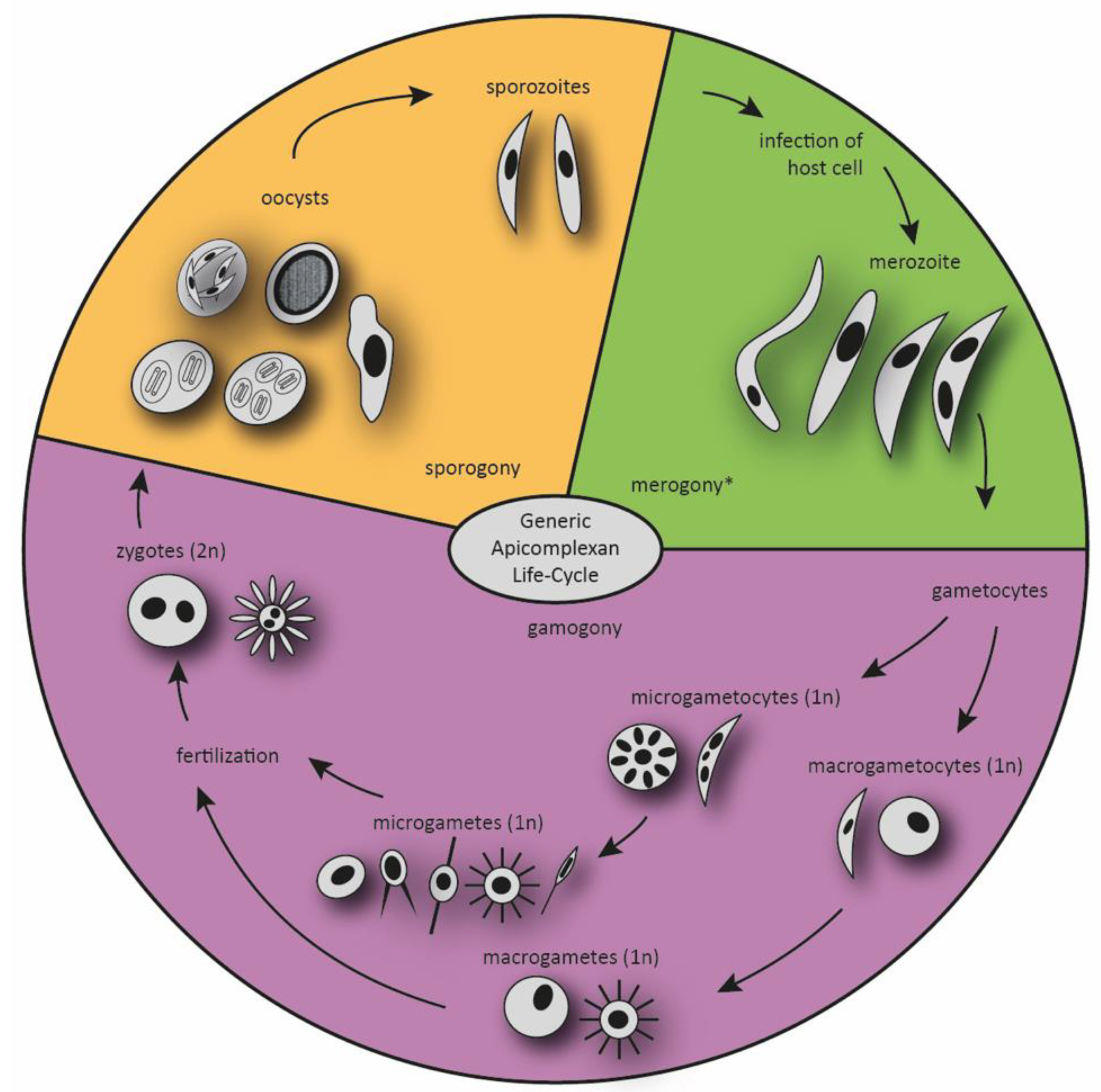
2. Sexual Reproduction in Apicomplexa
2.1. Class Conoidasida
2.1.1. Subclass Gregarinasina
2.1.2. Subclass Cryptogregaria
2.1.3. Subclass Coccidiasina
2.2. Class Aconoidasida
2.2.1. Order Haemosporida
2.2.2. Order Piroplasmida
3. Breaking the Cycle—Targeting Sexual Stages of Apicomplexan Parasites for Intervention
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Schurko, A.M.; Neiman, M.; Logsdon, J.M. Signs of sex: What we know and how we know it. Trends Ecol. Evol. 2009, 24, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Schurko, A.M.; Logsdon, J.M. Using a meiosis detection toolkit to investigate ancient asexual “scandals” and the evolution of sex. BioEssays 2008, 30, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Cavaliersmith, T. Origins of the machinery of recombination and sex. Heredity 2002, 88, 125–141. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Origin of the cell nucleus, mitosis and sex: Roles of intracellular coevolution. Biol. Direct 2010, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleckner, N. Meiosis: How could it work? Proc. Natl. Acad. Sci. USA 1996, 93, 8167–8174. [Google Scholar] [CrossRef] [Green Version]
- Solari, A.J. Primitive forms of meiosis: The possible evolution of meiosis. Biocell 2002, 26, 1–13. [Google Scholar]
- Goodenough, U.; Heitman, J. Origins of eukaryotic sexual reproduction. Cold Spring Harb. Perspect. Biol. 2014, 6, a016154. [Google Scholar] [CrossRef] [Green Version]
- Weedall, G.D.; Hall, N. Sexual reproduction and genetic exchange in parasitic protists. Parasitology 2014, 142, S120–S127. [Google Scholar] [CrossRef] [Green Version]
- Martens, K.; Rossetti, G.; Horne, D.J. How ancient are ancient asexuals? Proc. Biol. Sci. 2003, 270, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, M.; Hülsmann, N. Protists–A textbook example for a paraphyletic taxon. Org. Divers. Evol. 2007, 7, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Taylor, F.J.R.M. The collapse of the two-kingdom system, the rise of protistology and the founding of the International Society for Evolutionary Protistology (ISEP). Int. J. Syst. Evol. Microbiol. 2003, 53, 1707–1714. [Google Scholar] [CrossRef]
- Adl, S.M.; Simpson, A.; Lane, C.E.; Lukes, J.; Bass, D.; Bowser, S.S.; Brown, M.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–514. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W. The sexual side of parasitic protists. Mol. Biochem. Parasitol. 2021, 243, 111371. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Syst. Evol. Microbiol. 2002, 52, 297–354. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.; Akbar, H.; Ahmad, L.; Hassan, M.A.; Ashraf, K.; Saeed, K.; Gharbi, M. A systematic review on modelling approaches for economic losses studies caused by parasites and their associated diseases in cattle. Parasitology 2019, 146, 129–141. [Google Scholar] [CrossRef]
- Tomley, F.M.; Shirley, M.W. Livestock infectious diseases and zoonoses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2637–2642. [Google Scholar] [CrossRef] [Green Version]
- Joachim, A. Vaccination against parasites–status quo and the way forward. Porc. Health Manag. 2016, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Palatnik-De-Sousa, C.B.; Nico, D. The delay in the licensing of protozoal vaccines: A comparative history. Front. Immunol. 2020, 11, 204. [Google Scholar] [CrossRef]
- McAllister, M.M. Successful vaccines for naturally occurring protozoal diseases of animals should guide human vaccine research. A review of protozoal vaccines and their designs. Parasitolgy 2014, 141, 624–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledford, H. Malaria vaccine shows promise—Now come tougher trials. Nat. Cell Biol. 2021, 593, 17. [Google Scholar] [CrossRef]
- Datoo, M.S.; Natama, M.H.; Somé, A.; Traoré, O.; Rouamba, T.; Bellamy, D.; Yameogo, P.; Valia, D.; Tegneri, M.; Ouedraogo, F.; et al. High Efficacy of a Low Dose Candidate Malaria Vaccine, R21 in 1 Adjuvant Matrix-MTM, with seasonal administration to children in Burkina Faso. SSRN Electron. J. 2021, 397, 1809–1818. [Google Scholar] [CrossRef]
- Laurens, M.B. RTS,S/AS01 vaccine (Mosquirix™): An overview. Hum. Vaccines Immunother. 2020, 16, 480–489. [Google Scholar] [CrossRef]
- Smith, T.G.; Walliker, D.; Ranford-Cartwright, L. Sexual differentiation and sex determination in the Apicomplexa. Trends Parasitol. 2002, 18, 315–323. [Google Scholar] [CrossRef]
- Walker, R.; Ferguson, D.; Miller, C.; Smith, N. Sex and Eimeria: A molecular perspective. Parasitology 2013, 140, 1701–1717. [Google Scholar] [CrossRef]
- Ralph, S.A.; Cortés, A. Plasmodium sexual differentiation: How to make a female. Mol. Microbiol. 2019, 112, 1627–1631. [Google Scholar] [CrossRef] [Green Version]
- Josling, G.A.; Llinás, M. Sexual development in Plasmodium parasites: Knowing when it’s time to commit. Nat. Rev. Microbiol. 2015, 13, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Votýpka, J.; Modrý, D.; Oborník, M.; Šlapeta, J.; Lukeš, J. Apicomplexa. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Margulis, L., Melkonian, M., Chapman, D.J., Corliss, J.O., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–58. ISBN 978-3-319-32669-6. [Google Scholar]
- Gubbels, M.-J.; Keroack, C.D.; Dangoudoubiyam, S.; Worliczek, H.L.; Paul, A.S.; Bauwens, C.; Elsworth, B.; Engelberg, K.; Howe, D.K.; Coppens, I.; et al. Fussing about fission: Defining variety among mainstream and exotic apicomplexan cell division modes. Front. Cell. Infect. Microbiol. 2020, 10, 269. [Google Scholar] [CrossRef]
- Guttery, D.; Roques, M.; Holder, A.; Tewari, R. Commit and transmit: Molecular players in Plasmodium sexual development and zygote differentiation. Trends Parasitol. 2015, 31, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Francia, M.; Striepen, B. Cell division in apicomplexan parasites. Nat. Rev. Microbiol. 2014, 12, 125–136. [Google Scholar] [CrossRef]
- Ferguson, D.J.P.; Sahoo, N.; Pinches, R.A.; Bumstead, J.M.; Tomley, F.M.; Gubbels, M.-J. MORN1 has a conserved role in asexual and sexual development across the apicomplexa. Eukaryot. Cell 2008, 7, 698–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Striepen, B.; Jordan, C.N.; Reiff, S.; Van Dooren, G.G. Building the perfect parasite: Cell division in apicomplexa. PLoS Pathog. 2007, 3, e78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, J.; Bunker, B. The life cycle and fitness domain of gregarine (Apicomplexa) parasites. Ecol. Model. 2012, 233, 31–40. [Google Scholar] [CrossRef]
- Lee, J.J.; Leedale, G.F.; Bradbury, P.C. An Illustrated Guide to the Protozoa: Organisms Traditionally Referred to as Protozoa, or Newly Discovered Groups, 2nd ed.; Allen Press Inc.: Lawrence, KS, USA, 2000. [Google Scholar]
- Morrison, D.A. Evolution of the Apicomplexa: Where are we now? Trends Parasitol. 2009, 25, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Chambouvet, A.; Valigurová, A.; Mesquita, L.; Richards, T.A.; Jirků, M. Nematopsis temporariae (Gregarinasina, Apicomplexa, Alveolata) intracellular infectious agent of tadpole livers. Environ. Microbiol. Rep. 2016, 8, 675–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueckert, S.; Betts, E.L.; Tsaousis, A.D. The symbiotic spectrum: Where do the gregarines fit? Trends Parasitol. 2019, 35, 687–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desportes, I.; Schrével, J. Treatise on zoology-anatomy, taxonomy, biology. In The Gregarines (2 vols): The Early Branching Apicomplexa; Brill: Leiden, The Netherlands, 2013; ISBN 978-90-04-25605-7. [Google Scholar]
- Boisard, J.; Florent, I. Why the–omic future of Apicomplexa should include gregarines. Biol. Cell 2020, 112, 173–185. [Google Scholar] [CrossRef]
- Lantova, L.; Volf, P. Mosquito and sand fly gregarines of the genus Ascogregarina and Psychodiella (Apicomplexa: Eugregarinorida, Aseptatorina)—Overview of their taxonomy, life cycle, host specificity and pathogenicity. Infect. Genet. Evol. 2014, 28, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Ryan, U.; Hijjawi, N. New developments in Cryptosporidium research. Int. J. Parasitol. 2015, 45, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Clode, P.L.; Koh, W.H.; Thompson, R.A. Life without a host cell: What is Cryptosporidium? Trends Parasitol. 2015, 31, 614–624. [Google Scholar] [CrossRef]
- Ryan, U.; Paparini, A.; Monis, P.; Hijjawi, N. It’s official—Cryptosporidium is a gregarine: What are the implications for the water industry? Water Res. 2016, 105, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalier-Smith, T. Gregarine site-heterogeneous 18S rDNA trees, revision of gregarine higher classification, and the evolutionary diversification of Sporozoa. Eur. J. Protistol. 2014, 50, 472–495. [Google Scholar] [CrossRef] [PubMed]
- Hijjawi, N.; Meloni, B.; Ryan, U.; Olson, M.; Thompson, R. Successful in vitro cultivation of Cryptosporidium andersoni: Evidence for the existence of novel extracellular stages in the life cycle and implications for the classification of Cryptosporidium. Int. J. Parasitol. 2002, 32, 1719–1726. [Google Scholar] [CrossRef]
- Hijjawi, N. Cryptosporidium: New developments in cell culture. Exp. Parasitol. 2010, 124, 54–60. [Google Scholar] [CrossRef]
- Sinden, R.E. Targeting the parasite to suppress malaria transmission. Adv. Parasitol. 2017, 97, 147–185. [Google Scholar] [CrossRef]
- Fédry, J.; Liu, Y.; Pehau-Arnaudet, G.; Pei, J.; Li, W.; Tortorici, M.A.; Traincard, F.; Meola, A.; Bricogne, G.; Grishin, N.V.; et al. The ancient gamete fusogen HAP2 is a eukaryotic class II fusion protein. Cell 2017, 168, 904–915.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tandel, J.; English, E.D.; Sateriale, A.; Gullicksrud, J.A.; Beiting, D.P.; Sullivan, M.C.; Pinkston, B.; Striepen, B. Life cycle progression and sexual development of the apicomplexan parasite Cryptosporidium parvum. Nat. Microbiol. 2019, 4, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Lippuner, C.; Ramakrishnan, C.; Basso, W.; Schmid, M.W.; Okoniewski, M.; Smith, N.; Hässig, M.; Deplazes, P.; Hehl, A.B. RNA-Seq analysis during the life cycle of Cryptosporidium parvum reveals significant differential gene expression between proliferating stages in the intestine and infectious sporozoites. Int. J. Parasitol. 2018, 48, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Templeton, T.J.; Lancto, C.A.; Vigdorovich, V.; Liu, C.; London, N.R.; Hadsall, K.Z.; Abrahamsen, M.S. The Cryptosporidium oocyst wall protein is a member of a multigene family and has a homolog in Toxoplasma. Infect. Immun. 2004, 72, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Feix, A.S.; Cruz-Bustos, T.; Ruttkowski, B.; Joachim, A. Characterization of Cystoisospora suis sexual stages in vitro. Parasites Vectors 2020, 13, 143. [Google Scholar] [CrossRef]
- Walker, A.R.; Sharman, A.P.; Miller, C.M.; Lippuner, C.; Okoniewski, M.; Eichenberger, R.M.; Ramakrishnan, C.; Brossier, F.; Deplazes, P.; Hehl, A.B.; et al. RNA Seq analysis of the Eimeria tenella gametocyte transcriptome reveals clues about the molecular basis for sexual reproduction and oocyst biogenesis. BMC Genom. 2015, 16, 94. [Google Scholar] [CrossRef] [Green Version]
- Matthews, H.; Duffy, C.W.; Merrick, C.J. Checks and balances? DNA replication and the cell cycle in Plasmodium. Parasites Vectors 2018, 11, 216. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, C.; Maier, S.; Walker, R.A.; Rehrauer, H.; Joekel, D.E.; Winiger, R.; Basso, W.U.; Grigg, M.E.; Hehl, A.B.; Deplazes, P.; et al. An experimental genetically attenuated live vaccine to prevent transmission of Toxoplasma gondii by cats. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Tomasina, R.; Francia, M.E. The Structural and molecular underpinnings of gametogenesis in Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2020, 10, 608291. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.; Dubremetz, J.-F. The ultrastructure of Toxoplasma gondii. In Toxoplasma Gondii, 3rd ed.; Weis, L., Kim, K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 21–61. [Google Scholar] [CrossRef]
- Di Genova, B.M.; Knoll, L.J. Comparisons of the sexual cycles for the coccidian parasites Eimeria and Toxoplasma. Front. Cell. Infect. Microbiol. 2020, 10, 776. [Google Scholar] [CrossRef]
- Laxer, M.A.; Healey, M.C.; Youssef, N.N. Production of monoclonal antibodies specific for Eimeria tenella microgametocytes. J. Parasitol. 1987, 73, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Madden, A.P.; Vetterling, J.M. Scanning electron microscopy of Eimeria tenella microgametogenesis and fertilization. J. Parasitol. 1977, 63, 607–610. [Google Scholar] [CrossRef]
- Di Genova, B.M.; Wilson, S.K.; Dubey, J.P.; Knoll, L.J. Intestinal delta-6-desaturase activity determines host range for Toxoplasma sexual reproduction. PLoS Biol. 2019, 17, e3000364. [Google Scholar] [CrossRef]
- Pinto, J.D.S. Parasitic castration in males of Sardinia pilchardus (Walb.) due to testicular infestation by the coccidian Eimeria sardinae (Thélohan). In Revista da Faculdade de Ciencias Naturalis; Lisbon Universidade: Lisbon, Portugal, 1956; Volume 5, pp. 209–224. [Google Scholar]
- Sivajothi, S.; Reddy, B.S.; Rayulu, V.C. Study on impression smears of hepatic coccidiosis in rabbits. J. Parasit. Dis. 2014, 40, 906–909. [Google Scholar] [CrossRef] [Green Version]
- Ammar, S.I.; Watson, A.; Craig, L.E.; Cope, E.R.; Schaefer, J.J.; Mulliniks, J.T.; Gerhold, R.W. Eimeria gilruthi—Associated abomasitis in a group of ewes. J. Vet. Diagn. Investig. 2019, 31, 128–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, J.P. Coccidiosis in the Gallbladder of a Goat. Proc. Helminthol. Soc. Wash. 1986, 53, 227–281. [Google Scholar]
- Dubey, J.P. Coccidiosis in livestock, poultry, companion animals, and humans. In Coccidiosis in Livestock, Poultry, Companion Animals, and Humans, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Dubey, J.; Lindsay, D.; Lappin, M.R. Toxoplasmosis and other intestinal coccidial infections in cats and dogs. Vet. Clin. N. Am. Small Anim. Pract. 2009, 39, 1009–1034. [Google Scholar] [CrossRef]
- Lappin, M.R. Chapter 82-Isosporiasis. In Canine and Feline Infectious Diseases; Sykes, J., Ed.; W.B. Saunders: Saint Louis, MO, USA, 2014; pp. 793–796. ISBN 978-1-4377-0795-3. [Google Scholar]
- Lindsay, D.S.; Dubey, J. Neosporosis, toxoplasmosis, and sarcocystosis in ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 205–222. [Google Scholar] [CrossRef]
- Reid, A.; Vermont, S.J.; Cotton, J.; Harris, D.; Hill-Cawthorne, G.; Könen-Waisman, S.; Latham, S.M.; Mourier, T.; Norton, R.; Quail, M.A.; et al. Comparative genomics of the apicomplexan parasites Toxoplasma gondii and Neospora caninum: Coccidia differing in host range and transmission strategy. PLoS Pathog. 2012, 8, e1002567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangoura, B.; Bardsley, K.D. Ruminant Coccidiosis. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Hou, Z.; Liu, D.; Jianping, T.; Wang, L.; Xu, J.; Tao, J. Comparative transcriptome analysis of second-and third-generation merozoites of Eimeria necatrix. Parasites Vectors 2017, 10, 388. [Google Scholar] [CrossRef]
- Mehlhorn, H. Elektronenmikroskopische Untersuchungen an Entwicklungsstadien von Eimeria maxima aus dem Haushuhn. Zeitschrift Parasitenkd. 1972, 40, 243–260. [Google Scholar] [CrossRef]
- Scholtyseck, E.; Mehlhorn, H.; Hammond, D.M. Electron microscope studies of microgametogenesis in coccidia and related groups. Zeitschrift Parasitenkd 1972, 38, 95–131. [Google Scholar] [CrossRef]
- Elwasila, M. The fine structure of an early stage in the process of fertilization of Eimeria maxima (Apicomplexa, Eimeriina). Zeitschrift Parasitenkd. Parasitol. Res. 1982, 69, 135–138. [Google Scholar] [CrossRef]
- Su, S.; Hou, Z.; Liu, D.; Jia, C.; Wang, L.; Xu, J.; Tao, J. Comparative transcriptome analysis of Eimeria necatrix third-generation merozoites and gametocytes reveals genes involved in sexual differentiation and gametocyte development. Vet. Parasitol. 2018, 252, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasites Vectors 2020, 13, 588. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J. The history and life cycle of Toxoplasma gondii. In Toxoplasma Gondii: The Model Apicomplexan-Perspectives and Methods, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 1–17. ISBN 9780123964816. [Google Scholar]
- Shrestha, A.; Abd-Elfattah, A.; Freudenschuss, B.; Hinney, B.; Palmieri, N.; Ruttkowski, B.; Joachim, A. Cystoisospora suis—A model of mammalian cystoisosporosis. Front. Vet. Sci. 2015, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, N.; Shrestha, A.; Ruttkowski, B.; Beck, T.; Vogl, C.; Tomley, F.; Blake, D.P.; Joachim, A. The genome of the protozoan parasite Cystoisospora suis and a reverse vaccinology approach to identify vaccine candidates. Int. J. Parasitol. 2017, 47, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Fritz, H.M.; Bowyer, P.W.; Bogyo, M.; Conrad, P.A.; Boothroyd, J.C. Proteomic Analysis of fractionated Toxoplasma oocysts reveals clues to their environmental resistance. PLoS ONE 2012, 7, e29955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Possenti, A.; Fratini, F.; Fantozzi, L.; Pozio, E.; Dubey, J.P.; Ponzi, M.; Pizzi, E.; Spano, F. Global proteomic analysis of the oocyst/sporozoite of Toxoplasma gondii reveals commitment to a host-independent lifestyle. BMC Genom. 2013, 14, 183. [Google Scholar] [CrossRef] [Green Version]
- Fritz, H.M.; Buchholz, K.R.; Chen, X.; Durbin-Johnson, B.; Rocke, D.; Conrad, P.A.; Boothroyd, J.C. Transcriptomic Analysis of Toxoplasma development reveals many novel functions and structures specific to sporozoites and oocysts. PLoS ONE 2012, 7, e29998. [Google Scholar] [CrossRef]
- Ramakrishnan, C.; Smith, N.C. Recent achievements and doors opened for coccidian parasite research and development through transcriptomics of enteric sexual stages. Mol. Biochem. Parasitol. 2021, 243, 111373. [Google Scholar] [CrossRef]
- Feix, A.S.; Cruz-Bustos, T.; Ruttkowski, B.; Mötz, M.; Rümenapf, T.; Joachim, A. Progression of asexual to sexual stages of Cystoisospora suis in a host cell-free environment as a model for Coccidia. Parasitology 2021, 148, 1–25. [Google Scholar] [CrossRef]
- Barta, J.R.; Ogedengbe, J.D.; Martin, D.S.; Smith, T.G. Phylogenetic position of the Adeleorinid Coccidia (Myzozoa, Apicomplexa, Coccidia, Eucoccidiorida, Adeleorina) inferred using 18S rDNA sequences. J. Eukaryot. Microbiol. 2012, 59, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Maia, J.P.; Carranza, S.; Harris, D.J. Comments on the systematic revision of adeleid haemogregarines: Are More data needed? J. Parasitol. 2016, 102, 549–552. [Google Scholar] [CrossRef]
- O’Donoghue, P. Haemoprotozoa: Making biological sense of molecular phylogenies. Int. J. Parasitol. Parasites Wildl. 2017, 6, 241–256. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Abdel-Ghaffar, F.; Dkhil, M.A.; Abdel-Gaber, R. Haemogregarines and criteria for identification. Animals 2021, 11, 170. [Google Scholar] [CrossRef]
- O’Dwyer, L.H.; Moço, T.C.; Da Silva, R.J. Description of the gamonts of a small species of Hepatozoon sp. (Apicomplexa, Hepatozoidae) found in Crotalus durissus terrificus (Serpentes, Viperidae). Parasitol. Res. 2003, 92, 110–112. [Google Scholar] [CrossRef]
- Laakkonen, J.; Sukura, A.K.K.; Oksanen, A.; Henttonen, H.; Soveri, T. Haemogregarines of the genus Hepatozoon (Apicomplexa: Adeleina) in rodents from northern Europe. Folia Parasitol. 2001, 48, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilgūnas, M.; Chagas, C.R.F.; Bukauskaitė, D.; Bernotienė, R.; Iezhova, T.; Valkiūnas, G. The life-cycle of the avian haemosporidian parasite Haemoproteus majoris, with emphasis on the exoerythrocytic and sporogonic development. Parasites Vectors 2019, 12, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkiunas, G. Avian Malaria Parasites and Other Haemosporidia, 1st ed.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar] [CrossRef]
- Fletcher, K.; Träff, J.; Gustafsson, L. Importance of infection of haemosporidia blood parasites during different life history stages for long-term reproductive fitness of collared flycatchers. J. Avian Biol. 2019, 50, 50. [Google Scholar] [CrossRef]
- Levin, I.I.; Parker, P.G. Chapter 47-Haemosporidian parasites: Impacts on avian hosts. In Zoo and Wild Animal Medicine Current Therapy; Miller, R.E., Lamberski, N., Calle, P.P., Eds.; W.B. Saunders: Saint Louis, MO, USA, 2012; pp. 356–363. ISBN 978-1-4377-1986-4. [Google Scholar]
- Ngotho, P.; Soares, A.B.; Hentzschel, F.; Achcar, F.; Bertuccini, L.; Marti, M. Revisiting gametocyte biology in malaria parasites. FEMS Microbiol. Rev. 2019, 43, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, F.; Matuschewski, K. Plasmodium Sporozoite Biology. Cold Spring Harb. Perspect. Med. 2017, 7, a025478. [Google Scholar] [CrossRef] [Green Version]
- Guttery, D.S.; Holder, A.A.; Tewari, R. Sexual development in Plasmodium: Lessons from functional analyses. PLoS Pathog. 2012, 8, e1002404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, J.; Oberstaller, J.; Adams, J. Targeting gametocytes of the malaria parasite Plasmodium falciparum in a functional genomics era: Next steps. Pathogens 2021, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Vega-Rodríguez, J.; Jacobs-Lorena, M. The Plasmodium bottleneck: Malaria parasite losses in the mosquito vector. Memórias Instituto Oswaldo Cruz 2014, 109, 644–661. [Google Scholar] [CrossRef]
- Painter, H.J.; Carrasquilla, M.; Llinás, M. Capturing in vivo RNA transcriptional dynamics from the malaria parasite Plasmodium falciparum. Genome Res. 2017, 27, 1074–1086. [Google Scholar] [CrossRef] [Green Version]
- Neveu, G.; Beri, D.; Kafsack, B.F. Metabolic regulation of sexual commitment in Plasmodium falciparum. Curr. Opin. Microbiol. 2020, 58, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Poran, A.; Nötzel, C.; Aly, O.; Trinchant, N.M.; Harris, C.T.; Guzman, M.L.; Hassane, D.C.; Elemento, O.; Kafsack, B.F. Single-cell RNA sequencing reveals a signature of sexual commitment in malaria parasites. Nature 2017, 551, 95–99. [Google Scholar] [CrossRef]
- Josling, G.A.; Russell, T.J.; Venezia, J.; Orchard, L.; Van Biljon, R.; Painter, H.J.; Llinás, M. Dissecting the role of PfAP2-G in malaria gametocytogenesis. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kafsack, B.F.; Rovira-Graells, N.; Clark, T.; Bancells, C.; Crowley, V.; Campino, S.G.; Williams, A.; Drought, L.G.; Kwiatkowski, D.; Baker, D.; et al. A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 2014, 507, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Morillo, R.C.; Tong, X.; Xie, W.; Lenz, T.; Batugedara, G.; Tabassum, N.; Orchard, L.M.; Daher, W.; Patel, D.J.; Noble, W.S.; et al. Homeodomain protein 1 is an essential regulator of gene expression during sexual differentiation of malaria parasites. bioRxiv 2020. bioRxiv:2020.10.26.352583. [Google Scholar] [CrossRef]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The complexity of piroplasms life cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Mehlhorn, H.; Schein, E. The piroplasms: Life cycle and sexual stages. Adv. Parasitol. 1985, 23, 37–103. [Google Scholar] [CrossRef]
- Rudzinska, M.A.; Spielman, A.; Riek, R.F.; Lewengrub, S.J.; Plesman, J. Intraerythrocytic ’gametocytes’ of Babesia microti and their maturation in ticks. Can. J. Zool. 1979, 57, 424–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beugnet, F.; Moreau, Y. Babesiosis. Rev. Sci. Tech. 2015, 34, 627–639. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Schein, E. The piroplasms: “A long story in short” or “Robert Koch has seen it”. Eur. J. Protistol. 1993, 29, 279–293. [Google Scholar] [CrossRef]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia life cycle—When phylogeny meets biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Rudzinska, M.A.; Spielman, A.; Lewengrub, S.; Trager, W.; Piesman, J. Sexuality in piroplasms as revealed by electron microscopy in Babesia microti. Proc. Natl. Acad. Sci. USA 1983, 80, 2966–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Pieszko, M.; Weir, W.; Goodhead, I.; Kinnaird, J.; Shiels, B. ApiAP2 Factors as candidate regulators of stochastic commitment to merozoite production in Theileria annulata. PLoS Negl. Trop. Dis. 2015, 9, e0003933. [Google Scholar] [CrossRef] [Green Version]
- Alzan, H.F.; Knowles, D.P.; Suarez, C.E. Comparative Bioinformatics analysis of transcription factor genes indicates conservation of key regulatory domains among Babesia bovis, Babesia microti, and Theileria equi. PLoS Negl. Trop. Dis. 2016, 10, e0004983. [Google Scholar] [CrossRef] [PubMed]
- Alzan, H.; Cooke, B.M.; Suarez, C.E. Transgenic Babesia bovis lacking 6-Cys sexual-stage genes as the foundation for non-transmissible live vaccines against bovine babesiosis. Ticks Tick-Borne Dis. 2019, 10, 722–728. [Google Scholar] [CrossRef]
- Schneider, P.; Reece, S.E. The private life of malaria parasites: Strategies for sexual reproduction. Mol. Biochem. Parasitol. 2021, 244, 111375. [Google Scholar] [CrossRef]
- Coutinho-Abreu, I.V.; Ramalho-Ortigao, M. Transmission blocking vaccines to control insect-borne diseases: A review. Memórias Instituto Oswaldo Cruz 2010, 105, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmenarejo, G.; Lozano, S.; González-Cortés, C.; Calvo, D.; Sanchez-Garcia, J.; Matilla, J.-L.P.; Leroy, D.; Rodrigues, J. Predicting transmission blocking potential of anti-malarial compounds in the mosquito feeding assay using Plasmodium falciparum male gamete inhibition Assay. Sci. Rep. 2018, 8, 7764. [Google Scholar] [CrossRef] [PubMed]
- Birkholtz, L.-M.; Coetzer, T.L.; Mancama, D.; Leroy, D.; Alano, P. Discovering new transmission-blocking antimalarial compounds: Challenges and opportunities. Trends Parasitol. 2016, 32, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Wallach, M.G.; Mencher, D.; Yarus, S.; Pillemer, G.; Halabi, A.; Pugatsch, T. Eimeria maxima: Identification of gametocyte protein antigens. Exp. Parasitol. 1989, 68, 49–56. [Google Scholar] [CrossRef]
- Wallach, M.; Halabi, A.; Pillemer, G.; Sar-Shalom, O.; Mencher, D.; Gilad, M.; Bendheim, U.; Danforth, H.D.; Augustine, P.C. Maternal immunization with gametocyte antigens as a means of providing protective immunity against Eimeria maxima in chickens. Infect. Immun. 1992, 60, 2036–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krücken, J.; Hosse, R.J.; Mouafo, A.N.; Entzeroth, R.; Bierbaum, S.; Marinovski, P.; Hain, K.; Greif, G.; Wunderlich, F. Excystation of Eimeria tenella sporozoites impaired by antibody recognizing gametocyte/oocyst antigens GAM22 and GAM56. Eukaryot. Cell 2008, 7, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Liu, Q.-R.; Han, J.-P.; Qian, W.-F.; Liu, Q. Anti-recombinant gametocyte 56 protein IgY protected chickens from homologous coccidian infection. J. Integr. Agric. 2012, 11, 1721–1728. [Google Scholar] [CrossRef]
- Wallach, M.G.; Ashash, U.; Michael, A.; Smith, N. Field Application of a subunit vaccine against an enteric protozoan disease. PLoS ONE 2008, 3, e3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fried, M.; Mencher, D.; Sar-Shalom, O.; Wallach, M. Developmental gene expression of a 230-kilodalton macrogamete-specific protein of the avian coccidial parasite, Eimeria maxima. Mol. Biochem. Parasitol. 1992, 51, 251–262. [Google Scholar] [CrossRef]
- Rafiqi, S.I.; Garg, R.; Ram, H.; Reena, K.K.; Asari, M.; Kumari, P.; Kundave, V.R.; Singh, M.; Banerjee, P.S. Immunoprophylactic evaluation of recombinant gametocyte 22 antigen of Eimeria tenella in broiler chickens. Parasitol. Res. 2019, 118, 945–953. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Tao, J. Efficacy of a DNA vaccine carrying Eimeria maxima Gam56 antigen gene against coccidiosis in chickens. Korean J. Parasitol. 2013, 51, 147–154. [Google Scholar] [CrossRef]
- Wiedmer, S.; Alnassan, A.A.; Volke, B.; Thabet, A.; Daugschies, A.; Lendner, M.; Kurth, M. Passive immunization with Eimeria tenella gametocyte antigen 56 (EtGAM56) specific antibodies and active immunization trial with the epitope containing peptide. Vet. Parasitol. 2017, 247, 100–107. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.-W.; Park, M.S.; Cha, S.-R.; Lillehoj, E.P.; Subramanian, B.M.; Sriraman, R.; Srinivasan, V. Eimeria maxima recombinant Gam82 gametocyte antigen vaccine protects against coccidiosis and augments humoral and cell-mediated immunity. Vaccine 2010, 28, 2980–2985. [Google Scholar] [CrossRef]
- Sharman, P.A.; Smith, N.C.; Wallach, M.G.; Katrib, M. Chasing the golden egg: Vaccination Against poultry coccidiosis. Parasite Immunol. 2010, 32, 590–598. [Google Scholar] [CrossRef]
- Venkatas, J.; Adeleke, M.A. A review of Eimeria antigen identification for the development of novel anticoccidial vaccines. Parasitol. Res. 2019, 118, 1701–1710. [Google Scholar] [CrossRef]
- Larsen, N.C.; Rasmussen, K.R.; Healey, M.C. Production and partial characterization of monoclonal antibodies specific for the gamonts of Eimeria tenella. J. Parasitol. 1991, 77, 1012–1015. [Google Scholar] [CrossRef]
- Suarez, C.; Bishop, R.; Alzan, H.; Poole, W.; Cooke, B. Advances in the application of genetic manipulation methods to apicomplexan parasites. Int. J. Parasitol. 2017, 47, 701–710. [Google Scholar] [CrossRef]
- Wong, J.L.; Johnson, M.A. Is HAP2-GCS1 an ancestral gamete fusogen? Trends Cell Biol. 2010, 20, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; von Besser, K.; Zhou, Q.; Smith, E.; Aux, G.; Patton, D.; Levin, J.; Preuss, D. Arabidopsis hapless mutations define essential gametophytic functions. Genetics 2004, 168, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Pradel, G. Proteins of the malaria parasite sexual stages: Expression, function and potential for transmission blocking strategies. Parasitology 2007, 134, 1911–1929. [Google Scholar] [CrossRef] [PubMed]
- Chichester, J.A.; Green, B.J.; Jones, R.M.; Shoji, Y.; Miura, K.; Long, C.A.; Lee, C.K.; Ockenhouse, C.F.; Morin, M.J.; Streatfield, S.J.; et al. Safety and immunogenicity of a plant-produced Pfs25 virus-like particle as a transmission blocking vaccine against malaria: A Phase 1 dose-escalation study in healthy adults. Vaccine 2018, 36, 5865–5871. [Google Scholar] [CrossRef] [PubMed]
- Duffy, P.E.; Gorres, J.P. Malaria vaccines since 2000: Progress, priorities, products. Vaccine 2020, 5, 1–9. [Google Scholar] [CrossRef]
- Coelho, C.H.; Tang, W.K.; Burkhardt, M.; Galson, J.D.; Muratova, O.; Salinas, N.D.; e Silva, T.L.A.; Reiter, K.; MacDonald, N.J.; Nguyen, V.; et al. A human monoclonal antibody blocks malaria transmission and defines a highly conserved neutralizing epitope on gametes. Nat. Commun. 2021, 12, 1750. [Google Scholar] [CrossRef] [PubMed]
- Blagborough, A.M.; Sinden, R.E. Plasmodium berghei HAP2 induces strong malaria transmission-blocking immunity in vivo and in vitro. Vaccine 2009, 27, 5187–5194. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhao, Y.; Liu, F.; Ye, B.; Zhao, Z.; Thongpoon, S.; Roobsoong, W.; Sattabongkot, J.; Cui, L.; Fan, Q.; et al. Evaluation of Plasmodium vivax HAP2 as a transmission-blocking vaccine candidate. Vaccine 2020, 38, 2841–2848. [Google Scholar] [CrossRef]
- Angrisano, F.; Sala, K.A.; Da, D.F.; Liu, Y.; Pei, J.; Grishin, N.V.; Snell, W.J.; Blagborough, A.M. Targeting the conserved fusion loop of HAP2 inhibits the transmission of Plasmodium berghei and falciparum. Cell Rep. 2017, 21, 2868–2878. [Google Scholar] [CrossRef] [Green Version]
- Talman, A.M.; Prieto, J.H.; Marques, S.R.L.O.; Ubaida-Mohien, C.; Lawniczak, M.; Wass, M.N.; Xu, T.; Frank, R.; Ecker, A.; Stanway, R.S.; et al. Proteomic analysis of the Plasmodium male gamete reveals the key role for glycolysis in flagellar motility. Malar. J. 2014, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Straschil, U.; Talman, A.M.; Ferguson, D.J.P.; Bunting, K.A.; Xu, Z.; Bailes, E.; Sinden, R.E.; Holder, A.A.; Smith, E.F.; Coates, J.C.; et al. The armadillo repeat protein PF16 Is Essential for flagellar structure and function in Plasmodium male gametes. PLoS ONE 2010, 5, e12901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeeshan, M.; Ferguson, D.J.; Abel, S.; Burrrell, A.; Rea, E.; Brady, D.; Daniel, E.; Delves, M.; Vaughan, S.; Holder, A.A.; et al. Kinesin-8B controls basal body function and flagellum formation and is key to malaria transmission. Life Sci. Alliance 2019, 2, e201900488. [Google Scholar] [CrossRef] [Green Version]
- Hussein, H.E.; Bastos, R.G.; Schneider, D.A.; Johnson, W.C.; Adham, F.K.; Davis, W.C.; Laughery, J.M.; Herndon, D.R.; Alzan, H.F.; Ueti, M.W.; et al. The Babesia bovis hap2 gene is not required for blood stage replication, but expressed upon in vitro sexual stage induction. PLoS Negl. Trop. Dis. 2017, 11, e0005965. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Nuez, M.; Hernández-Silva, D.J.; Castañeda-Ortiz, E.J.; Paredes-Martínez, M.E.; Rocha-Martínez, M.K.; Alvarez-Sánchez, M.E.; Mercado-Curiel, R.F.; Aguilar-Tipacamu, G.; Mosqueda, J. Hap2, a novel gene in Babesia bigemina is expressed in tick stages, and specific antibodies block zygote formation. Parasites Vectors 2017, 10, 568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehlhorn, H. Vaccination against protozoa. In Encyclopedia of Parasitology; Springer: Berlin, Germany, 2008; p. 1545. [Google Scholar]


{kind=link}
| Class. | Conoidasida | Aconoidasida | ||||||
|---|---|---|---|---|---|---|---|---|
| Subclass | Gregarinasina | Cryptogregaria | Coccidiasina | |||||
| Order | Archigregarinida | Eugregarinida | Neogregarinida | Cryptogregarida | Eucoccidiorida | Haemosporida | Piroplasmida | |
| Suborder | Adeleorina | Eimeriorina | ||||||
| Heteroxenous | ✕ | ✕ | ✕ | ✕ | ✔ | ✔ | ✔ | ✔ |
| Monoxenous | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✕ | ✕ |
| Merogony | ✕ | ✕ | ✕✔ a | ✔ | ✔ | ✔ | ✔ | ✔ |
| Syzygy | ✔ | ✔ | ✔ | ✔ | ✔ | ✕ | ✕ | ✕ |
| Gamogony In | Invertebrates | Invertebrates | Invertebrates | Vertebrates | Vertebrates | Vertebrates | Invertebrates | Invertebrates |
| Spiky-Rayed-Stages | ✕ | ✕ | ✕ | ✕ | ✕ | ✕ | ✕ | ✔ b |
| Free Gametocytes | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Differentiation of Gametes | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✕ |
| Microgamete motile | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✕✔ c |
| Macrogamete motile | ✕ | ✕ | ✕ | ✕ | ✕ | ✕ | ✔ | ✔ |
| Sexual Cycle Intracellular | ? | ? | ? | ✕ | ✔ | ? | ✔ | ✔ |
| Gametes Fusion | ✔ | ✔ d | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Zygote Formation | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Oocyst Formation | ✕ | ✕ | ✕ | ✔ | ✔ | ✔ | ✔ | ✔ |
| Final Product | Oocyst | Oocyst | Oocyst | Oocyst | Oocyst | Oocyst | Ookinete/Oocyst | Ookinete |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Bustos, T.; Feix, A.S.; Ruttkowski, B.; Joachim, A. Sexual Development in Non-Human Parasitic Apicomplexa: Just Biology or Targets for Control? Animals 2021, 11, 2891. https://doi.org/10.3390/ani11102891
Cruz-Bustos T, Feix AS, Ruttkowski B, Joachim A. Sexual Development in Non-Human Parasitic Apicomplexa: Just Biology or Targets for Control? Animals. 2021; 11(10):2891. https://doi.org/10.3390/ani11102891
Chicago/Turabian StyleCruz-Bustos, Teresa, Anna Sophia Feix, Bärbel Ruttkowski, and Anja Joachim. 2021. "Sexual Development in Non-Human Parasitic Apicomplexa: Just Biology or Targets for Control?" Animals 11, no. 10: 2891. https://doi.org/10.3390/ani11102891
APA StyleCruz-Bustos, T., Feix, A. S., Ruttkowski, B., & Joachim, A. (2021). Sexual Development in Non-Human Parasitic Apicomplexa: Just Biology or Targets for Control? Animals, 11(10), 2891. https://doi.org/10.3390/ani11102891