
Parturition in Mammals: Animal Models, Pain and Distress

 , , ,
, , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Physiological Response to Labour: Comparative Studies
3. Labour Pain (Parturition Pain Stimuli)
4. Origin and Transmission of Parturition Pain Stimuli

5. Factors Affecting Labour Pain, Dystocia

{kind=link}
{kind=link}
{kind=link}
| Factor That Increases Labour Pain | References |
|---|---|
| Maternal characteristics: Prolonged labours
| [102,103,104,105,106,107] |
6. Pain Assessment Scales
7. Managing Labour Pain
8. Consequences of Pain
9. Methods to Improve Pain Management during Parturition
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Van Rens, B.T.T.M.; Van Der Lende, T. Parturition in gilts: Duration of farrowing, birth intervals and placenta expulsion in relation to maternal, piglet and placental traits. Theriogenology 2004, 62, 331–352. [Google Scholar] [CrossRef]
- Lezama-García, K.; Mariti, C.; Mota-Rojas, D.; Martínez-Burnes, J.; Barrios-García, H.; Gazzano, A. Maternal behaviour in domestic dogs. Int. J. Vet. Sci. Med. 2019, 7, 20–30. [Google Scholar] [CrossRef]
- Ison, S.H.; Jarvis, S.; Hall, S.A.; Ashworth, C.J.; Rutherford, K.M.D. Periparturient behavior and physiology: Further insight into the farrowing process for primiparous and multiparous sows. Front. Vet. Sci. 2018, 5, 122. [Google Scholar] [CrossRef]
- Mędrzycka-Dabrowska, W.; Czyż-Szypenbejl, K.; Pietrzak, J. A review of randomized trials comparisons of epidural with parenteral forms of pain relief during labour and its impact on operative and cesarean delivery rate. Ginekol. Pol. 2018, 89, 460–467. [Google Scholar] [CrossRef]
- Lowe, N.K. The nature of labor pain. Am. J. Obstet. Gynecol. 2002, 186, S16–S24. [Google Scholar] [CrossRef]
- Loeser, J.D.; Treede, R.D. The Kyoto protocol of IASP basic pain terminology. Pain 2008, 137, 473–477. [Google Scholar] [CrossRef]
- IASP. Classification of Chronic Pain. In Descriptions of Chronic Pain Syndromes and Definitions Of Pain Terms; Merskey, H., Ed.; IASP Press: Seattle, WA, USA, 1994; p. 222. [Google Scholar]
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. Chronic pain as a symptom or a disease: The IASP classification of chronic pain for the international classification of diseases (ICD-11). Pain 2019, 160, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Aydede, M. Does the IASP definition of pain need updating? Pain Rep. 2019, 4, e777. [Google Scholar] [CrossRef]
- Hernández-Avalos, I.; Mota-Rojas, D.; Mora-Medina, P.; Martínez-Burnes, J.; Casas Alvarado, A.; Verduzco-Mendoza, A.; Lezama-García, K.; Olmos-Hernandez, A. Review of different methods used for clinical recognition and assessment of pain in dogs and cats. Int. J. Vet. Sci. Med. 2019, 7, 43–54. [Google Scholar] [CrossRef]
- Cohen, M.; Quintner, J.; van Rysewyk, S. Reconsidering the International Association for the Study of Pain definition of pain. Pain Rep. 2018, 3, e634. [Google Scholar] [CrossRef]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef]
- Labor, S.; Maguire, S. The Pain of Labour. Rev. Pain 2008, 2, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Nanji, J.A.; Carvalho, B. Pain management during labor and vaginal birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2020, 67, 100–112. [Google Scholar] [CrossRef]
- Catheline, G.; Touquet, B.; Besson, J.-M.; Lombard, M.-C. Parturition in the Rat. Anesthesiology 2006, 104, 1257–1265. [Google Scholar] [CrossRef]
- Mathews, K.; Kronen, P.W.; Lascelles, D.; Nolan, A.; Robertson, S.; Steagall, P.V.; Wright, B.; Yamashita, K. guidelines for recognition, assessment and treatment of Pain. J. Small Anim. Pract. 2014, 55, E10–E68. [Google Scholar] [CrossRef]
- Herskin, D.G.P. Pain in pigs: Characterisation, mechanisms an indicators. In Advances in Pig Welfare; Spinka, M., Ed.; Woodhead Publishing: Kidlington, UK, 2018; pp. 325–356. [Google Scholar]
- Gregory, N.G. (Ed.) Physiology and Behaviour of Animal Suffering; Blackwell Publishing: Oxford, UK, 2004; pp. 1–260. [Google Scholar]
- Mainau, E.; Manteca, X. Pain and discomfort caused by parturition in cows and sows. Appl. Anim. Behav. Sci. 2011, 135, 241–251. [Google Scholar] [CrossRef]
- da Rocha Pereira, R.; Franco, S.C.; Baldin, N. Pain and the protagonism of women in parturition. Braz. J. Anesthesiol. 2011, 61, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Trehan, G.; Gonzalez, M.N.; Kamel, I. Pain management during labor part 2: Techniques for labor analgesia. Top. Obs. Gynecol. 2016, 36, 1–10. [Google Scholar] [CrossRef]
- Morton, D.B.; Griffiths, P.H. Guidelines on the recognition of pain, distress and discomfort in experimental animals and an hypothesis for assessment. Vet. Rec. 1985, 116, 431–436. [Google Scholar] [CrossRef]
- Buchanan, K.L.; Burt de Perera, T.; Carere, C.; Carter, T.; Hailey, A.; Hubreacht, R.; Jenning, D.J.; Metcalfe, N.B.; Pitcher, T.E.; Péron, F.; et al. Guidelines for the Use of Animals Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 2012, 83, 301–309. [Google Scholar] [CrossRef]
- Veronesi, M.C.; Panzani, S.; Faustini, M.; Rota, A. An Apgar scoring system for routine assessment of newborn puppy viability and short-term survival prognosis. Theriogenology 2009, 72, 401–407. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Martínez-Burnes, J.; Napolitano, F.; Domínguez-Muñoz, M.; Guerrero-Legarreta, I.; Mora-Medina, P.; Ramírez-Necoechea, R.; Lezama-García, K.; González-Lozano, M. Dystocia: Factors affecting parturition in domestic animals. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2020, 15, 1–16. [Google Scholar] [CrossRef]
- Menon, R.; Bonney, E.A.; Condon, J.; Mesiano, S.; Taylor, R.N. Novel concepts on pregnancy clocks and alarms: Redundancy and synergy in human parturition. Hum. Reprod. Update 2016, 22, 535–560. [Google Scholar] [CrossRef]
- Hadley, E.E.; Richardson, L.S.; Torloni, M.R.; Menon, R. Gestational tissue inflammatory biomarkers at term labor: A systematic review of literature. Am. J. Reprod. Immunol. 2018, 79, e12776. [Google Scholar] [CrossRef]
- Bernitz, S.; Dalbye, R.; Øian, P.; Zhang, J.; Eggebø, T.M.; Blix, E. Study protocol: The Labor Progression Study, LAPS—Does the use of a dynamic progression guideline in labor reduce the rate of intrapartum cesarean sections in nulliparous women? A multicenter, cluster randomized trial in Norway. BMC Pregnancy Childbirth 2017, 17, 370. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.B.; Buhimschi, C.S.; Norwitz, E.R. Normal labor: Mechanism and duration. Obstet. Gynecol. Clin. N. Am. 2005, 32, 145–164. [Google Scholar] [CrossRef]
- Hutchison, J.; Mahdy, H.; Hutchison, J. Stages of Labor; StatPearls Publishing LLC: San Francisco, CA, USA, 2020. [Google Scholar]
- Maghoma, J.; Buchmann, E.J. Maternal and fetal risks associated with prolonged latent phase of labour. J. Obstet. Gynaecol. 2002, 22, 16–19. [Google Scholar] [CrossRef]
- Lauzon, L.; Hodnett, E.D. Antenatal education for self-diagnosis of the onset of active labour at term. Cochrane Database Syst. Rev. 1998, 1998, CD000935. [Google Scholar] [CrossRef]
- Chelmow, D.; Kilpatrick, S.J.; Laros, R.K. Maternal and neonatal outcomes after prolonged latent phase. Obstet. Gynecol. 1993, 81, 486–491. [Google Scholar]
- Hanley, G.E.; Munro, S.; Greyson, D.; Gross, M.M.; Hundley, V.; Spiby, H.; Janssen, P.A. Diagnosing onset of labor: A systematic review of definitions in the research literature. BMC Pregnancy Childbirth 2016, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Khasawneh, W.; Obeidat, N.; Yusef, D.; Alsulaiman, J.W. The impact of cesarean section on neonatal outcomes at a university-based tertiary hospital in Jordan. BMC Pregnancy Childbirth 2020, 20, 335. [Google Scholar] [CrossRef] [PubMed]
- Uystepruyst, C.; Coghe, J.; Dorts, T.; Harmegnies, N.; Delsemme, M.H.; Art, T.; Lekeux, P. Optimal timing of elective caesarean section in Belgian White and Blue breed of cattle: The calf’s point of view. Vet. J. 2002, 163, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Tribe, R.M.; Taylor, P.D.; Kelly, N.M.; Rees, D.; Sandall, J.; Kennedy, H.P. Parturition and the perinatal period: Can mode of delivery impact on the future health of the neonate? J. Physiol. 2018, 596, 5709–5722. [Google Scholar] [CrossRef]
- Sandall, J.; Tribe, R.M.; Avery, L.; Mola, G.; Visser, G.H.; Homer, C.S.; Gibbons, D.; Kelly, N.M.; Kennedy, H.P.; Kidanto, H.; et al. Short-term and long-term effects of caesarean section on the health of women and children. Lancet 2018, 392, 1349–1357. [Google Scholar] [CrossRef]
- Renfrew, M.J.; McFadden, A.; Bastos, M.H.; Campbell, J.; Channon, A.A.; Cheung, N.F.; Silva, D.R.A.D.; Downe, S.; Kennedy, H.P.; Malata, A.; et al. Midwifery and quality care: Findings from a new evidence-informed framework for maternal and newborn care. Lancet 2014, 384, 1129–1145. [Google Scholar] [CrossRef]
- Behnia, F.; Taylor, B.D.; Woodson, M.; Kacerovsky, M.; Hawkins, H.; Fortunato, S.J.; Saade, G.R.; Menon, R. Chorioamniotic membrane senescence: A signal for parturition? Am. J. Obstet. Gynecol. 2015, 213, 359.e1–359.e16. [Google Scholar] [CrossRef]
- Menon, R. Initiation of human parturition: Signaling from senescent fetal tissues via extracellular vesicle mediated paracrine mechanism. Obstet. Gynecol. Sci. 2019, 62, 199. [Google Scholar] [CrossRef]
- Hashemi Asl, B.M.; Vatanchi, A.; Golmakani, N.; Najafi, A. Relationship between behavioral indices of pain during labor pain with pain intensity and duration of delivery. Electron. Physician 2018, 10, 6240–6248. [Google Scholar] [CrossRef] [Green Version]
- Kovác, G.; Tóthová, C.; Nagy, O.; Seidel, H. Acute phase proteins during the reproductive cycle of sows. Acta Vet. Brno. 2008, 58, 459–466. [Google Scholar] [CrossRef]
- Verheyen, A.J.M.; Maes, D.G.D.; Mateusen, B.; Deprez, P.; Janssens, G.P.J.; de Lange, L.; Counotte, G. Serum biochemical reference values for gestating and lactating sows. Vet. J. 2007, 174, 92–98. [Google Scholar] [CrossRef]
- Kostro, K.; Wawron, W.; Szczubiał, M.; Luft-Deptula, D.; Gliński, Z. C-reactive protein in monitoring and evaluation of effects of therapy of the MMA syndrome of sows. Pol. J. Vet. Sci. 2003, 6, 235–238. [Google Scholar]
- Winkler, M.; Kemp, B.; Fischer, D.; Maul, H.; Hlubekl, M.; Rath, W. Tissue concentrations of cytokines in the lower uterine segment during preterm parturition. J. Perinat. Med. 2001, 29, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.; Winkler, M.; Maas, A.; Maul, H.; Ruck, P.; Reineke, T.; Rath, W. Cytokine concentrations in the amniotic fluid during parturition at term: Correlation to lower uterine segment values and to labor. Acta Obstet. Gynecol. Scand. 2002, 81, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.; Kemp, B.; Fischer, D.C.; Ruck, P.; Rath, W. Expression of adhesion molecules in the lower uterine segment during term and preterm parturition. Microsc. Res. Tech. 2003, 60, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Condon, J.C.; Hardy, D.B.; Kovaric, K.; Mendelson, C.R. Up-regulation of the progesterone receptor (pr)-c isoform in laboring myometrium by activation of nuclear factor-κb may contribute to the onset of labor through inhibition of PR function. Mol. Endocrinol. 2006, 20, 764–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindström, T.M.; Bennett, P.R. The role of nuclear factor kappa B in human labour. Reproduction 2005, 130, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Terzidou, V.; Lee, Y.; Lindström, T.; Johnson, M.; Thornton, S.; Bennett, P.R. Regulation of the human oxytocin receptor by nuclear factor-κB and CCAAT/Enhancer-binding protein-β. J. Clin. Endocrinol. Metab. 2006, 91, 2317–2326. [Google Scholar] [CrossRef]
- Thomson, A.J.; Telfer, J.F.; Young, A.; Campbell, S.; Stewart, C.J.R.; Cameron, I.T.; Greer, I.A.; Norman, J.E. Leukocytes infiltrate the myometrium during human parturition: Further evidence that labour is an inflammatory process. Hum. Reprod. 1999, 14, 229–236. [Google Scholar] [CrossRef]
- Young, A.; Thomson, A.J.; Ledingham, M.; Jordan, F.; Greer, I.A.; Norman, J.E. Immunolocalization of proinflammatory cytokines in myometrium, cervix, and fetal membranes during human parturition at term1. Biol. Reprod. 2002, 66, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Osman, I.; Young, A.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte density and proinflammatory mediator expression in regional human fetal membranes and decidua before and during labot at term. J. Soc. Gynecol. Investig. 2006, 13, 97–103. [Google Scholar] [CrossRef]
- Hutchinson, J.L.; Rajagopal, S.P.; Yuan, M.; Norman, J.E. Lipopolysaccharide promotes contraction of uterine myocytes via activation of Rho/ROCK signaling pathways. FASEB J. 2014, 28, 94–105. [Google Scholar] [CrossRef]
- Rajagopal, S.P.; Hutchinson, J.L.; Dorward, D.A.; Rossi, A.G.; Norman, J.E. Crosstalk between monocytes and myometrial smooth muscle in culture generates synergistic pro-inflammatory cytokine production and enhances myocyte contraction, with effects opposed by progesterone. Mol. Hum. Reprod. 2015, 21, 672–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, D.B.; Janowski, B.A.; Corey, D.R.; Mendelson, C.R. Progesterone receptor plays a major antiinflammatory role in human myometrial cells by antagonism of nuclear factor-κB activation of cyclooxygenase 2 expression. Mol. Endocrinol. 2006, 20, 2724–2733. [Google Scholar] [CrossRef]
- Condon, J.C.; Jeyasuria, P.; Faust, J.M.; Mendelson, C.R. Surfactant protein secreted by the maturing mouse fetal lung acts as a hormone that signals the initiation of parturition. Proc. Natl. Acad. Sci. USA 2004, 101, 4978–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, R.; Pease, J.E.; Sooranna, S.R.; Viney, J.M.; Nelson, S.M.; Myatt, L.; Bennett, P.R.; Johnson, M.R. Stretch and inflammatory cytokines drive myometrial chemokine expression via NF-κB activation. Endocrinology 2012, 153, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Herbert, B.; Sooranna, G.R.; Orsi, N.M.; Edey, L.; Dasgupta, T.; Sooranna, S.R.; Yellon, S.M.; Johnson, M.R. Is myometrial inflammation a cause or a consequence of term human labour? J. Endocrinol. 2017, 235, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Whitburn, L.Y.; Jones, L.E.; Davey, M.A.; McDonald, S. The nature of labour pain: An updated review of the literature. Women Birth 2019, 32, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Runcan, E.E.; Coutinho da Silva, M.A. Whelping and dystocia: Maximizing success of medical management. Top. Companion Anim. Med. 2018, 33, 12–16. [Google Scholar] [CrossRef]
- Taverne, M.; Noakes, D. Parturition and the care of parturient animals, including the newborn. In Arthur’s Veterinary Reproduction and Obstetrics, 9th ed.; Noakes, D.E., Parkinson, T.J., England, G.C.W., Eds.; Saunders Elsevier: Philadelphia, PA, USA, 2001; pp. 155–187. [Google Scholar]
- Challis, J.R.G.; Lye, S.J. Parturition. Oxf. Rev. Reprod. Biol. 1986, 8, 61–129. [Google Scholar]
- Jenkin, G.; Young, I.R. Mechanisms responsible for parturition; the use of experimental models. In Proceedings of the Animal Reproduction Science. Anim. Reprod. Sci. 2004, 82–83, 567–581. [Google Scholar] [CrossRef]
- Taverne, M.A.M. Physiology of parturition. Anim. Reprod. Sci. 1992, 28, 433–440. [Google Scholar] [CrossRef]
- Sennström, M.B.; Ekman, G.; Westergren-Thorsson, G.; Malmström, A.; Byström, B.; Endrésen, U.; Mlambo, N.; Norman, M.; Ståbi, B.; Brauner, A. Human cervical ripening, an inflammatory process mediated by cytokines. Mol. Hum. Reprod. 2000, 6, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellon, S.M. Immunobiology of Cervix Ripening. Front. Immunol. 2019, 10, 3156. [Google Scholar] [CrossRef] [Green Version]
- Mousa, O.; Abdelhafez, A.A.; Abdelraheim, A.R.; Yousef, A.M.; Ghaney, A.A.; El Gelany, S. Perceptions and practice of labor pain-relief methods among health professionals conducting delivery in minia maternity units in Egypt. Obstet. Gynecol. Int. 2018, 2018, 3060953. [Google Scholar] [CrossRef] [Green Version]
- Serruya, S.J.; Cecatti, J.G.; di Giacomo do Lago, T. Programa de Humanização no Pré-natal e Nascimento do Ministério da Saúde no Brasil: Resultados iniciais. Cad. Saude Publica 2004, 20, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, S.; Permezel, M. Physiology of pain in labour. Baillieres. Clin. Obstet. Gynaecol. 1998, 12, 347–362. [Google Scholar] [CrossRef]
- Miquelutti, M.A.; Silveira, C.; Cecatti, J.G. Kinesiologic tape for labor pain control: Randomized controlled trial. Physiother. Theory Pract. 2019, 35, 614–621. [Google Scholar] [CrossRef]
- Stenglin, M.; Foureur, M. Designing out the Fear Cascade to increase the likelihood of normal birth. Midwifery 2013, 29, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Taverne, M.A.M.; Naaktgeboren, C.; Elsaesser, F.; Forsling, M.L.; van der Weyden, G.C.; Ellendorff, F.; Smidt, D. Myometrial electrical activity and plasma concentrations of progesterone, estrogens and oxytocin during late pregnancy and parturition in the miniature pig. Biol. Reprod. 1979, 21, 1125–1134. [Google Scholar] [CrossRef]
- Gilbert, C.L.; Goode, J.A.; McGrath, T.J. Pulsatile secretion of oxytocin during parturition in the pig: Temporal relationship with fetal expulsion. J. Physiol. 1994, 475, 129–137. [Google Scholar] [CrossRef]
- Olcese, J.; Beesley, S. Clinical significance of melatonin receptors in the human myometrium. Fertil. Steril. 2014, 102, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, J.T.; Puttaramu, R.; Word, R.A.; Olcese, J. Melatonin synergizes with oxytocin to enhance contractility of human myometrial smooth muscle cells. J. Clin. Endocrinol. Metab. 2009, 94, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Bennett, P.R.; Terzidou, V. Advances in the role of oxytocin receptors in human parturition. Mol. Cell. Endocrinol. 2017, 449, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Gorodeski, G.I.; Sheean, L.A.; Utian, W.H. Myometrial oxytocin receptors levels in the pregnant rat are higher in distal than in proximal portions of the horn and correlate with disparate oxytocin responsive myometrial contractility in these segments. Endocrinology 1990, 127, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.-R. Prostaglandin F2alpha and oxytocin interactions in ovarian and uterine function. J. Steroid Biochem. 1987, 27, 1073–1080. [Google Scholar] [CrossRef]
- Soloff, M.S. Uterine receptor for oxytocin: Effects of estrogen. Biochem. Biophys. Res. Commun. 1975, 65, 205–212. [Google Scholar] [CrossRef]
- Gimpl, G.; Fahrenholz, F. The oxytocin receptor system: Structure, function, and regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.; Peters, T.J.; Campbell, R. Techniques to reduce perineal pain during spontaneous vaginal delivery and perineal suturing: A UK survey of midwifery practice. Midwifery 2005, 21, 154–160. [Google Scholar] [CrossRef]
- Lowe, N.K.; Roberts, J.E. The convergence between in-labor report and postpartum recall of parturition pain. Res. Nurs. Health 1988, 11, 11–21. [Google Scholar] [CrossRef]
- Melzack, R.; Schaffelberg, D. Low-back pain during labor. Am. J. Obstet. Gynecol. 1987, 156, 901–905. [Google Scholar] [CrossRef]
- Schabel, J.E.; Poppers, P.J. Lumbar Epidural Analgesia for Labor and Vaginal Delivery. Gynecol. Obstet. Investig. 1997, 44, 73–81. [Google Scholar] [CrossRef]
- Jarvis, S.; Lawrence, A..; McLean, K..; Chirnside, J.; Deans, L..; Calvert, S. The effect of environment on plasma cortisol and β-endorphin in the parturient pig and the involvement of endogenous opioids. Anim. Reprod. Sci. 1998, 52, 139–151. [Google Scholar] [CrossRef]
- Jarvis, S.; McLean, K.A.; Calvert, S.K.; Deans, L.A.; Chirnside, J.; Lawrence, A.B. The responsiveness of sows to their piglets in relation to the length of parturition and the involvement of endogenous opioids. Appl. Anim. Behav. Sci. 1999, 63, 195–207. [Google Scholar] [CrossRef]
- Lawrence, A.B.; Petherick, J.C.; McLean, K.; Gilbert, C.L.; Chapman, C.; Russell, J.A. Naloxone prevents interruption of parturition and increases plasma oxytocin following environmental disturbance in parturient sows. Physiol. Behav. 1992, 52, 917–923. [Google Scholar] [CrossRef]
- Jarvis, S.; McLean, K.A.; Chirnside, J.; Deans, L.A.; Calvert, S.K.; Molony, V.; Lawrence, A.B. Opioid-mediated changes in nociceptive threshold during pregnancy and parturition in the sow. Pain 1997, 72, 153–159. [Google Scholar] [CrossRef]
- Ison, S.H.; Clutton, R.E.; Di Giminiani, P.; Rutherford, K.M.D. A review of pain assessment in pigs. Front. Vet. Sci. 2016, 3, 108. [Google Scholar] [CrossRef] [PubMed]
- González-Lozano, M.; Mota-Rojas, D.; Orihuela, A.; Martínez-Burnes, J.; Di Francia, A.; Braghieri, A.; Berdugo-Gutiérrez, J.; Mora-Medina, P.; Ramírez-Necoechea, R.; Napolitano, F. Review: Behavioral, physiological, and reproductive performance of buffalo cows during eutocic and dystocic parturitions. Appl. Anim. Sci. 2020, 36, 407–422. [Google Scholar] [CrossRef]
- Hautakangas, T.; Palomäki, O.; Eidstø, K.; Huhtala, H.; Uotila, J. Impact of obesity and other risk factors on labor dystocia in term primiparous women: A case control study. BMC Pregnancy Childbirth 2018, 18, 304. [Google Scholar] [CrossRef]
- Sandström, A.; Altman, M.; Cnattingius, S.; Johansson, S.; Ahlberg, M.; Stephansson, O. Durations of second stage of labor and pushing, and adverse neonatal outcomes: A population-based cohort study. J. Perinatol. 2017, 37, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, M.; Nematollahi, A.; Farahmand, M.; Amooee, S. The effect of two-staged warm compress on the pain duration of first and second labor stages and apgar score in prim gravida women: A Randomized clinical trial. J. Caring Sci. 2018, 7, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Rooks, J.P. Labor pain management other than neuraxial: What do we know and where do we go next? Birth 2012, 39, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Slone, M.E.; James, S.R.; Murray, S.S.; Nelson, K.A.; Aswill, J.W. Maternal-Child Nursing, 5th ed.; Elsevier Saunders: St. Louis, MI, USA, 2018; pp. 1–1592. [Google Scholar]
- Beagley, J.C.; Whitman, K.J.; Baptiste, K.E.; Scherzer, J. Physiology and treatment of retained fetal membranes in cattle. J. Vet. Intern. Med. 2010, 24, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Mollo, A.; Veronesi, M.C.; Cairoli, F.; Soldano, F. The use of oxytocin for the reduction of cow placental retention, and subsequent endometritis. Anim. Reprod. Sci. 1997, 48, 47–51. [Google Scholar] [CrossRef]
- Zobel, R. Endometritis in simmental cows: Incidence, causes, and therapy options. Turkish J. Vet. Anim. Sci. 2013, 37, 134–140. [Google Scholar] [CrossRef]
- Barraclough, R.A.C.; Shaw, D.J.; Boyce, R.; Haskell, M.J.; Macrae, A.I. The behavior of dairy cattle in late gestation: Effects of parity and dystocia. J. Dairy Sci. 2020, 103, 14–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.W.; Caughey, A.B. Defining and managing normal and abnormal second stage of labor. Obstet. Gynecol. Clin. N. Am. 2017, 44, 547–566. [Google Scholar] [CrossRef]
- Zaki, M.N.; Hibbard, J.U.; Kominiarek, M.A. Contemporary labor patterns and maternal age. Obstet. Gynecol. 2013, 122, 1018–1024. [Google Scholar] [CrossRef]
- Laughon, S.K.; Berghella, V.; Reddy, U.M.; Sundaram, R.; Lu, Z.; Hoffman, M.K. Neonatal and maternal outcomes with prolonged second stage of labor. Obstet. Gynecol. 2014, 124, 57–67. [Google Scholar] [CrossRef]
- Senécal, J.; Xiong, X.; Fraser, W.D. Effect of fetal position on second-stage duration and labor outcome. Obstet. Gynecol. 2005, 105, 763–772. [Google Scholar] [CrossRef]
- Feinstein, U.; Sheiner, E.; Levy, A.; Hallak, M.; Mazor, M. Risk factors for arrest of descent during the second stage of labor. Int. J. Gynecol. Obstet. 2002, 77, 7–14. [Google Scholar] [CrossRef]
- Sizer, A.R.; Evans, J.; Bailey, S.M.; Wiener, J. A second-stage partogram. Obstet. Gynecol. 2000, 96, 678–683. [Google Scholar] [CrossRef]
- Darvelid, A.W.; Linde-Forsberg, C. Dystocia in the bitch: A retrospective study of 182 cases. J. Small Anim. Pract. 1994, 35, 402–407. [Google Scholar] [CrossRef]
- Linde Forsberg, C.; Persson, G. A survey of dystocia in the Boxer breed. Acta Vet. Scand. 2007, 49, 8. [Google Scholar] [CrossRef] [Green Version]
- Veiga, G.A.L.; Milazzotto, M.P.; Nichi, M.; Lúcio, C.F.; Silva, L.C.G.; Angrimani, D.S.R.; Vannucchi, C.I. Gene expression of estrogen and oxytocin receptors in the uterus of pregnant and parturient bitches. Braz. J. Med. Biol. Res. 2015, 48, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Egloff, S.; Reichler, I.M.; Kowalewski, M.P.; Keller, S.; Goericke-Pesch, S.; Balogh, O. Uterine expression of smooth muscle alpha- and gamma-actin and smooth muscle myosin in bitches diagnosed with uterine inertia and obstructive dystocia. Theriogenology 2020, 156, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Lúcio, C.; Silva, L.; Rodrigues, J.; Veiga, G.; Vannucchi, C. Peripartum haemodynamic status of bitches with normal birth or dystocia. Reprod. Domest. Anim. 2009, 44, 133–136. [Google Scholar] [CrossRef]
- Münnich, A.; Küchenmeister, U. Dystocia in numbers—Evidence-based parameters for intervention in the dog: Causes for dystocia and treatment recommendations. Reprod. Domest. Anim. 2009, 44, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.S.; Axnér, E.; Öhlund, M.; Möller, L.; Egenvall, A. Dystocia in the cat evaluated using an insurance database. J. Feline Med. Surg. 2017, 19, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Dalayeun, J.F.; Norès, J.M.; Bergal, S. Physiology of β-endorphins. A close-up view and a review of the literature. Biomed. Pharmacother. 1993, 47, 311–320. [Google Scholar] [CrossRef]
- Farmer, S.; Robert, C. Hormonal, behavioural and performance characteristics of Meishan sows during pregnancy and lactation. Can. J. Anim. Sci. 2003, 83, 1–12. [Google Scholar] [CrossRef]
- van Dijk, A.J.; van Rens, B.T.T.M.; van der Lende, T.; Taverne, M.A.M. Factors affecting duration of the expulsive stage of parturition and piglet birth intervals in sows with uncomplicated, spontaneous farrowings. Theriogenology 2005, 64, 1573–1590. [Google Scholar] [CrossRef]
- Oliviero, C.; Heinonen, M.; Valros, A.; Peltoniemi, O. Environmental and sow-related factors affecting the duration of farrowing. Anim. Reprod. Sci. 2010, 119, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Björkman, S.; Oliviero, C.; Kauffold, J.; Soede, N.M.; Peltoniemi, O.A.T. Prolonged parturition and impaired placenta expulsion increase the risk of postpartum metritis and delay uterine involution in sows. Theriogenology 2018, 106, 87–92. [Google Scholar] [CrossRef]
- Borges, V.F.; Bernardi, M.L.; Bortolozzo, F.P.; Wentz, I. Risk factors for stillbirth and foetal mummification in four Brazilian swine herds. Prev. Vet. Med. 2005, 70, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, M.H.; Friend, D.W. Factors influencing, and repeatability of the duration of farrowing in yorkshire sows. Can. J. Anim. Sci. 1981, 61, 17–22. [Google Scholar] [CrossRef]
- Vestergaard, K.H.L. Tethered versus loose sows: Ethological observations and measures of productivity I. Ethological observations during pregnancy and farrowing. Ann. Rech. Vet. 1984, 15, 245–256. [Google Scholar] [PubMed]
- Fraser, D.; Phillips, P.A.; Thompson, B.K. Farrowing behaviour and stillbirth in two environments: An evaluation of the restraint-stillbirth hypothesis. Appl. Anim. Behav. Sci. 1997, 55, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Holm, B.; Bakken, M.; Vangen, O.; Rekaya, R. Genetic analysis of age at first service, return rate, litter size, and weaning-to-first service interval of gilts and sows1. J. Anim. Sci. 2005, 83, 41–48. [Google Scholar] [CrossRef]
- Holm, B.; Bakken, M.; Vangen, O.; Rekaya, R. Genetic analysis of litter size, parturition length, and birth assistance requirements in primiparous sows using a joint linear-threshold animal model1. J. Anim. Sci. 2004, 82, 2528–2533. [Google Scholar] [CrossRef]
- Oliviero, C.; Junnikkala, S.; Peltoniemi, O. The challenge of large litters on the immune system of the sow and the piglets. Reprod. Domest. Anim. 2019, 54, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Pearodwong, P.; Muns, R.; Tummaruk, P. Prevalence of constipation and its influence on post-parturient disorders in tropical sows. Trop. Anim. Health Prod. 2016, 48, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Ison, S.H.; Jarvis, S.; Rutherford, K.M.D. The identification of potential behavioural indicators of pain in periparturient sows. Res. Vet. Sci. 2016, 109, 114–120. [Google Scholar] [CrossRef]
- Nagel, C.; Aurich, C.; Aurich, J. Stress effects on the regulation of parturition in different domestic animal species. Anim. Reprod. Sci. 2019, 207, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Nagel, C.; Erber, R.; Ille, N.; von Lewinski, M.; Aurich, J.; Möstl, E.; Aurich, C. Parturition in horses is dominated by parasympathetic activity of the autonomous nervous system. Theriogenology 2014, 82, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Roussel, S.; Hemsworth, P.H.; Boissy, A.; Duvaux-Ponter, C. Effects of repeated stress during pregnancy in ewes on the behavioural and physiological responses to stressful events and birth weight of their offspring. Appl. Anim. Behav. Sci. 2004, 85, 259–276. [Google Scholar] [CrossRef]
- Ritter, C.; Beaver, A.; von Keyserlingk, M.A.G. The complex relationship between welfare and reproduction in cattle. Reprod. Domest. Anim. 2019, 54, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landa, L. Pain in domestic animals and how to assess it: A review. Vet. Med. 2012, 57, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Herr, K.; Coyne, P.J.; Key, T.; Manworren, R.; McCaffery, M.; Merkel, S.; Pelosi-Kelly, J.; Wild, L. Pain Assessment in the Nonverbal Patient: Position Statement with Clinical Practice Recommendations. Pain Manag. Nurs. 2006, 7, 44–52. [Google Scholar] [CrossRef]
- Molony, V.; Kent, J.E. Assessment of acute pain in farm animals using behavioral and physiological measurements. J. Anim. Sci. 1997, 75, 266. [Google Scholar] [CrossRef]
- Mellor, D.; Stafford, K.; Todd, S.; Lowe, T.; Gregory, N.; Bruce, R.; Ward, R. A comparison of catecholamine and cortisol responses of young lambs and calves to painful husbandry procedures. Aust. Vet. J. 2002, 80, 228–233. [Google Scholar] [CrossRef]
- Keating, S.C.J.; Thomas, A.A.; Flecknell, P.A.; Leach, M.C. Evaluation of EMLA Cream for Preventing Pain during Tattooing of Rabbits: Changes in Physiological, Behavioural and Facial Expression Responses. PLoS ONE 2012, 7, e44437. [Google Scholar] [CrossRef] [Green Version]
- Flecknell, P.A.; Kirk, A.J.B.; Liles, J.H.; Hayes, P.H.; Dark, J.H. Post-operative analgesia following thoracotomy in the dog: An evaluation of the effects of bupivacaine intercostal nerve block and nalbuphine on respiratory function. Lab. Anim. 1991, 25, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Kalliokoski, O.; Abelson, K.S.P.; Koch, J.; Boschian, A.; Thormose, S.F.; Fauerby, N.; Rasmussen, R.S.; Johansen, F.F.; Hau, J. The effect of voluntarily ingested buprenorphine on rats subjected to surgically induced global cerebral ischaemia. In Vivo 2010, 24, 641–646. [Google Scholar]
- Schiavenato, M.; Craig, K.D. Pain assessment as a social transaction: Beyond the “gold standard”. Clin. J. Pain 2010, 26, 667–676. [Google Scholar] [CrossRef]
- Holton, L.; Pawson, P.; Nolan, A.; Reid, J.; Scott, E.M. Development of a behaviour-based scale to measure acute pain in dogs. Vet. Rec. 2001, 148, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.M.; Reid, J.; Scott, E.M.; Holton, L.L.; Nolan, A.M. Application of a scaling model to establish and validate an interval level pain scale for assessment of acute pain in dogs. Am. J. Vet. Res. 2005, 66, 2154–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gónzalez-Lozano, M.; Trujillo, O.M.E.; Becerril-Herrera, M.; Alonso-Spilsbury, M.; Rosales-Torres, A.M.; Mota-Rojas, D. Uterine activity and fetal electronic monitoring in parturient sows treated with vetrabutin chlorhydrate. J. Vet. Pharmacol. Therap. 2009, 33, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B. Analgesia for the critically ill dog or cat: An update. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Roughan, J.V.; Wright-Williams, S.L.; Flecknell, P.A. Automated analysis of postoperative behaviour: Assessment of HomeCageScan as a novel method to rapidly identify pain and analgesic effects in mice. Lab. Anim. 2009, 43, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Conklin, D.R.; Liu, B.; Ririe, D.G.; Eisenach, J.C. Assessment of behavior during labor in rats and effect of intrathecal morphine. Anesthesiology 2008, 108, 1081–1086. [Google Scholar] [CrossRef] [Green Version]
- Castrén, H.; Algers, B.; de Passillé, A.-M.; Rushen, J.; Uvnäs-Moberg, K. Preparturient variation in progesterone, prolactin, oxytocin and somatostatin in relation to nest building in sows. Appl. Anim. Behav. Sci. 1993, 38, 91–102. [Google Scholar] [CrossRef]
- Mainau, E.; Dalmau, A.; Ruiz-de-la-Torre, J.L.; Manteca, X. A behavioural scale to measure ease of farrowing in sows. Theriogenology 2010, 74, 1279–1287. [Google Scholar] [CrossRef]
- Rutherford, K.M.D.; Robson, S.K.; Donald, R.D.; Jarvis, S.; Sandercock, D.A.; Scott, E.M.; Nolan, A.M.; Lawrence, A.B. Pre-natal stress amplifies the immediate behavioural responses to acute pain in piglets. Biol. Lett. 2009, 5, 452–454. [Google Scholar] [CrossRef] [PubMed]
- de Fátima de Assunção Braga, A.; Carvalho, V.H.; da Silva Braga, F.S.; Pereira, R.I.C. Bloqueio combinado raquiperidural para analgesia de parto. Estudo comparativo com bloqueio peridural contínuo. Braz. J. Anesthesiol. 2019, 69, 7–12. [Google Scholar] [CrossRef]
- Leeman, L.; Fontaine, P.; King, V.; Klein, M.C.; Ratcliffe, S. The nature and management of labor pain: Part I. Nonpharmacologic pain relief. Am. Fam. Physician 2003, 68, 1109–1112. [Google Scholar]
- Orihuela, A.; Mota-Rojas, D.; Strappini, A.; Serrapica, F.; Braghieri, A.; Mora-Medina, P.; Napolitano, F. Neurophysiological Mechanisms of Cow–Calf Bonding in Buffalo and Other Farm Animals. Animals 2021, 11, 1968. [Google Scholar] [CrossRef]
- Eisenach, J. Neurophysiology of labour pain. Eur. Soc. Anaesthesiol. 2010, 1, 1–4. [Google Scholar]
- Volmanen, P.; Akural, E.I.; Raudaskoski, T.; Alahuhta, S. Remifentanil in Obstetric Analgesia: A Dose-Finding Study. Anesth. Analg. 2002, 94, 913–917. [Google Scholar] [CrossRef]
- Michelsen, L.G.; Hug, C.C. The pharmacokinetics of remifentanil. J. Clin. Anesth. 1996, 8, 679–682. [Google Scholar] [CrossRef]
- Olofsson, C.; Ekblom, A.; Ekman-Ordeberg, G.; Hjelm, A.; Irestedt, L. Lack of analgesic effect of systemically administered morphine or pethidine on labour pain. BJOG An. Int. J. Obstet. Gynaecol. 1996, 103, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.C. Parenteral opioids for labor analgesia. Clin. Obstet. Gynecol. 2003, 46, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.J.A.; MacArthur, C.; Hewitt, C.A.; Handley, K.; Gao, F.; Beeson, L.; Daniels, J. Intravenous remifentanil patient-controlled analgesia versus intramuscular pethidine for pain relief in labour (RESPITE): An open-label, multicentre, randomised controlled trial. Lancet 2018, 392, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Shaaban, O.M.; Abbas, A.M.; Mohamed, R.A.; Hafiz, H. Lack of pain relief during labor is blamable for the increase in the women demands towards cesarean delivery: A cross-sectional study. Facts Views Vis. ObGyn 2017, 9, 175–180. [Google Scholar] [PubMed]
- Loughnan, B.A.; Carli, F.; Romney, M.; Doré, C.J.; Gordon, H. Randomized controlled comparison of epidural bupivacaine versus pethidine for analgesia in labour. Br. J. Anaesth. 2000, 84, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Sheiner, E.; Shoham-Vardi, I.; Sheiner, E.K.; Press, F.; Hackmon-Ram, R.; Mazor, M.; Katz, M. A comparison between the effectiveness of epidural analgesia and parenteral pethidine during labor. Arch. Gynecol. Obstet. 2000, 263, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, N.; Barbarossa, A.; Cunto, M.; Ballotta, G.; Zambelli, D.; Armorini, S.; Zaghini, A.; Lambertini, C. Evaluation of methadone concentrations in bitches and in umbilical cords after epidural or systemic administration for caesarean section: A randomized trial. Vet. Anaesth. Analg. 2019, 46, 375–383. [Google Scholar] [CrossRef]
- Mathews, K.A. Pain management for the pregnant, lactating, and neonatal to pediatric cat and dog. Vet. Clin. North Am. Small Anim. Pract. 2008, 38, 1291–1308. [Google Scholar] [CrossRef] [PubMed]
- Mainau, E.; Temple, D.; Manteca, X. Experimental study on the effect of oral meloxicam administration in sows on pre-weaning mortality and growth and immunoglobulin G transfer to piglets. Prev. Vet. Med. 2016, 126, 48–53. [Google Scholar] [CrossRef]
- Ison, S.H.; Jarvis, S.; Ashworth, C.J.; Rutherford, K.M.D. The effect of post-farrowing ketoprofen on sow feed intake, nursing behaviour and piglet performance. Livest. Sci. 2017, 202, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.J.; Sorrell, J.T.; Rodgers, C.S.; Lebeck, M.M. Anxiety sensitivity as a predictor of labor pain. Eur. J. Pain 2006, 10, 263–270. [Google Scholar] [CrossRef]
- Jones, L.E.; Whitburn, L.Y.; Davey, M.-A.; Small, R. Assessment of pain associated with childbirth: Women’s perspectives, preferences and solutions. Midwifery 2015, 31, 708–712. [Google Scholar] [CrossRef]
- Aneiros, F.; Vazquez, M.; Valiño, C.; Taboada, M.; Sabaté, S.; Otero, P.; Costa, J.; Carceller, J.; Vázquez, R.; Díaz-Vieito, M.; et al. Does epidural versus combined spinal-epidural analgesia prolong labor and increase the risk of instrumental and cesarean delivery in nulliparous women? J. Clin. Anesth. 2009, 21, 94–97. [Google Scholar] [CrossRef]
- National Research Council. Recognition and Alleviation of Pain in Laboratory Animals; National Academies Press: Washington, DC, USA, 2009; pp. 1–199. [Google Scholar]
- De Cramer, K.G.M.; Joubert, K.E.; Nöthling, J.O. Puppy survival and vigor associated with the use of low dose medetomidine premedication, propofol induction and maintenance of anesthesia using sevoflurane gas-inhalation for cesarean section in the bitch. Theriogenology 2017, 96, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S. Anaesthetic management for caesarean sections in dogs and cats. Practice 2016, 38, 327–339. [Google Scholar] [CrossRef]
- Gabas, D.T.; Oliva, V.N.L.S.; Matsuba, L.M.; Perri, S.H.V. Estudo clínico e cardiorrespiratório em cadelas gestantes com parto normal ou submetidas à cesariana sob anestesia inalatória com sevofluorano. Arq. Bras. Med. Vet. Zootec. 2006, 58, 518–524. [Google Scholar] [CrossRef] [Green Version]
- Luna, S.P.L.; Basílio, A.C.; Steagall, P.V.M.; Machado, L.P.; Moutinho, F.Q.; Takahira, R.K.; Brandão, C.V.S. Evaluation of adverse effects of long-term oral administration of carprofen, etodolac, flunixin meglumine, ketoprofen, and meloxicam in dogs. Am. J. Vet. Res. 2007, 68, 258–264. [Google Scholar] [CrossRef]
- Mullins, K.B.; Thomason, J.M.; Lunsford, K.V.; Pinchuk, L.M.; Langston, V.C.; Wills, R.W.; McLaughlin, R.M.; Mackin, A.J. Effects of carprofen, meloxicam and deracoxib on platelet function in dogs. Vet. Anaesth. Analg. 2012, 39, 206–217. [Google Scholar] [CrossRef]
- McLean, M.K.; Khan, S.A. Toxicology of frequently encountered nonsteroidal anti-inflammatory drugs in dogs and cats. Vet. Clin. North Am. Small Anim. Pract. 2018, 48, 969–984. [Google Scholar] [CrossRef] [PubMed]
- Cima, G. Synthetic Opioids Put Police Dogs at Risk. 18 January 2018. Available online: https://www.avma.org/javma-news/2018-02-01/synthetic-opioids-put-police-dogs-risk (accessed on 22 February 2021).
- Huxley, J.N.; Whay, H.R. Current attitudes of cattle practitioners to pain and the use of analgesics in cattle. Vet. Rec. 2006, 159, 662–668. [Google Scholar] [CrossRef]
- Sabaté, D.; Salichs, M.; Bosch, J.; Ramó, P.H.J. Efficacy of ketoprofen in the reduction of pre-weaning piglet mortality associated with sub-clinical forms of post-partum dysgalactia syndrome in sows. Pig J. 2012, 67, 1923. [Google Scholar]
- Mainau, E.; Ruiz-de-la-Torre, J.L.; Dalmau, A.; Salleras, J.M.; Manteca, X. Effects of meloxicam (Metacam®) on post-farrowing sow behaviour and piglet performance. Animal 2012, 6, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homedes, J.; Salichs, M.; Sabaté, D.; Sust, M.; Fabre, R. Effect of ketoprofen on pre-weaning piglet mortality on commercial farms. Vet. J. 2014, 201, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Viitasaari, E.; Hänninen, L.; Heinonen, M.; Raekallio, M.; Orro, T.; Peltoniemi, O.; Valros, A. Effects of post-partum administration of ketoprofen on sow health and piglet growth. Vet. J. 2013, 198, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Brownridge, P. The nature and consequences of childbirth pain. Eur. J. Obstet. Gynecol. Reprod. Biol. 1995, 59, S9–S15. [Google Scholar] [CrossRef]
- Frey, H.; Tuuli, M.; Cortez, S.; Odibo, A.; Roehl, K.; Shanks, A.; Macones, G.; Cahill, A. Does delayed pushing in the second stage of labor impact perinatal outcomes? Am. J. Perinatol. 2012, 29, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, M.; Moradi, Z.; Hadianfard, M.J.; Zare, N.; Jowkar, A. Comparison of the effect of mono-stage and bi- stage acupressure at sp6 point on the severity of labor pain and the delivery outcome. Int. J. Community Based Nurs. Midwifery 2013, 1, 165–172. [Google Scholar]
- Beigi, N.M.A.; Broumandfar, K.; Bahadoran, P.; Abedi, H.A. Women’s experience of pain during childbirth. Iran. J. Nurs. Midwifery Res. 2010, 15, 77–82. [Google Scholar]
- Kolås, T.; Hofoss, D.; Daltveit, A.K.; Nilsen, S.T.; Henriksen, T.; Häger, R.; Ingemarsson, I.; Øian, P. Indications for cesarean deliveries in Norway. Am. J. Obstet. Gynecol. 2003, 188, 864–870. [Google Scholar] [CrossRef]
- Altman, M.; Sandström, A.; Petersson, G.; Frisell, T.; Cnattingius, S.; Stephansson, O. Prolonged second stage of labor is associated with low Apgar score. Eur. J. Epidemiol. 2015, 30, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.W.; Shaffer, B.L.; Nicholson, J.M.; Caughey, A.B. Second Stage of Labor and Epidural Use. Obstet. Gynecol. 2014, 123, 527–535. [Google Scholar] [CrossRef]
- Allen, V.M.; Baskett, T.F.; O’Connell, C.M.; McKeen, D.; Allen, A.C. Maternal and perinatal outcomes with increasing duration of the second stage of labor. Obstet. Gynecol. 2009, 113, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Deussen, A.R.; Ashwood, P.; Martis, R. Analgesia for relief of pain due to uterine cramping/involution after birth. Cochrane Database Syst. Rev. 2011, 11, CD004908. [Google Scholar] [CrossRef] [PubMed]
- Kredatusova, G.; Hajurka, J.; Szakallova, I.; Valencakova, A.; Vojtek, B. Physiological events during parturition and possibilities for improving puppy survival: A review. Vet. Med. 2011, 56, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Zhang, S.; Qiao, Y.; Luo, Z.; Zhang, J.; Zeng, Y.; Wang, L. Predicting pregnancy-induced hypertension with dynamic hemodynamics. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 117, 162–168. [Google Scholar] [CrossRef]
- Reynolds, F. The effects of maternal labour analgesia on the fetus. Best Pract. Res. Clin. Obstet. Gynaecol. 2010, 24, 289–302. [Google Scholar] [CrossRef]
- Singer, D. Neonatal tolerance to hypoxia: A comparative-physiological approach. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1999, 123, 221–234. [Google Scholar] [CrossRef]
- Muns, R.; Nuntapaitoon, M.; Tummaruk, P. Non-infectious causes of pre-weaning mortality in piglets. Livest. Sci. 2016, 184, 46–57. [Google Scholar] [CrossRef]
- Münnich, A.; Küchenmeister, U. Causes, diagnosis and therapy of common diseases in neonatal puppies in the first days of life: Cornerstones of practical approach. Reprod. Domest. Anim. 2014, 49, 64–74. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Orozco, H.; Villanueva, D.; Bonilla, H.; Suárez, X.; Hernandez, R.; Roldan, P.; Trujillo, M. Foetal and neonatal energy metabolism in pigs and humans: A review. Vet. Med. 2011, 56, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Villanueva-García, D.; Velázquez-Armenta, E.Y.; Nava-Ocampo, A.A.; Ramírez-Necoechea, R.; Alonso-Spilsbury, M.; Trujillo, M.E. Influence of time at which oxytocin is administered during labor on uterine activity and perinatal death in pigs. Biol. Res. 2007, 40, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Martinez-Burnes, J.; Alonso-Spilsbury, M.L.; Lopez, A.; Ramirez-Necoechea, R.; Trujillo-Ortega, M.E.; Medina-Hernandez, F.J.; Cruz, N.I.; Albores-Torres, V.; Loredo-Osti, J. Meconium staining of the skin and meconium aspiration in porcine intrapartum stillbirths. Livest. Sci. 2006, 102, 155–162. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Martínez-Burnes, J.; Trujillo, M.E.; López, A.; Rosales, A.M.; Ramírez, R.; Orozco, H.; Merino, A.; Alonso-Spilsbury, M. Uterine and fetal asphyxia monitoring in parturient sows treated with oxytocin. Anim. Reprod. Sci. 2005, 86, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Nava-Ocampo, A.A.; Trujillo, M.E.; Velázquez-Armenta, Y.; Ramírez-Necoechea, R.; Martínez-Burnes, J.; Alonso-Spilsbury, Y.M. Dose minimization study of oxytocin in early labor in sows: Uterine activity and fetal outcome. Reprod. Toxicol. 2005, 20, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Spilsbury, M.; Mota-Rojas, D.; Martínez-Burnes, J.; Arch, E.; López Mayagoitia, A.; Ramírez-Necoechea, R.; Olmos, A.; Trujillo, M.E. Use of oxytocin in penned sows and its effect on fetal intra-partum asphyxia. Anim. Reprod. Sci. 2004, 84, 157–167. [Google Scholar] [CrossRef]
- Tummaruk, P. Post-parturient Disorders and Backfat Loss in Tropical Sows in Relation to Backfat Thickness before Farrowing and Postpartum Intravenous Supportive Treatment. Asian-Australas. J. Anim. Sci. 2013, 26, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Muns, R.; Manzanilla, E.G.; Sol, C.; Manteca, X.; Gasa, J. Piglet behavior as a measure of vitality and its influence on piglet survival and growth during lactation. J. Anim. Sci. 2013, 91, 1838–1843. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Spilsbury, M.; Mota-Rojas, D.; Villanueva-García, D.; Martínez-Burnes, J.; Orozco, H.; Ramírez-Necoechea, R.; Mayagoitia, A.L.; Trujillo, M.E. Perinatal asphyxia pathophysiology in pig and human: A review. Anim. Reprod. Sci. 2005, 90, 1–30. [Google Scholar] [CrossRef]
- Martínez-Burnes, J.; Mota-Rojas, D.; Villanueva-García, D.; Ibarra-Rios, D.; Lezama-García, K.; Barrios-García, H.; Lopez, A. Meconium aspiration syndrome in mammals. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- Edwards, S. Perinatal mortality in the pig: Environmental or physiological solutions? Livest. Prod. Sci. 2002, 78, 3–12. [Google Scholar] [CrossRef]
- Kalantaridou, S.; Makrigiannakis, A.; Zoumakis, E.; Chrousos, G. Stress and the female reproductive system. J. Reprod. Immunol. 2004, 62, 61–68. [Google Scholar] [CrossRef]
- Lezama-García, K.; Orihuela, A.; Olmos-Hernández, A.; Reyes-Long, S.; Mota-Rojas, D. Facial expressions and emotions in domestic animals. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–12. [Google Scholar] [CrossRef]
- van Gelder, K.N.; Bilkei, G. The course of acute-phase proteins and serum cortisol in mastitis metritis agalactia (MMA) of the sow and sow performance. Tijdschr. Diergeneeskd. 2005, 130, 38–41. [Google Scholar] [PubMed]
- Alexopoulos, J.G.; Lines, D.S.; Hallett, S.; Plush, K.J. A review of success factors for piglet fostering in lactation. Animals 2018, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Kushnir, Y.; Epstein, A. Anesthesia for the pregnant cat and dog. Isr. J. Vet. Med. 2012, 67, 19–23. [Google Scholar]
- Radbrook, C.A.; Clark, L. State of the art analgesia- recent developments in pharmacological approaches to acute pain management in dogs and cats. Part 1. Vet. J. 2018, 238, 76–82. [Google Scholar] [CrossRef]
- Bai, Y.H.; Pak, S.C.; Choi, B.C.; Wilson, L. Tocolytic effect of morphine via increased metabolic clearance of oxytocin in the baboon. Yonsei Med. J. 2002, 43, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Arici, G.; Karsli, B.; Kayacan, N.; Akar, M. The effects of bupivacaine, ropivacaine and mepivacaine on the contractility of rat myometrium. Int. J. Obstet. Anesth. 2004, 13, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Castagnetti, C.; Mariella, J. Anti-inflammatory drugs in equine neonatal medicine. Part I: Nonsteroidal anti-inflammatory drugs. J. Equine Vet. Sci. 2015, 35, 475–480. [Google Scholar] [CrossRef]
- Kukkar, A.; Bali, A.; Singh, N.; Jaggi, A.S. Implications and mechanism of action of gabapentin in neuropathic pain. Arch. Pharm. Res. 2013, 36, 237–251. [Google Scholar] [CrossRef]
- Elikottil, J.; Gupta, P.; Gupta, K. The analgesic potential of cannabinoids. J. Opioid Manag. 2009, 5, 341–357. [Google Scholar] [CrossRef]
- Bonar, M.; Hance Ellington, E.; Lewis, K.P.; Wal, E. Implementing a novel movement-based approach to inferring parturition and neonate caribou calf survival. PLoS ONE 2018, 13, e192204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, P.; Garel, M.; Morellet, N.; Benoit, L.; Chaval, Y.; Itty, C.; Petit, E.; Cargnelutti, B.; Hewison, A.J.M.; Loison, A. A standardised biologging approach to infer parturition: An application in large herbivores across the hider-follower continuum. Methods Ecol. Evol. 2021, 12, 1017–1030. [Google Scholar] [CrossRef]
- Zheng, T.; Moustafa, Y.; Finn, C.; Scott, S.; Haase, C.J.; Carpinelli, N.A.; Osorio, J.S.; McKinstry, K.K.; Strutt, T.M.; Huo, Q. A rapid blood test to monitor immunity shift during pregnancy and potential application for animal health management. Sensors Int. 2020, 1, 100009. [Google Scholar] [CrossRef]


| Factor That Increases Labour Pain | References |
|---|---|
| Maternal characteristics: Prolonged labours
| [62,108,113] |
| Factor That Increases Labour Pain | References |
|---|---|
| Maternal characteristics: Duration of farrowing (more than 3 h):
| [1,19,116,117,120] |
| Responses | Variables and Scales | References |
|---|---|---|
| Sympathetic-Adreno-Medullary System |
| [135,138] |
| Hypothalamic Pituitary Adrenocortical System |
| [135,136,139] |
| Species | Variables and Scales | References |
|---|---|---|
| Women | (a) McGill Pain Questionnaire [MPQ], (b) The Short-Form MPQ, (c) Visual analogue scales [VAS]. Verbal rating scales, or simple ordinal scales. | [5,134,141] |
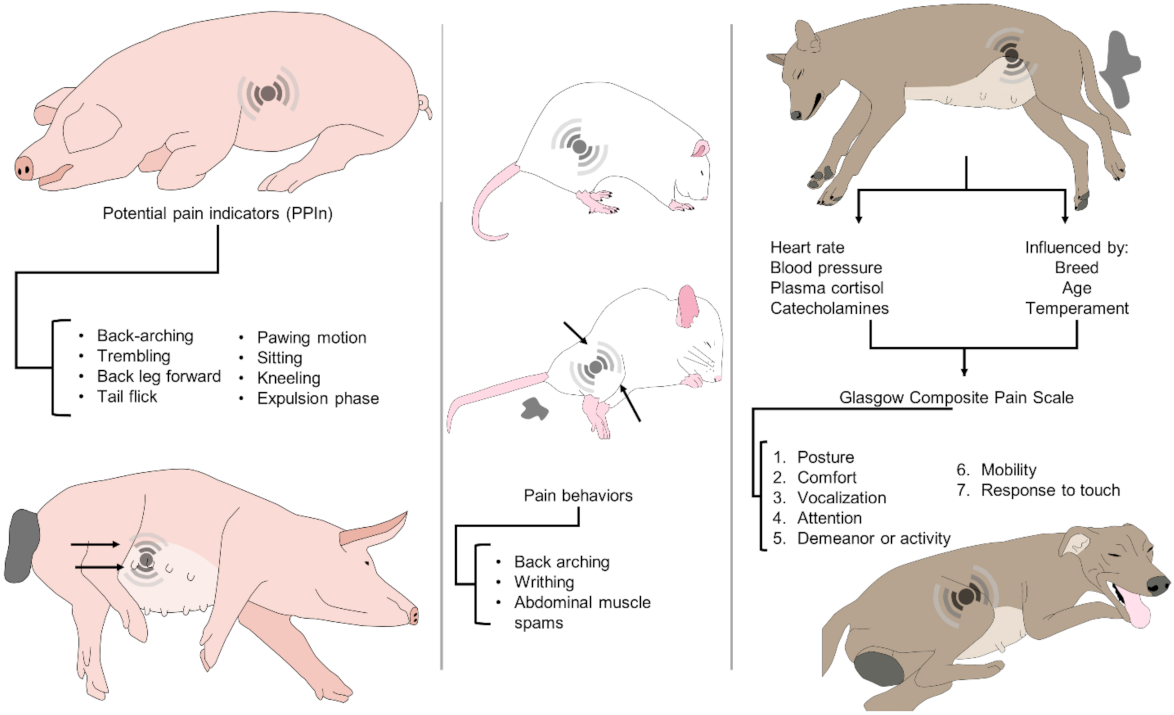
| Dog | The behavioural representation of pain is species-specific and affected by age, breed, temperament, and additional stressors, including anxiety or fear; hence, it is necessary to identify the normal behaviour of the bitch. The composite measure pain scale (CMPS) has been described for use in dogs with acute pain, based on seven categories: (1) posture, (2) comfort, (3) vocalization, (4) attention to wound, (5) demeanour, (6) mobility, and (7) response to touch. | [16,23,133,142,144] |
| Rat | Activity of spinal neurons that receive afferents, using immunodetection of c-Fos protein, an indirect indicator of harmfully activated spinal neurons. Labour induced the expression of neuronal c-Fos in segments T12-S2 of the spinal cord one hour following the delivery of the first pup. Automated systems for the detection of pain behavior. General activities, including food and water consumption, rearing (upright exploration), and chest and head grooming, are evaluated to determine spontaneous behaviour during labour. Phase stretching behaviour during labour included: crushing (an asymmetrical contraction of the lower body and limbs), sidelong contraction (an asymmetric contraction of the lower body and limbs), lengthening (stretching of the abdomen and the four limbs) and phasic humpback posture. Maternal care activities included building nests, licking pups, and eating the placenta. | [15,145,146] |
| Sow | Numerical pain score per animal is calculated by evaluating behavioural, clinical, and physiological patterns in a specific term. Observation of certain behaviours as potential pain indicators (PPIn) in sows before, during and after farrowing: hind leg forward (in a side-lying posture, the back leg is pulled ahead and/or in towards the body), back arch (in a side-lying position, one or both legs turn rigid and are pushed aside from the body and/or inwards towards the centre, creating a curvature in the back). Putative indicators of pain used to evaluate the pain behaviour in the periparturient sow. Also, it coincides with an activity shift from nest buildings to passivity, an increase of the myometrial electrical activity, and the increment of oxytocin levels before the beginning of the ejection of piglets. Postural changes considered: stand (upright, with all feet on the floor), sit (front legs straight and back end down on the floor), lie lateral (lying on one side with udder exposed), kneel (front knees on the ground, with hinds legs straight), lie ventral (lying with the udder on the floor), tremble (visibly shaking as if shivering when in a lateral lying position), and others; tail-flick (tail is moved rapidly up and down), paw (in a lateral lying position, the front leg scraped in a pawing motion), piglet delivery (piglet completely ejected from the dam). | [3,74,147,148] |
| Species | Route | Drug | Observations | References |
|---|---|---|---|---|
| Women | Inhalational analgesia (reduce pain perception). | 1. Entonox. 2. Isoflurane. 3. Desflurane. 4. Sevoflurane. | Use limited to developed countries. | [13,21,150] |
| Parenteral opioids (reduce pain perception). | 1. Pethidine. 2. Morphine. 3. Diamorphine. 4. Fentanyl. 5. Remifentanil. 6. Meperidine. | Adverse effects have been observed. | [13,21,150,153,154,155,156,157,158,159] | |
| Regional analgesia (reduce pain transmission). | 1. Epidural. 2. Combined spinal-epidural (CSE). Lidocaine. Xilacina. Levobupivacaine. Bupivacaine. | Epidural analgesia is the “gold standard” for labour analgesia. | [13,153,158,160,161] | |
| Bitch | Intravenous Intramuscular. | 1. Opioids (Methadone, Morphine) 2. AINEs (Meloxicam). | Use limited to caesarean section. | [162,163] |
| Rat | Intrathecal. | Morphine. | Used in research model for labour pain. | [146] |
| Sow | Intramuscular, Oral. | Ketoprofen. Meloxicam. | [3,164,165] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Burnes, J.; Muns, R.; Barrios-García, H.; Villanueva-García, D.; Domínguez-Oliva, A.; Mota-Rojas, D. Parturition in Mammals: Animal Models, Pain and Distress. Animals 2021, 11, 2960. https://doi.org/10.3390/ani11102960
Martínez-Burnes J, Muns R, Barrios-García H, Villanueva-García D, Domínguez-Oliva A, Mota-Rojas D. Parturition in Mammals: Animal Models, Pain and Distress. Animals. 2021; 11(10):2960. https://doi.org/10.3390/ani11102960
Chicago/Turabian StyleMartínez-Burnes, Julio, Ramon Muns, Hugo Barrios-García, Dina Villanueva-García, Adriana Domínguez-Oliva, and Daniel Mota-Rojas. 2021. "Parturition in Mammals: Animal Models, Pain and Distress" Animals 11, no. 10: 2960. https://doi.org/10.3390/ani11102960
APA StyleMartínez-Burnes, J., Muns, R., Barrios-García, H., Villanueva-García, D., Domínguez-Oliva, A., & Mota-Rojas, D. (2021). Parturition in Mammals: Animal Models, Pain and Distress. Animals, 11(10), 2960. https://doi.org/10.3390/ani11102960