1. Introduction
Coccidiosis causes tremendous economic losses in broiler production by impairing gut health and depressing growth performance and feed efficiency of broiler chickens, and expensive anti-coccidial treatments also increase the overall production cost [
1,
2]. Avian coccidiosis are induced by
Eimeria spp., which are protozoan parasites, and there are 7 known
Eimeria spp. that can infect chickens:
Eimeria acervulina,
E. maxima,
E. tenella,
E. brunetti,
E. necatrix,
E. mitis, and
E. praecox [
3]. Each species resides at the different section of the intestinal tract of broiler chickens, and thereby it has different modes of actions to affect growth performance and gut health of broilers [
4]. Teng et al. [
5] reported that
E. maxima decreased digestibility of crude proteins and amino acids in broilers.
Eimeria spp. can be transmitted via the fecal–oral route. The infection is initiated by ingestion of sporulated (infectious) oocysts, and after the asexual and sexual replications, un-sporulated oocysts are excreted with feces [
6]. In an appropriate environment, the oocysts can be sporulated and become infectious, and this life cycle can be repeated with poultry growth cycle.
Eimeria tenella (
E. tenella) resides in the mucus membrane of ceca, and during its replications, epithelial cells in ceca are damaged, resulting in hemorrhagic diarrhea and impaired growth performance and intestinal health in broilers [
7]. The ceca, the main intestinal compartment for bacterial fermentation, can be reservoirs for pathogenic bacteria and their toxins that can cause oxidative stress after entering the blood stream of broilers [
8]. However, ceca also play crucial roles in producing beneficial bacterial metabolites including vitamins, volatile fatty acids (VFA), lactic acid, and antimicrobial compounds via bacterial fermentation [
9]. The VFA are not only inhibit the growth of pathogenic bacteria, but also are energy substrates for the host and induce gut development of chickens by accelerating gut epithelial cell proliferation [
10]. Moreover, VFA interact with fat metabolism via mitogen-activated protein kinase (MAPK) pathway [
11]. These suggest that VFA are closely associated with feed efficiency by providing extra energy to the host or influencing metabolism of chickens.
To cope with
Eimeria spp. infection in broiler production, anti-coccidial drugs, and vaccination has been used in the broiler industry. However, the use of anti-coccidial drugs has been restricted by inhibiting the use of old anti-coccidial drugs and requiring Veterinary Feed Directives (VFD) registrations because of the spread of resistant
Eimeria strain and consumer pressure [
12,
13]. Furthermore, vaccination is expensive and can prevent spread of
Eimeria spp. [
14]. Recently, a lot of attention has been paid to find nutritional interventions to control
Eimeria spp. infection in broilers. Diverse bioactive compounds, including essential oils [
15], probiotics [
16], sodium butyrate [
17], and plant extracts [
18], were studied to control or to ameliorate negative effects of
E. tenella infection and in broilers. The modes of actions of those bioactive compounds may include damaging cell wall of
E. tenella, modulating cecal microbiota, and/or enhancing the immunity of broilers. To find suitable nutritional interventions, it is important to understand mode of actions of
E. tenella on the growth of chickens and to set up appropriate experimental infection models to test novel nutritional interventions. Therefore, the hypothesis of this study was that impaired cecal health due to
E. tenella infection may result in reduced growth performance and impaired intestinal health because of reduced VFA production and increased oxidative stress. The purpose of the study was to investigate the effects of different inoculation dosages of
E. tenella on growth performance, gastrointestinal permeability, oocyst shedding, fecal consistency, intestinal morphology, ileal apparent digestibility, antioxidant capacity, and cecal VFA in broiler chickens.
4. Discussion
The purpose of the study was to investigate the effects of different inoculation dosages of
E. tenella on growth performance, gastrointestinal permeability, oocyst shedding, intestinal morphology, fecal consistency, ileal apparent digestibility, antioxidant capacity, and cecal VFA profile in broiler chickens. Inoculation dosages were derived from our previous study [
19], and the same strain of
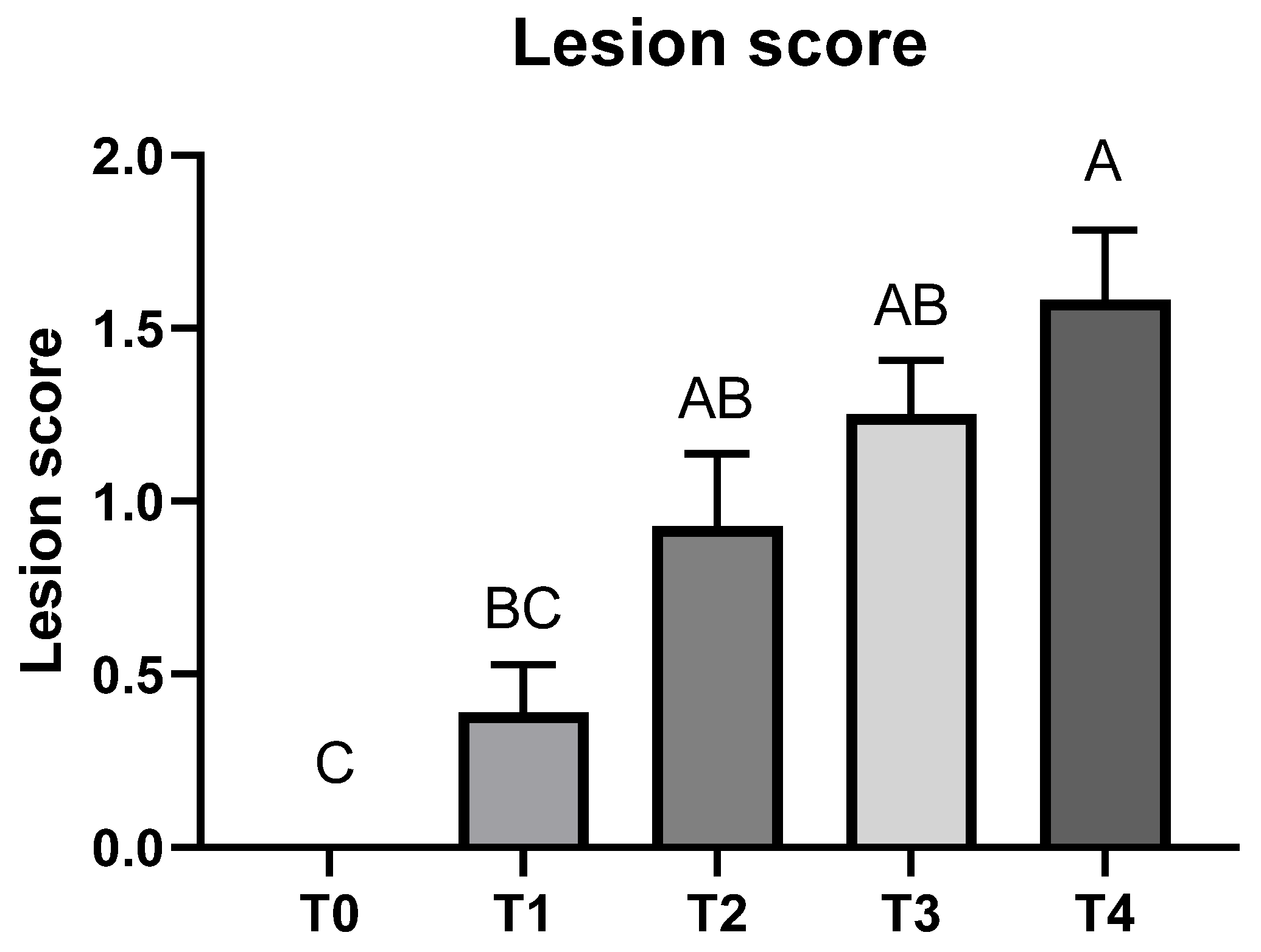
E. tenella was used. However, in the current study, milder infection (lesion score of T4: 1.6) was achieved compared to our previous study (lesion score of the T4 equivalent group: 2.7). The potential reason for milder infection would be that broilers were challenged with three different
Eimeria spp. (
E. acervulina,
E. maxima, and
E. tenella) in the previous study, which probably caused more severe infection by compensating the immune system in birds compared to the single species challenging in the current study. Mild-infection models are important to test a new bioactive compound because if the compounds do not show any beneficial effects at the mild infection model, they may not show any beneficial effects against coccidiosis at severe infection model either. In the present study,
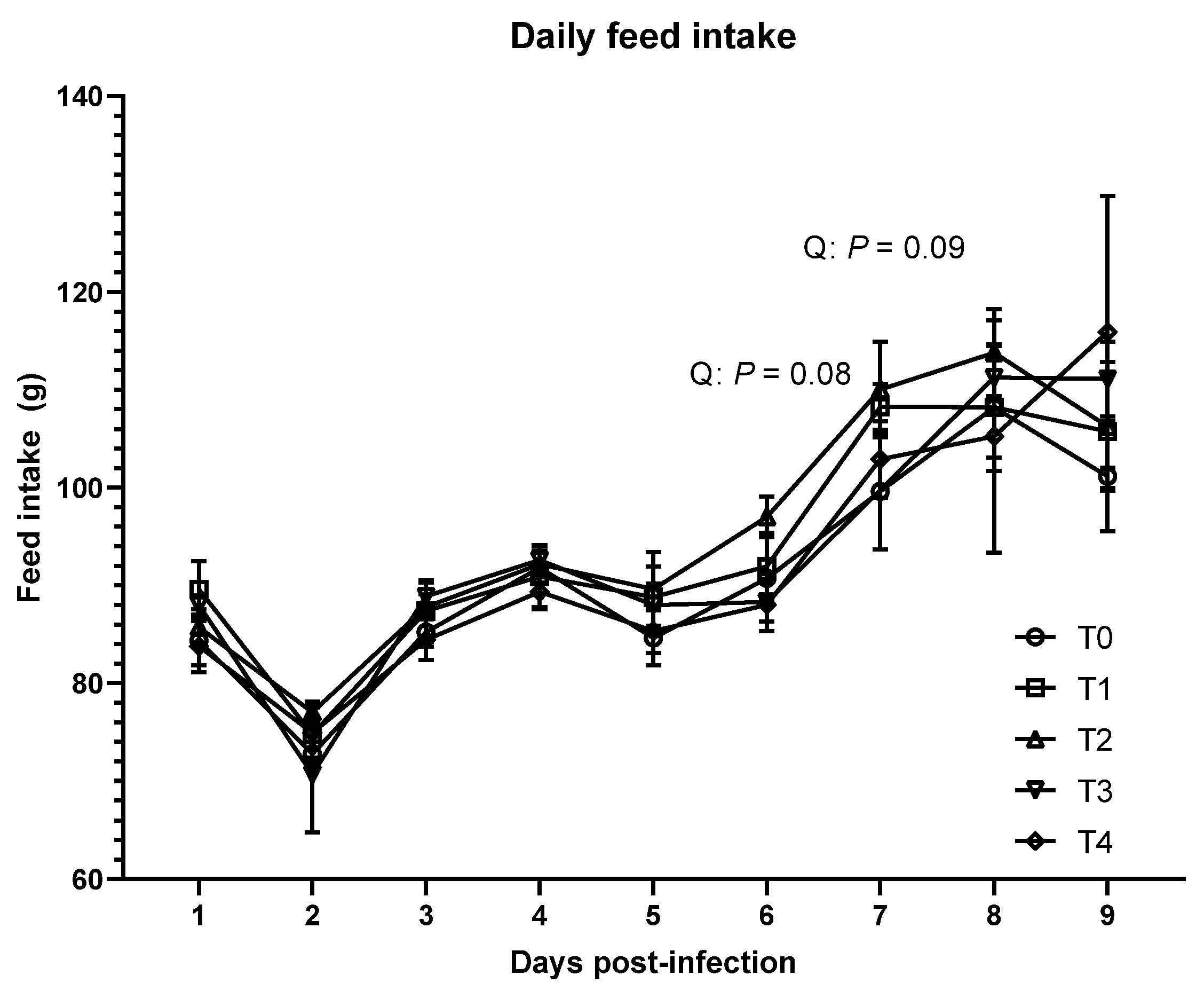
E. tenella infection decreased feed efficiency during the acute phase and increased feed intake on 6 and 7 dpi. These data are partially consistent with previous studies [
24,
25] which reported that FCR was increased along with reduced BW of broilers. In our current study, however,
E. tenella infection increased FCR by increasing the feed intake of birds, whereas BW was also numerically (
p = 0.16) decreased with a linear trend during the acute phase. Impaired feed efficiency in broilers infected with
E. tenella during the acute phase in the present study would be associated with reduced acetate and total VFA production in the ceca. Acetate is the most abundant VFA, which are produced via bacterial fermentation in the ceca of broilers [
26]. It is already well-established that
E. tenella infection can negatively affect cecal microbiome, which results in depressed VFA production in broilers according to many previous studies [
27,
28]. Cecal VFA production affects the host’s energy balance because VFA are important energy substrates for the host [
29]. Gasaway [
30] mentioned that VFA supply approximately 11% to 18% of the total energy production in chickens. In addition,
Eimeria spp. compete for energy and nutrients for their asexual and sexual replications with the host [
31]. Reduced available energy in the body can result in reduced BW or increased FCR by raising feed intake of the birds to decrease maintenance energy requirements or to meet the energy requirements, respectively [
32]. In addition, reduced production of acetate, which can induce the secretion of satiety-stimulating hormones from the gut, could increase the feed intake of the birds [
33]. Therefore, in this study, reduced VFA production in ceca due to
E. tenella infection potentially reduced available energy levels in the body, and this increased feed intake and FCR to supply more energy to meet the energy requirements in broiler chickens infected with
E. tenella.
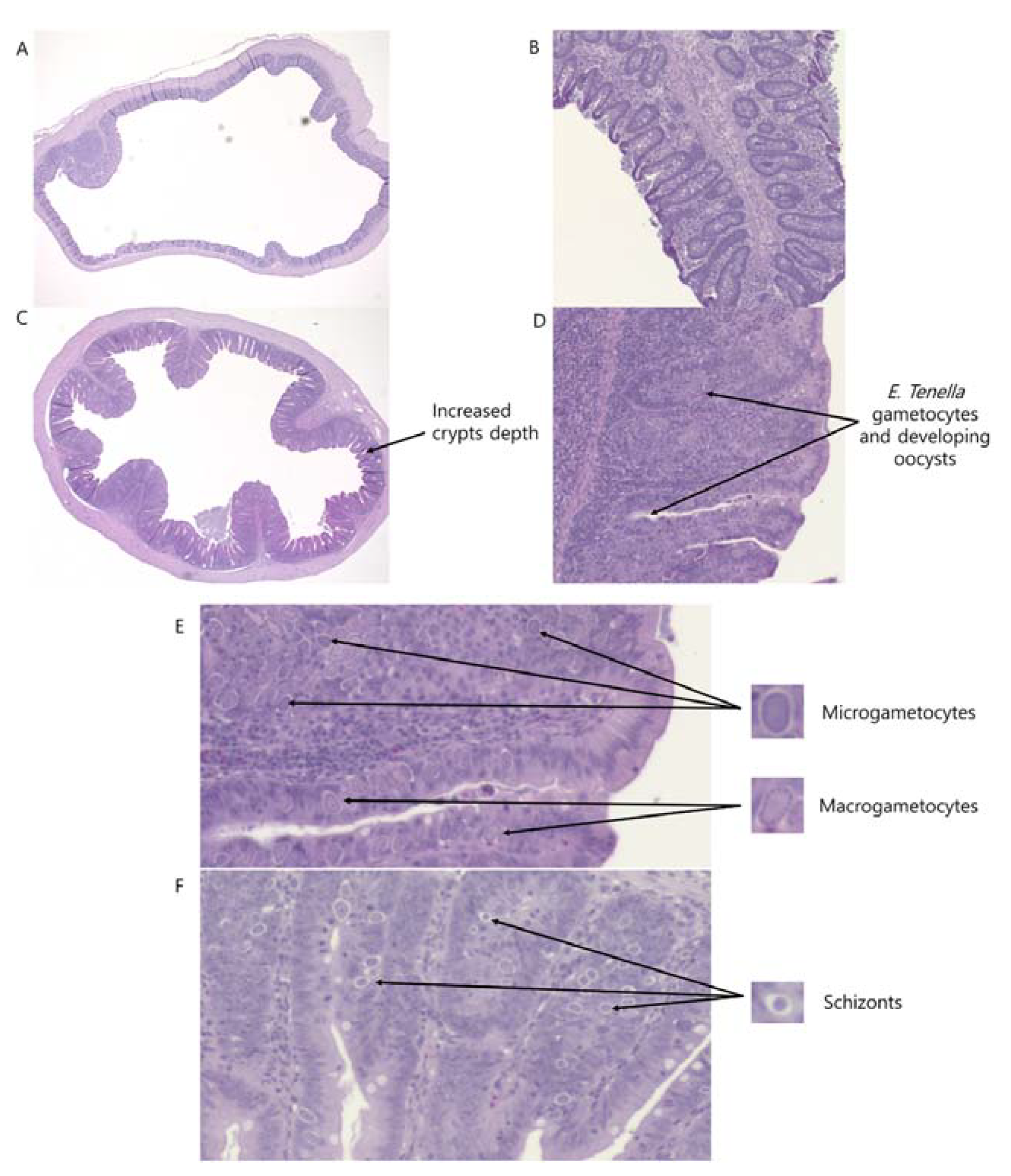
This study showed that
E. tenella infection deepened crypts depth of the ceca. The potential reasons for increased cecal crypts depth due to
E. tenella infection are still unclear. It is proposed that
E. tenella increased cecal CD (mucosa layer) to make their habitats in the ceca, or ceca crypts were deepened to increase VFA absorption because VFA production was restricted due to
E. tenella infection. Nevertheless, deepened CD could inhibit the production and absorption of VFA in the ceca. Increased CD possibly turned in increased total goblet cells in the ceca and mucus secretion into the cecal contents. This possibly reduced concentration of VFA and modulated VFA production in ceca content by affecting microbiome of the broiler chickens. Ceca only have villi at the entrance of the ceca to filter large particles away and to act as an immunological detector of cecal contents [
34,
35], and middle and distal parts of the ceca only have smooth mucous membrane without villi [
36]. Thereby, another possible reason for increased CD in broilers infected with
E. tenella would be that birds increased CD to let crypts function like villi as a defensive mechanism in the proximal ceca because
E. tenella infected ceca are vulnerable for further infections (e.g., bacterial infection).
We also hypothesized that impaired ceca health due to
E. tenella infection may indirectly affect small intestinal health (the main area for nutrient digestion and absorption) by causing energy deficiency and inducing oxidative stress, and this may account for decreased feed efficiency in the current study. Nevertheless, no differences were observed in DM, OM, and ash apparent ileal digestibility among the treatments on 6 and 9 dpi in the current study. According to our previous study,
E. maxima infection significantly decreased nutrient digestibility in broiler chickens, which indicates that different
Eimeria spp. affect nutrient digestibility of broilers differently [
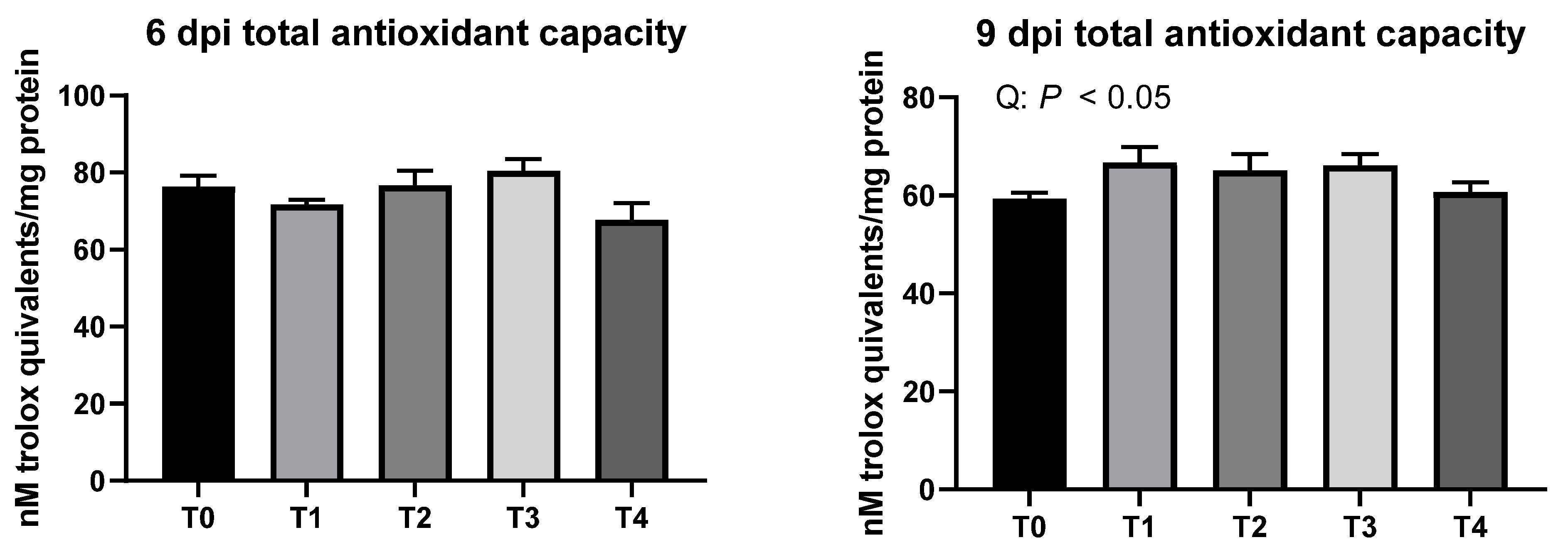
5]. Liver total antioxidant capacity was same among the treatments on 6 dpi and even increased in broilers infected with
E. tenella on 9 dpi, potentially because mild infection of
E. tenella may allow birds to stimulate their antioxidant defensive system. Still, over-production of antioxidants (enzymatic and non-enzymatic) can result in over-use of energy and nutrients, which also may decrease available energy and nutrients for growth in chickens. However, a previous study by Georgieva et al. [
25] reported that severe
E. tenella infection model decreased antioxidant capacity of broilers. Ileal crypt depth was decreased, and ileal VH:CD was increased without affecting VH in
E. tenella infected broilers in the current study. Probably, ileal morphology was enhanced with limited energy and nutrients sources as a compensation mechanism because cecal functionality was restricted due to
E. tenella in the current study. There were negative effects of
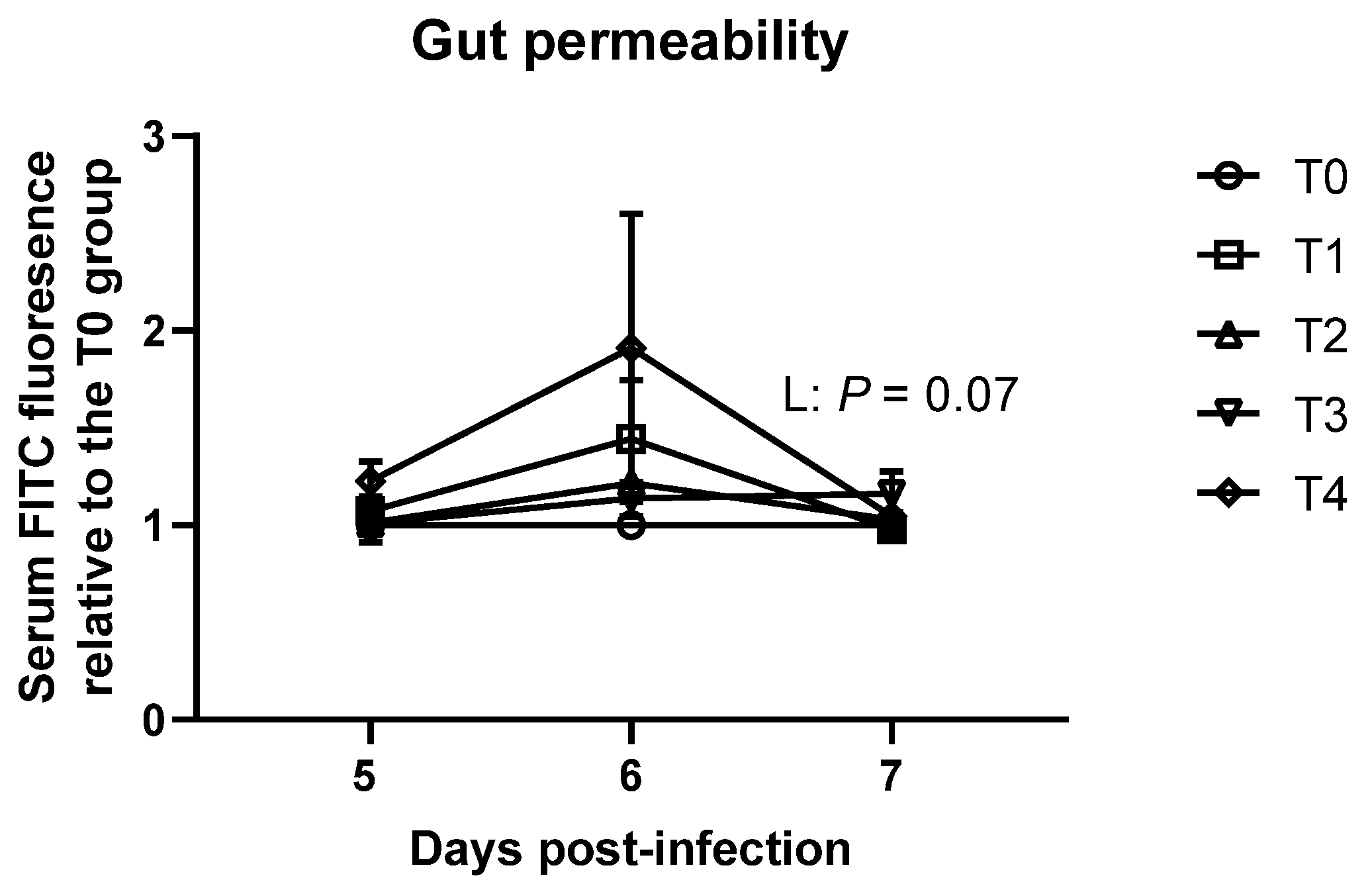
E. tenella on duodenal and jejunal morphology on 9 dpi and gastrointestinal permeability on 7 dpi. Energy and absorbed VFA play an important role in gut development in broiler chickens by stimulating gut epithelial cell proliferation [
9]. In this study, reduced cecal VFA production would be the main factor that negatively affected gut health of
E. tenella infected broilers rather than increased pathogens and toxin production because liver health was maintained in broilers infected with
E. tenella. Whereas it cannot be concluded that impaired intestinal health caused impaired feed efficiency during the acute phase, energy deficiency due to poor VFA yield subsequently damaged intestinal health of broilers infected with
E. tenella. Moreover, while differences were observed in gastrointestinal permeability, it was only less than two-fold difference. Our previous study [
37] reported that more than 200 folds differences of gastrointestinal permeability were observed due to
E. maxima infection. Hence, reduced VFA production in the ceca due to
E. tenella infection caused energy deficiency in the body, which resulted in compensated gut health in broiler chickens.
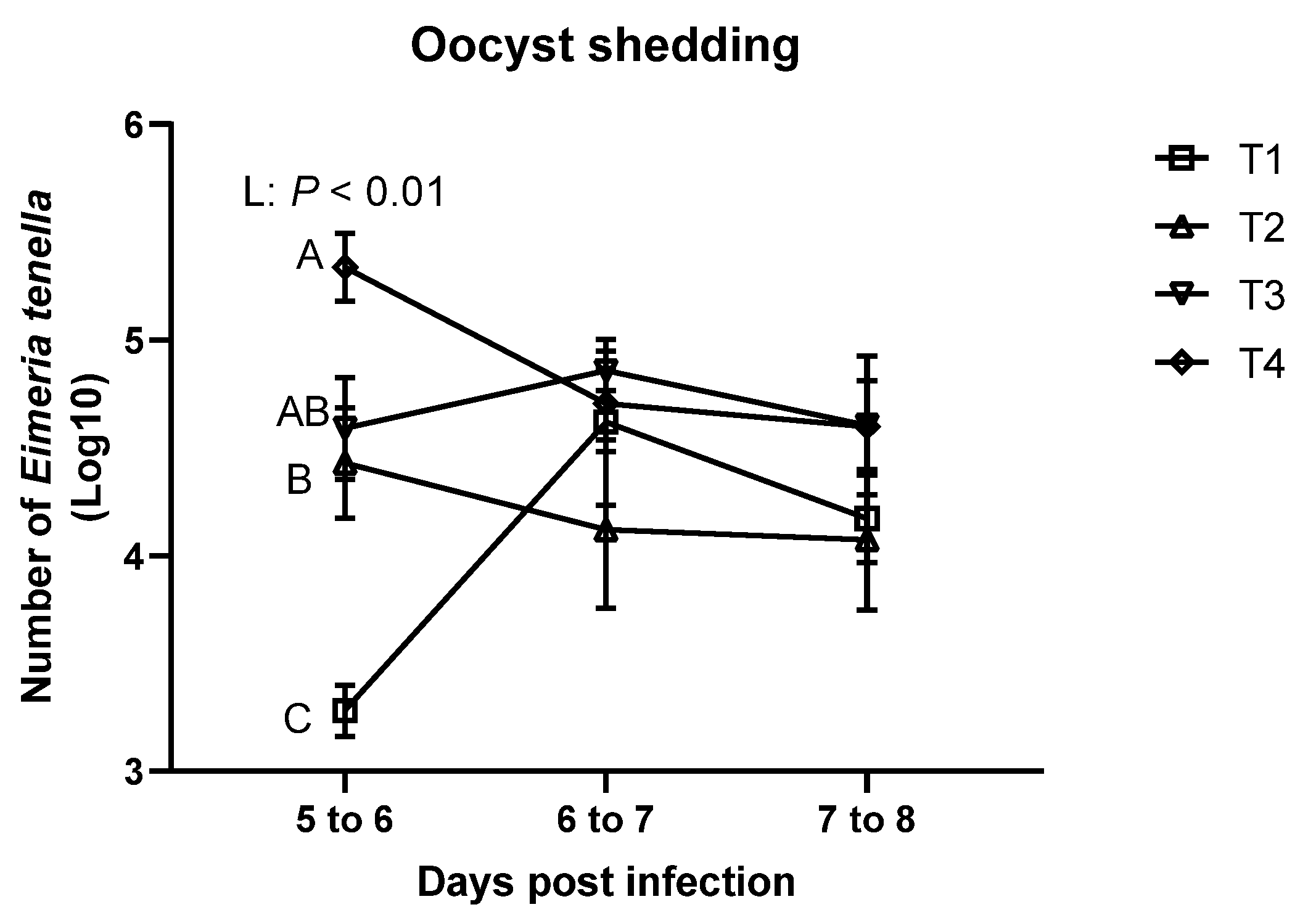
In this study, different
E. tenella inoculation dosages linearly increased oocyst shedding on 5 to 6 dpi, which implies that the higher inoculation dosages can results in higher oocyst shedding. Nonetheless, these data could be obtained in our study because our
E. tenella strain induced mild-infection (lesion score below 2) in the ceca. Williams [
38] demonstrated that higher inoculation dosage levels can linearly increase oocyst yields until reaching to the crowded dosages (e.g., maximally producing dosage), and higher dosages than the crowded dosages can decrease oocyst yields in broilers. However, no differences were observed on 6 to 7 dpi and 7 to 8 dpi in the current study. The current result also demonstrated that different challenge dosages of
E. tenella have different peak point of shedding. The potential reason would be that higher number of
E. tenella possibly decreased number of generation within the asexual and sexual stages of the life [
35], which resulted in an earlier peak date for the highest dosage group in the study. Our study was the first to find that different
E. tenella inoculation dosages resulted in different peak points for oocyst yields in the mildly infected broilers. Oocyst yields in severely infected (
E. tenella lesion score 3 to 4) broilers at different time points should be investigated further.
Ileal and fecal moisture contents were modulated due to Eimeria infection in the current study. Ileal (6 dpi) and fecal moisture contents (5 to 6 dpi) were quadratically and linearly decreased, respectively. Afterwards, fecal moisture content was increased on 6 to 8 dpi.
E. tenella are known to induce diarrhea containing mucus and blood. Although obvious bloody diarrhea was not achieved in the current study, fecal moisture content was increased after 6 dpi. Potentially,
E. tenella damaged enterocytes in the ceca and modulated microbiota, and this caused electrolyte imbalance in the ceca which may explain the modulated ileal and fecal moisture contents in the current study [
39]. Moreover, ceca play a crucial role in water absorption [
40], and water absorption ability of the ceca of birds infected with
E. tenella would be limited potentially due to thickened mucosa layer in the current study. Hypothetically, higher water loss due to
E. tenella infection increased water intake [
40] as birds increased their feed intake to compensate energy deficiency in the current study. In addition, increased fecal moisture content can result in increased litter moisture, and this can increase the incidence of food pad dermatitis in broilers [
41].
This current study showed that mild infection of
E. tenella impaired feed efficiency and gut health mainly through reducing VFA production in broilers. Thereby, supplementation of VFA or bioactive compounds that has high energy values and antimicrobial effects (e.g., medium chain fatty acids) can control mild-infection of
E. tenella in broilers [
11,
42].



 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}