1. Introduction
Mastitis remains a widely spread disease in dairy production. Current prevention and cure methods for mastitis are mainly antibiotic-based. The administration of antibiotics might cause the emergence of drug-resistant pathogens. Thus, alternative solutions for mastitis have long been necessary [
1]. The epithelial barrier system has an important immunological function, which prevents commensal and pathogenic microorganism infection. Particularly, epithelial innate immunity serves as the first line of defense against pathogens and has the potential to act coordinately with acquired immunity.
Antimicrobial peptides/proteins (AMPs) are one of the potent players of epithelial innate immunity, having a lower potential to cause antibiotic-resistant pathogens [
2]. Therefore, AMPs in bovine mammary glands are thought to be a good target for the development of mastitis prevention and treatment methods. The type of AMPs differ among species; e.g., bovines do not possess α-defensin, a potent mucosal AMP in humans and mice [
3]. This might imply that cows have a unique AMP system when compared with humans and rodents. Thus, revealing potent AMPs in the bovine mammary gland is necessary.
Chemerin, which was originally identified as a chemoattractant protein for antigen-presenting cells [
4], is currently recognized to have antimicrobial properties [
5]. This protein has an inhibitory effect on commensal pathogen activity, including
Escherichia coli and
Staphylococcus aureus which are the major mastitis-causing pathogens [
6,
7]. Taken together, chemerin has the potential to be a pivotal player of immunity against epithelial infection and thereby maintain tissue homeostasis. As we previously reported that
retinoic acid receptor responder 2 (
RARRES2), encoding chemerin was highly expressed in cows’ mammary glands [
8], we hypothesized that chemerin might act to regulate the mammary epithelial barrier mechanism. Studies including ours demonstrated that inflammatory cytokines strongly induce the expression of chemerin, which implies that immune response during mastitis pathogenesis or involution process might regulate chemerin signaling in the bovine mammary gland [
9,
10]. Therefore, the objectives of the present study were to identify chemerin-producing cells in mammary glands, and investigate the regulatory factor of its production, to understand its role in the mammary epithelial defense system.
2. Materials and Methods
2.1. Animals and Sample Collection
For gene expression evaluation of chemerin and its receptors, mammary gland and fresh milk samples were collected from multipara Holstein dairy cows in the lactating or dry-off period (n = 5 in total). For investigation of gene expressional change of chemerin, mammary gland and liver tissue were immediately separated from euthanized cows at 1 week (i.e., during the early-lactation stage; n = 3), 5 months (i.e., at the mid-lactation stage; n = 4), 9 months (i.e., at the late lactation stage; n = 3), and 15 months (i.e., at the dry-off period; n = 3) post parturition. The cows used in this experiment were multiparous (experiencing 3.1 ± 1.4 calving), except for two that were primiparous at the mid-lactation stage. All cows at late lactation were pregnant, whereas others were not. Euthanasia was performed by exsanguination from the carotid artery after a captive bolt or an overdose administration of sodium pentobarbital (Kyoritsu Seiyaku, Tokyo, Japan) via the jugular vein. Thereafter, all tissue samples were collected and frozen immediately in liquid nitrogen and stored at −80 °C for PCR and immunoblotting analyses or were fixed in phosphate-buffered 10% formalin for immunostaining. The other two multiparous Holstein dairy cows, in the dry-off period, were employed for the isolation and culture of peripheral blood mononuclear cells (PBMCs). Whole blood samples (20 mL) were drawn via jugular vein puncture using sodium heparin (10 IU/mL) (BD Biosciences, Billerica, MA, USA) as an anticoagulant. Blood samples were stored at room temperature and transferred to the laboratory for separation of PBMC.
2.2. Culture of Bovine Mammary Epithelial Cells
Immortalized bovine mammary epithelial cells (MAC-T cells) were cultured according to a previously described protocol [
8]. Briefly, subculturing was performed in Low-Glucose Dulbecco’s Modified Eagle Medium (DMEM) (Wako Pure Chem, Osaka, Japan) supplemented with 10% fetal bovine serum (FBS) (GE Healthcare, Little Chalfont, UK), 1% penicillin-streptomycin-amphotericin B solution (Wako Pure Chem), 5 µg/mL bovine insulin (Sigma-Aldrich, St. Louis, MO, USA), and 1 µg/mL progesterone (Sigma-Aldrich) on a 100 mm plastic culture dish (Corning, Corning, NY, USA). MAC-T cells were induced for differentiation prior to cytokine treatment. Confluent MAC-T cells cultured in 12-well plates (Corning) were differentiated for 5 d in DMEM supplemented with 10% FBS, 1% penicillin-streptomycin-amphotericin B solution, 5 µg/mL bovine insulin, 10 µg/mL dexamethasone (Sigma-Aldrich), 5 µg/mL ovine prolactin (Sigma-Aldrich), and human growth hormone (Wako Pure Chem). Cells were cultured at 37 °C in a humidified CO
2 incubator (5% CO
2).
2.3. Cell Proliferation Assay
The proliferation of MAC-T cells was assessed by the 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay. MAC-T cells were seeded at 1000 cells/well in 96-well culture plates, and cultured in DMEM supplemented with 10% FBS and a 1% penicillin-streptomycin-amphotericin B suspension (Wako Pure Chem) for 10 h at 37 °C in a humidified CO2 incubator (5% CO2). Thereafter, the medium was removed and MAC-T cells were incubated with DMEM supplemented with 10% FBS, a 1% antibiotic mixture, and recombinant human chemerin (R&D Systems, Minneapolis, MN, USA) at 10, 100, or 300 ng/mL for 24 h at 37 °C, under 5% CO2. Following incubation, the MTT solution (5 mg/mL in 1× PBS) was added to each cell, followed by another 4 h incubation at 37 °C, 5% CO2. The supernatant was removed and 100 µL/well of 0.04 N HCl/isopropanol was added to dissolve the MTT formazan precipitate. An optical density (OD) measurement was performed at 570 nm with a DTX-880 microplate reader (Beckman Coulter, Brea, CA, USA).
2.4. Measurement of Extracellular Lactate Dehydrogenase (LDH) Level
MAC-T cells were cultured using the same procedure for the MTT assay and treated with chemerin at 10, 100, or 300 ng/mL for 24 h. Conditioned medium (50 μL) from each well was transferred to a new 96-well plate. Assay buffer was prepared as follows: mixing buffer A (4 mM iodonitrotetrazolium chloride in 0.2 M Tris-HCl, pH8.2), buffer B (6.4 mM nicotinamide adenine dinucleo-tide sodium salt, 320 mM lithium lactate, in 0.2 M Tris-HCl buffer), and buffer C (150 mM 1-methoxyphenazinemethosulfate in Tris buffer) at a ratio of 5000:5000:1. Each sample of conditioned medium was mixed with 50 μL of assay buffer and incubated for 1 h at 25 °C. Fifty microliters of 1 molar acetic acid was added to each well and the OD measurement was performed at 490 nm.
2.5. Measurement of Transepithelial Electrical Resistance (TEER)
The effect of chemerin on the integrity of MAC-T cells was investigated by monitoring the TEER across cell monolayers. MAC-T cells were seeded on cell culture inserts (Corning) and cultured until the required confluency was reached. Thereafter, the medium in the culture insert was replaced with a fresh subculturing medium containing 0.6 M H2O2 with/without recombinant human chemerin (10 or 100 ng/mL; R&D Systems) to incubate for 3 h. Cell culturing and incubation were conducted at 37 °C, 5% CO2, and 95% relative humidity. After incubation with H2O2 with/without chemerin, cell culturing plates with inserts were taken outside the CO2 incubator and a Millicell Electrical Resistance system (Millipore, Burlington, MA, USA) connected to electrodes was used to measure TEER values. Control experiments were also conducted under the same experimental conditions.
2.6. Isolation of Peripheral Blood Mononuclear Cells (PBMCs) and Measurement of Cell Proliferation
Isolation and cell proliferation assay of PBMCs were performed as previously described [
11]. Briefly, whole blood samples were processed within 6 h of sample collection. PBMCs were separated by density gradient centrifugation; whole blood was diluted with 1× PBS and layered gently over Histopaque-1077 (Sigma-Aldrich). Isolated PBMCs were washed twice with 1× PBS. The PBMCs obtained were diluted with serum-free RPMI-1640 medium (Sigma-Aldrich) and the cell numbers were determined. The viability of PBMCs typically exceeded 85%, determined by trypan blue staining. The PBMCs were resuspended at 1 × 10
6 viable cells/mL in Roswell Park Memorial Institute (RPMI) 1640 medium containing 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) supplemented with heat-inactivated FBS, 2 mM L-glutamine, 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B (all supplements were from Sigma-Aldrich) in 96-well plates. The PBMCs were incubated at 37 °C under 5% CO
2 for 24 and 48 h with or without chemerin (100 ng/mL). Thereafter, cell proliferation was analyzed by the Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technology, Kumamoto, Japan) according to the manufacturer’s protocol.
2.7. Cytokine Treatment of Bovine Mammary Epithelial Cells
Confluent MAC-T cells were washed twice with prewarmed 1× PBS and incubated in DMEM containing 10% FBS and a 1% penicillin-streptomycin-amphotericin B suspension supplemented with recombinant mouse tumor necrosis factor alpha (TNF-α) (1, 10 or 100 ng/mL; R&D Systems), recombinant human interleukin beta 1 (IL-β1) (0.5, 5, or 50 ng/mL; R&D Systems), or recombinant human INF-γ (1, 10, or 100 ng/mL; R&D Systems) for 24 h in a CO2 incubator. Treated cells were collected for gene expression analysis.
2.8. Gene Expression Analysis
Total RNA was extracted using RNAiso Plus (Takara Bio, Shiga, Japan). The purity and integrity of the extracted RNA were checked using a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and agarose gel electrophoresis, respectively. First-strand cDNA was synthesized from 500 ng of total RNA using the PrimeScript RT Reagent Kit with the gDNA Eraser Perfect Real Time (Takara Bio), while no-input RNA reaction served as a negative control. RT-PCR for gene expression was performed using GoTaq Green Master Mix (Promega, Madison, WI, USA), followed by agarose gel electrophoresis stained by ethidium bromide. Quantitative RT-PCR (qRT-PCR), to investigate gene expressional levels of
RARRES2, chemerin receptor genes (
chemokine-like receptor 1; CMKLR1, C-C motif chemokine receptor like 2; CCRL2, G protein-coupled receptor 1; GPR1), and other genes related to mammary gland function, was performed using SYBR Premix Ex Taq II Tli RNaseH Plus (Takara Bio) with the Thermal Cycler Dice Real Time System II (Takara Bio). Reactions with no-input RNA or without cDNA served as negative controls. The primer sequences are shown in
Table S1. The PCR efficiencies were checked by constructing standard curves using 5 points serial dilution of pooled cDNA. The reference genes for expressional level analysis were chosen from 18S rRNA, cyclin G-associated kinase (
GAK), actin-beta (
ACTB), or vacuolar protein sorting 4 homolog A (
VPS4A) for each experiment based on their stability in the value of threshold cycle (Ct) among animal groups. Gene expression levels were calculated using the 2
−ddCt method and described as relative values to the control group.
2.9. Immunoblotting
Plasma samples were diluted 10 times with PBS. Frozen mammary tissue samples or cultured MAC-T cells were homogenized in radioimmunoprecipitation assay buffer supplemented with 1 mM Na
3VO
4 and a protease inhibitor cocktail (Nacalai Tesque, Kyoto, Japan). Fresh milk samples were centrifuged at 18,000×
g for 30 min and the intermediate clear phase was transferred to a new 1.5 mL tube. This was repeated once more to obtain soluble protein from fresh milk. To analyze chemerin abundance in mammary glands with and without mastitis, the soluble protein was extracted from paraffin-embedded tissue sections according to a published protocol [
12]. A total of 10 µg of these protein samples or 10 µL of diluted plasma were separated by 15% SDS-PAGE and electroblotted onto a PVDF membrane. The membranes were blocked using 3% bovine serum albumin (BSA) in Tris-buffered saline and Tween 20 (TBST) for 30 min. Membranes were incubated with primary antibodies overnight at 4 °C, followed by incubation with secondary antibodies for 1 h at 25 °C. The antibodies used for Western blotting were as follows: anti-bovine chemerin (custom-made rabbit polyclonal IgG raised against antigen peptide; CVTSVDNAADTLFPAGQF, purified by antigen affinity chromatography), anti-β-actin (sc47778; Santa Cruz Biotechnology, Dallas, TX, USA), anti-rabbit IgG horseradish peroxidase (HRP)-conjugated (W4011; Promega), and anti-mouse IgG HRP-conjugated (W4021; Promega). Detection of the probed band was achieved using an enhanced chemiluminescence (ECL) Prime Western Blotting Detection System and ImageQuant LAS 4000 (GE Healthcare).
2.10. Immunohistochemistry (IHC)
Frozen mammary gland tissues from Holstein dairy cows were sectioned to a 10 μm thickness, air-dried for 30 min at 25 °C, and fixed with 10% formalin for 20 min. Alternatively, formalin-fixed paraffin-embedded (FFPE) mammary glands were sectioned, deparaffinized with xylene, and rehydrated with a graded series of ethanol. The FFPE blocks of normal and mastitic tissue were adapted from a previous study [
13] using different animals from ones used for gene expression analysis or frozen sections mentioned in
Section 2.1. The mammary gland tissues for FFPE were obtained from cows with healthy or naturally occurring mastitic udder, euthanized in a local slaughterhouse. FFPE tissue sections were subjected to antigen retrieval by autoclave treatment in citrate buffer (pH 6.0), followed by incubation in 1% peroxide diluted in methanol to inactivate endogenous peroxidase. Thereafter, frozen or FFPE tissue sections were blocked with blocking buffer (2% normal goat serum diluted in TBST). Sections were incubated with anti-bovine chemerin rabbit IgG (custom-made polyclonal antibody: described above), anti-human CMKLR1 rabbit IgG (ab82773; Abcam, Cambridge, UK), or anti-human CCRL2 rabbit IgG (LS-A1094; LifeSpan Biosciences, Seattle, WA, USA) diluted in Can Get Signal Immunostain (Toyobo, Osaka, Japan) at 4 °C overnight. Following washing in TBST, tissue sections were probed with diluted Goat Anti-Rabbit IgG H&L conjugated with Alexa Fluor 488 or HRP at 25 °C for 1 h. The HRP-labeled immunocomplex was visualized using the diaminobenzidine substrate. Sections were observed by fluorescent or bright-field microscopy. To evaluate the specificity of the custom-made chemerin antibody, polyclonal IgG was adsorbed by an antigen-immobilized sepharose column, and the column flow-through was used as a negative control.
2.11. Statistical Analysis
Numerical data from experiments are presented as means ± standard error (SE). Data were statistically analyzed using student’s t-test or one-way ANOVA followed by Tukey-Kramer test, depending on the number of experimental groups. Significant differences were declared when
p < 0.05, and tendencies were declared when
p < 0.10. All statistical analyses were performed using the statistical package R (v3.4.4; available as a free download from
https://www.r-project.org, accessed on 8 November 2021).
4. Discussion
The present study examined the potential role of chemerin in the bovine mammary gland, and also assessed its involvement in mastitis, considering its role as a chemokine [
4,
15], and its recently recognized antimicrobial potential [
6]. The results of the expressional analysis demonstrated gene and protein expression of chemerin in mammary glands of dairy cows and its milk secretion. Chemerin was originally reported to be abundantly present in the inflammatory fluid and to have chemotactic potential to dendritic cells, macrophages, and natural killer (NK) cells, whereas its effect on monocytes is in deliberation [
4]. Antigen-presenting cells, particularly macrophages, reside in healthy ma-mmary glands, which contribute to the monitoring of bacterial invasion via alveoli epithelial cells. The present results show basal production of chemerin in mammary epithelial cells, suggesting its contribution to the local innate immunity of mammary glands by recruiting these immune cells. Chemerin is an antimicrobial agent in the human epidermis and inhibits the growth of commensal pathogens including E. coli and S. aureus, which are known as major mastitis-causing pathogens. This property is attributed to the hydrophobic internal region of chemerin [
6,
7]. Another study reported the production of chemerin in intestinal epithelia of human fetuses and infants [
16]. Thus, these reports indicate that chemerin produced in epithelial tissues has antimicrobial potential in addition to chemotactic ability, which is similar to complement factors or some specific types of chemokine [
17]. Importantly, despite a lack of putative apical-sorting signal in its amino acid sequence, chemerin was secreted from both apical and basolateral sides in a study using an intestinal cell line [
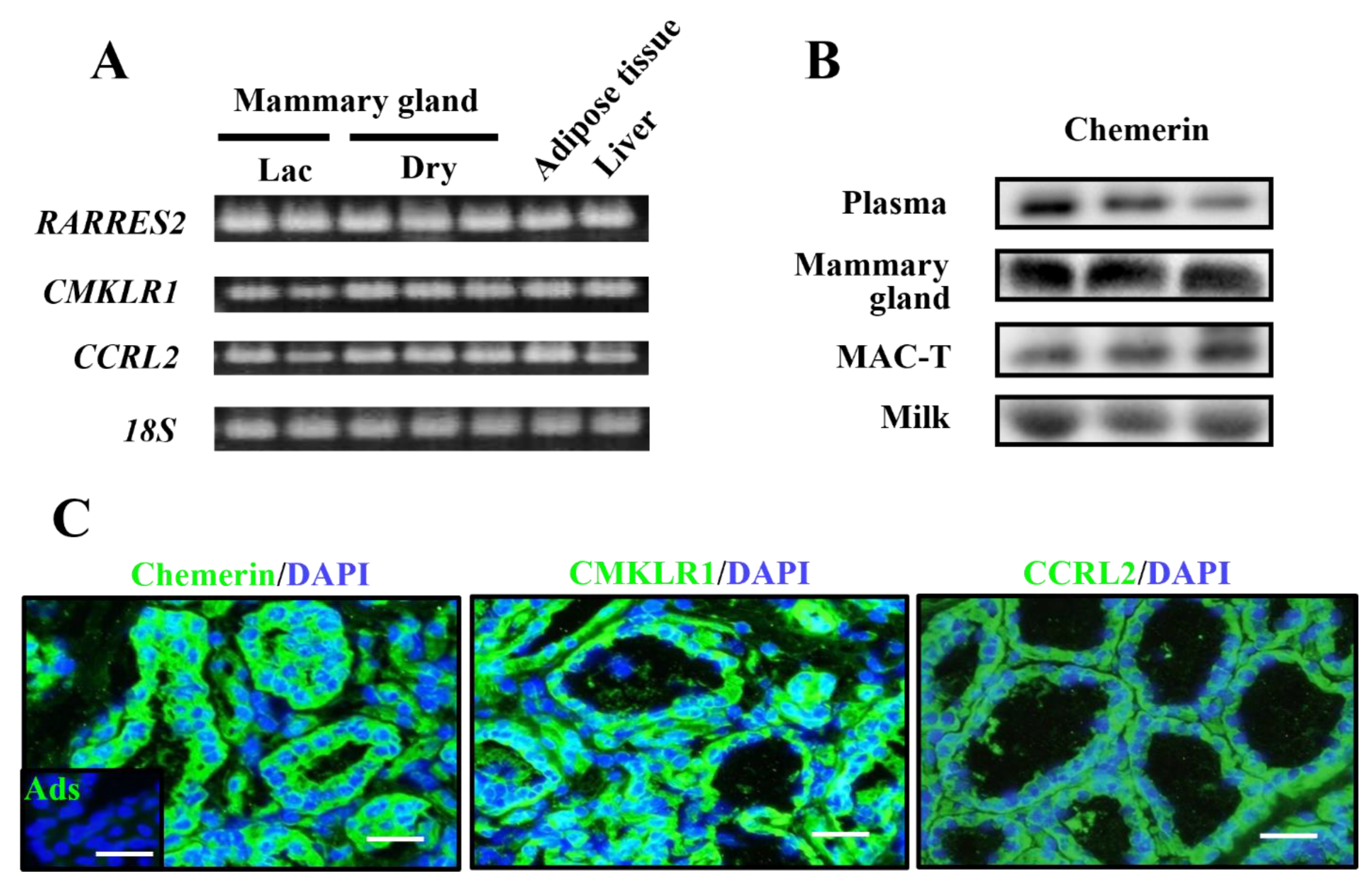
16]. This supports our data showing the detectable amount of secreted chemerin in milk. Thus, it is thought that chemerin potentially acts as an antimicrobial protein and a chemokine in a mammary epithelial barrier mechanism in dairy cows.
In addition to the reported functions of chemerin related to immunity, we previously revealed that chemerin upregulated GLUT1 and CSN3 (kappa-casein) expression in cultured mammary epithelial cells [
8], suggesting its positive effect on mammary gland functions including lactation. As chemerin was shown to be produced in the cow’s mammary gland, this protein can act in a paracrine as well as endocrine manner. This led us to further investigate if chemerin has another regulatory effect on mammary epithelial cells. Already, some research exists indicating the proliferative effect of chemerin on some cell types, including vascular smooth muscle cells, and preadipocytes [
18,
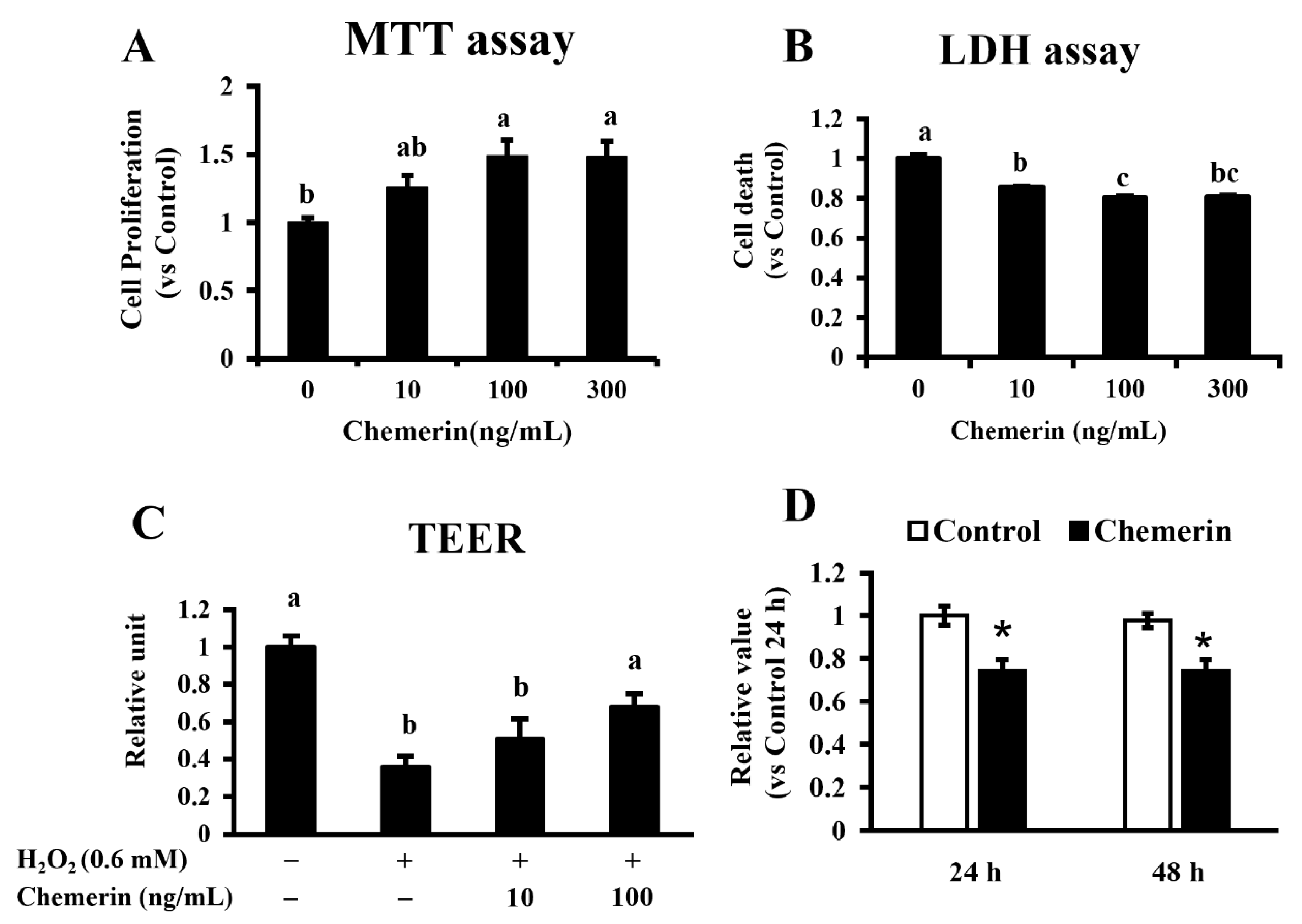
19]. In the present study, the treatment of chemerin was shown to promote the proliferation and suppressed LDH release in MAC-T cells. Our previous study found that chemerin elevates intracellular ERK activity in MAC-T cells, supporting the present result. This suggests the promot-ive effect of chemerin on the growth and survival of mammary epithelial cells.
The mammary barrier function is largely attributed to epithelial integrity, mediated by tight junctions. In the case of mastitis, epithelial integrity is impaired by inflammatory stimuli of molecules such as bacterial lipopolysaccharide (LPS), inflammatory cytokines, and reactive oxygen species (ROS) [
20]. We treated MAC-T cells with H
2O
2 to mimic ROS-induced damage in mastitis and evaluated the effect of chemerin in this process. The results showed that chemerin protected MAC-T cells from H
2O
2 induced epithelial disintegration. It is reported that elevated ROS and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) activity in mammary epithelial cells impairs cellular polarity, which is necessary for epithelial integrity [
21]. A previous study showed that chemerin inhibits NF-κB activity induced by inflammatory cytokine stimuli [
22]. This supports our result of the protective effect of chemerin on the epithelial barrier. The result that chemerin suppressed proliferation of PBMCs might simultaneously suppress the overactivity of immune response to maintain epithelial integrity. However, a previous report also indicated that chemerin activates the PI3K/Akt pathway, which might be involved in the dysfunction of epithelial integrity [
18]. Thus, the mechanism of chemerin on epithelial integrity requires careful investigation in future studies. Therefore, our results demonstrate the potential protective effect of chemerin on bovine mammary glands by preventing impaired epithelial integrity.
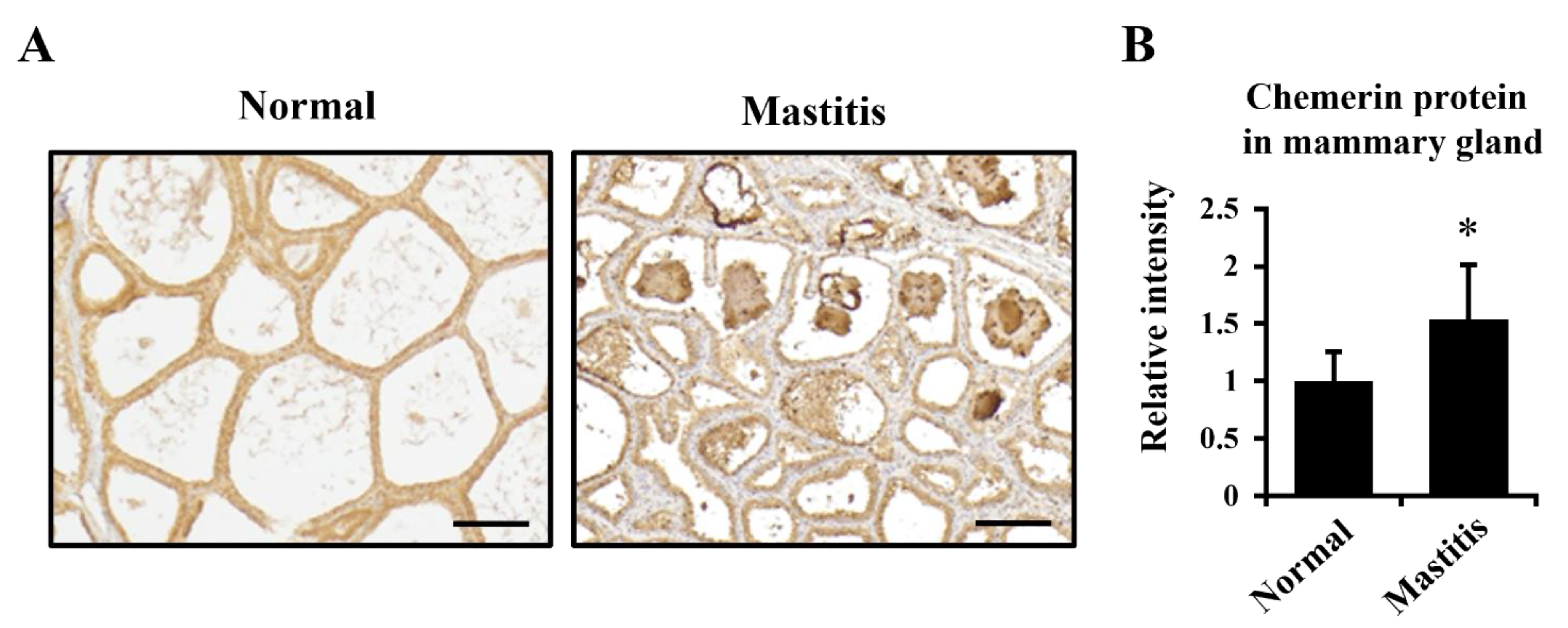
The present study suggested the role of chemerin in the inflammatory response in mammary glands because its production was upregulated in mastitis. Whereas the recruitment of neutrophils takes place acutely during mastitis, the simultaneous infiltration of macrophages is thought to occur in mammary epithelia [
23]. A previous study demonstrated that the stimulation of bovine mammary glands with LPS, lipoteichoic acid, and/or peptidoglycan induced upregulation of monocyte chemotactic protein-1 (MCP-1) production, a well-known chemokine for macrophages [
24]. Thus, chemerin might be involved in this step with its chemotactic activity. Alternatively, it is also reported that proteolytic activation of chemerin is performed by
S. aureus-derived serine proteases, as well as by cathepsins as host proteases, which are necessary for developing its chemotactic activity [
7]. This suggests a synergistic activation of chemerin by host and bacterial proteases to lead to the acute inflammatory response in the infected/damaged region of mammary tissue.
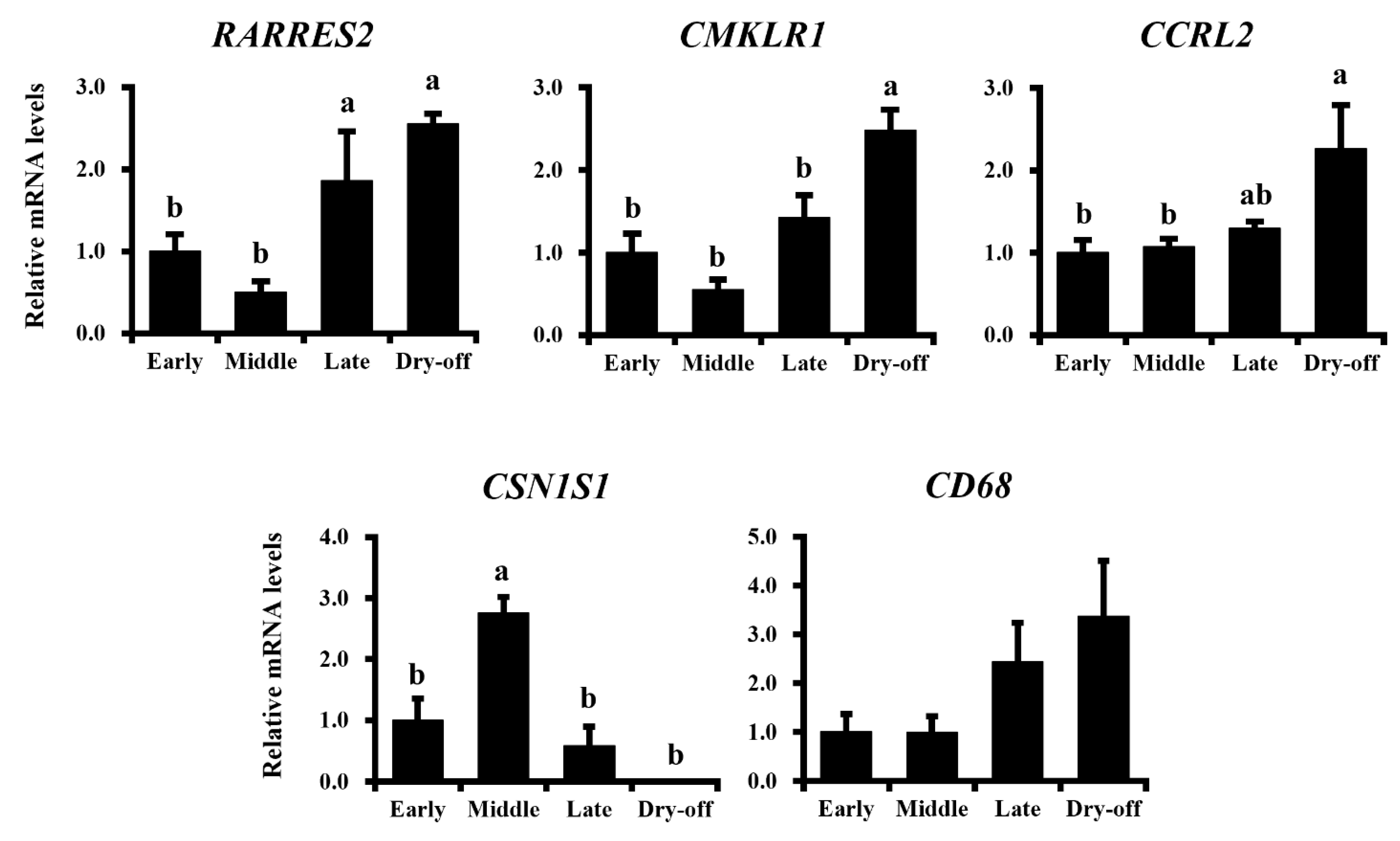
Our data demonstrated higher expression of chemerin in involuted mammary glands of non-pregnant dry-off cows. Following the onset of involution, secreted matrix metalloproteases activate chemokines to recruit immune cells into mammary glands for the depletion of apoptotic mammary epithelial cells. During this process, macrophages are recruited to remove apoptotic cells, resulting in the occupation of stromal cells in tissue [
25]. Additionally, Hu et al. reported that chemerin induced the activities of autophagy and apoptosis in cultured bovine mammary epithelial cells (BMECs) [
26]. The chemerin-induced apoptosis was further activated when autophagy was inhibited by chloroquine. This implicates that chemerin can induce the programmed death of bovine mammary epithelial cells during the process of involution and maintenance of the mammary gland during the dry-off period [
26]. Therefore, chemerin was suggested to act as a chemokine to antigen-presenting cells (APCs), and also as an inducer of apoptosis in involuting/involuted mammary glands, which supports the closed relationship between chemerin and the immune response in mammary glands. However, the difference in the direct effect of chemerin on bovine mammary epithelial cells observed in the study indicated above and the present study, promoting cell death or survival, might be dependent on lactation period and hormonal status, and must be investigated in detail in a future study.
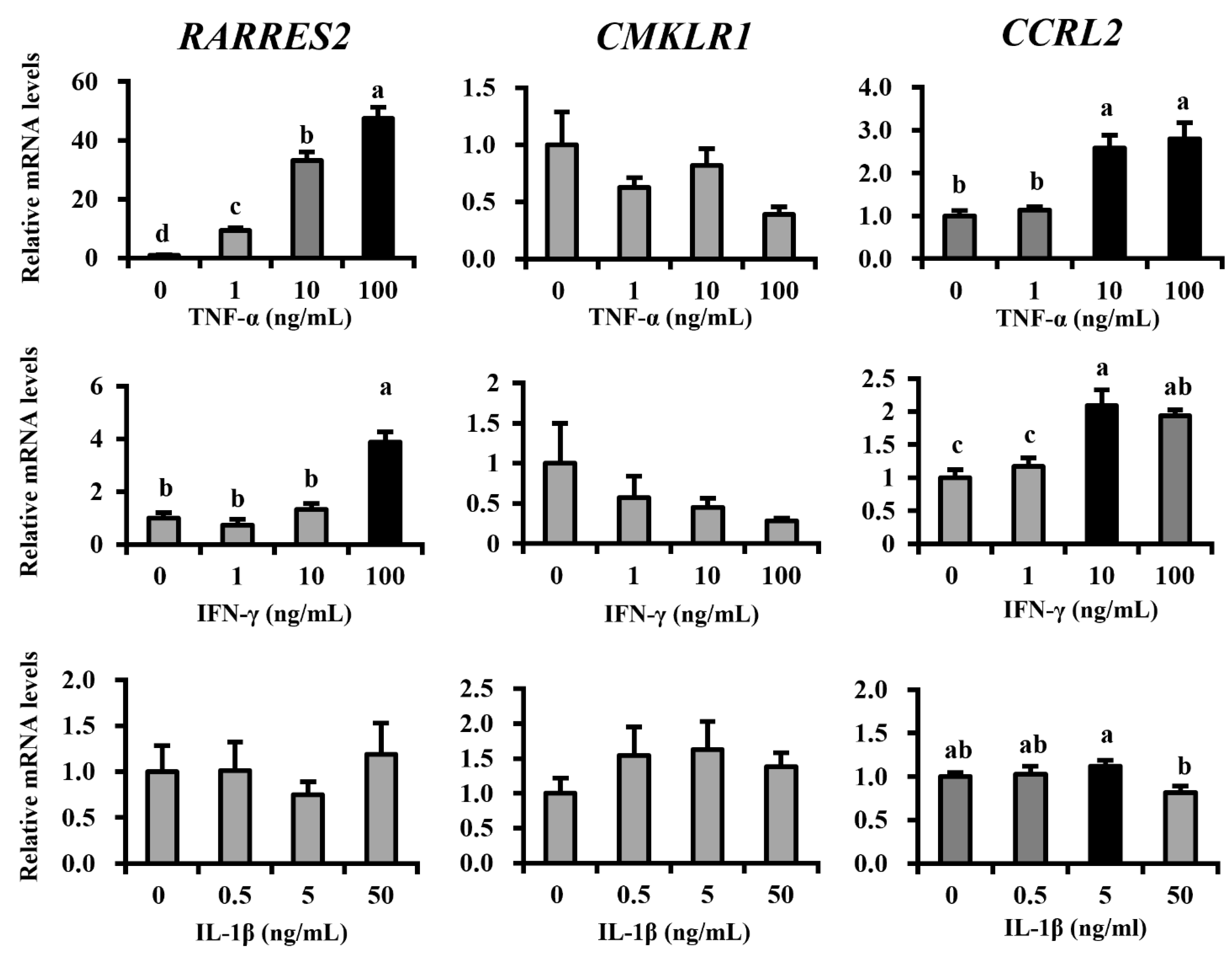
Here, we further tried to determine the regulatory factors of mammary epithelial chemerin expression. During the process of mastitis pathogenesis, the production of pro-inflammatory cytokines is acutely induced to activate immune responses: firstly IL-8 attracts neutrophils, followed by TNF-α, interferon gamma (IFN-γ), and IL-1β induces tissue inflammation [
27]. Our results showed that TNF-α strongly upregulated chemerin expression, as well as high-dose INF-γ. The effect of TNF-α on chemerin upregulation was consistent with the previous studies, including ours, using various cell types: epithelial cells, fibroblasts, and adipocytes [
28]. Meanwhile, the response of chemerin receptor expression for these cytokines varied between each receptor. The expression of
CMKLR1, a functional receptor for chemotaxis induction, was not affected by all cytokines used to treat MAC-T cells. Future studies should investigate its expression in APCs.
CCRL2 expression was upregulated by TNF-α and IFN-γ. CCRL2 was reported to capture chemerin on the cell surface for local concentration in supporting chemotaxis [
29]. Thus, this upregulation was beneficial for APC recruitment in inflammatory conditions in mammary glands. It could be concluded that TNF-α, of the assessed cytokines, is the most potent upregulator of chemerin in mastitis, which contributes to APC recruitment to the inflammatory site, in combination with upregulated
CCRL2.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




