Author Contributions
Conceptualisation, methodology, investigation and visualisation, H.M.G.P.H., S.J.P., P.R.K., H.T.B. and P.C.H.M.; software, H.M.G.P.H.; formal analysis, H.M.G.P.H., S.J.P. and P.C.H.M.; resource, S.J.P., P.R.K., H.T.B. and P.C.H.M.; data curation, H.M.G.P.H., S.J.P. and P.C.H.M.; writing—original draft preparation, H.M.G.P.H.; writing—review and editing, H.M.G.P.H., S.J.P., P.R.K., H.T.B. and P.C.H.M.; supervision, S.J.P., P.R.K., H.T.B. and P.C.H.M.; project administration, S.J.P., P.R.K., H.T.B. and P.C.H.M.; funding acquisition, P.R.K. and H.T.B. All authors have read and agreed to the published version of the manuscript.
Figure 1.
The site (A) and angle (B) of positioning of Ultrasound scanner to capture rumen images.
Figure 1.
The site (A) and angle (B) of positioning of Ultrasound scanner to capture rumen images.
Figure 2.
The length of papillae outer boundary of the stratum corneum (green line) and the respective straight length of rumen tissue (blue line) measured in dorsal and ventral sites of rumen by FIJI ImageJ software, to calculate the ratio between length of papillae surface/straight length of rumen tissue (PSL/STL) of lambs.
Figure 2.
The length of papillae outer boundary of the stratum corneum (green line) and the respective straight length of rumen tissue (blue line) measured in dorsal and ventral sites of rumen by FIJI ImageJ software, to calculate the ratio between length of papillae surface/straight length of rumen tissue (PSL/STL) of lambs.
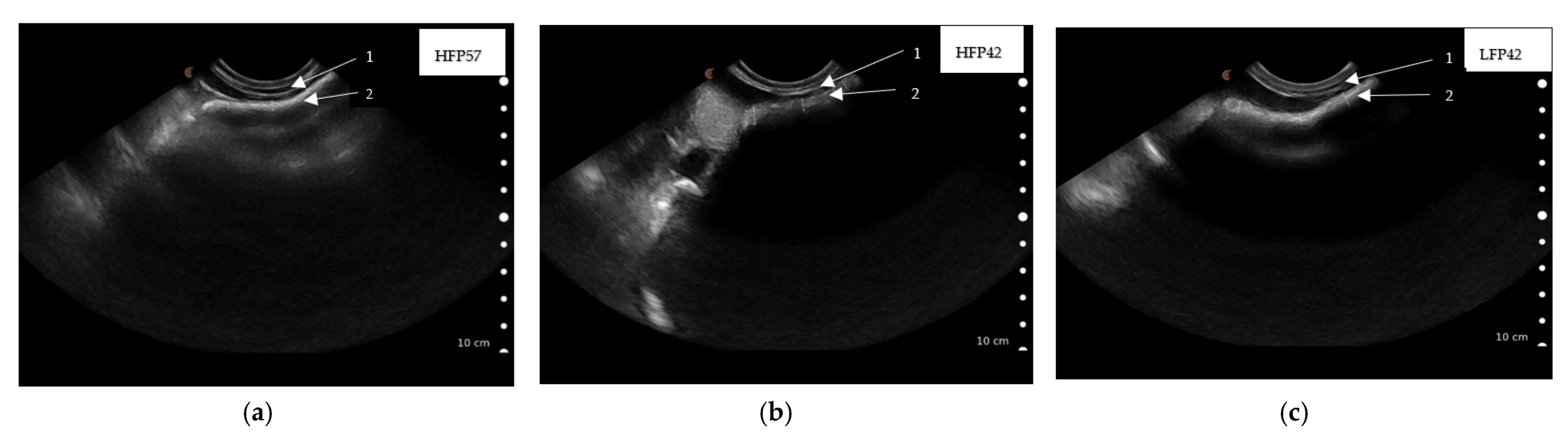
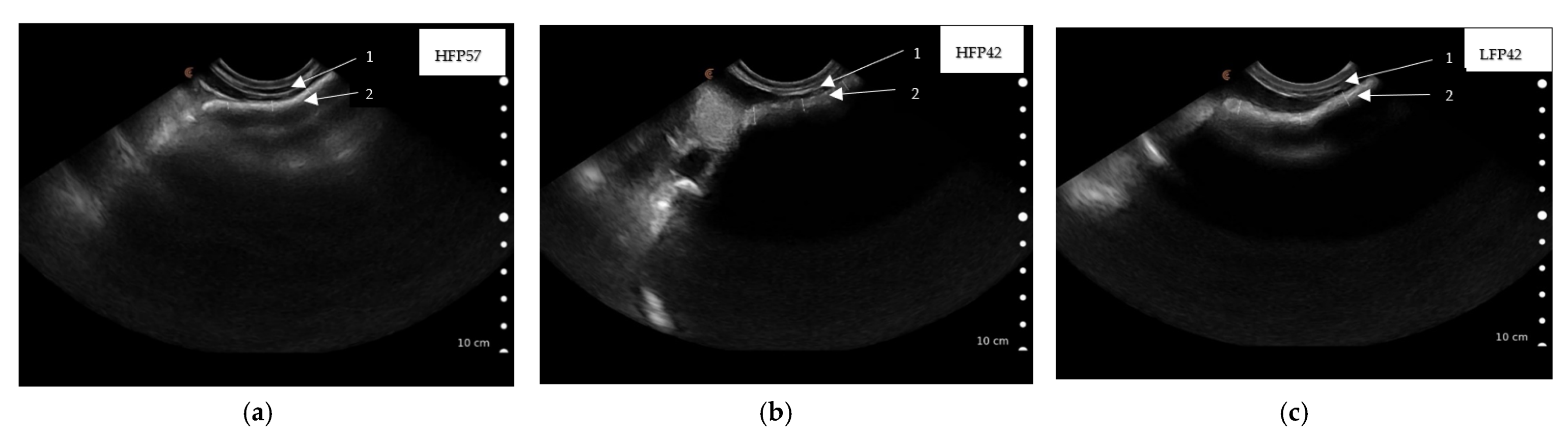
Figure 3.
Ultrasound scanned images showing rumen wall thickness captured caudal to the last rib by placing the Clarius ultrasound scanner (3.0–7.0 MHz) parallel to the last ribs of seven-week (49 ± 1.2 d)-old lambs ((a) HFP57, high fibre concentrate pellet and milk feeding to 57 d of age; (b) HFP42, high fibre concentrate pellet and weaning at 42 d of age; (c) LFP42, low fibre concentrate pellet and weaning at 42 d of age) lying on its right side, 1. Abdominal wall, 2. Rumen wall.
Figure 3.
Ultrasound scanned images showing rumen wall thickness captured caudal to the last rib by placing the Clarius ultrasound scanner (3.0–7.0 MHz) parallel to the last ribs of seven-week (49 ± 1.2 d)-old lambs ((a) HFP57, high fibre concentrate pellet and milk feeding to 57 d of age; (b) HFP42, high fibre concentrate pellet and weaning at 42 d of age; (c) LFP42, low fibre concentrate pellet and weaning at 42 d of age) lying on its right side, 1. Abdominal wall, 2. Rumen wall.
Figure 4.
Schematic diagram of correlations between dietary factors and rumen development parameters of artificially reared lambs given three rearing treatments (significant (p < 0.05) positive influence and tended positive influence). The parameters combined into a same-colour line influence the respective rumen development parameter (e.g., all parameters joined to the black-coloured solid line positively influenced the empty rumen weight).
Figure 4.
Schematic diagram of correlations between dietary factors and rumen development parameters of artificially reared lambs given three rearing treatments (significant (p < 0.05) positive influence and tended positive influence). The parameters combined into a same-colour line influence the respective rumen development parameter (e.g., all parameters joined to the black-coloured solid line positively influenced the empty rumen weight).
Table 1.
Pellet composition in fresh matter basis (Adapted from Herath et al. [
15]).
Table 1.
Pellet composition in fresh matter basis (Adapted from Herath et al. [
15]).
| Ingredient | Low Fibre Concentrate Pellet (LFP) | High Fibre Concentrate Pellet (HFP) |
|---|
| Barley, g/kg | 0.270 | 0.390 |
| Broll, g/kg 1 | 0.000 | 0.351 |
| Soya bean meal, g/kg | 0.225 | 0.218 |
| Wheat, g/kg | 0.389 | 0.000 |
| Molasses, g/kg 2 | 0.030 | 0.030 |
| Skim milk powder, g/kg | 0.075 | 0.000 |
| Limestones, g/kg | 0.010 | 0.010 |
| Sheep premix, g/kg 3 | 0.001 | 0.001 |
Table 2.
Overall means, standard deviation (SD), and range for pellet, lucerne chaff and nutrients intakes, empty rumen weight, and rumen volume of the lambs included in the study (DMI, dry matter intake; ME, metabolisable energy; CP, crude protein; ADF, acid detergent fibre; NDF, neutral detergent fibre).
Table 2.
Overall means, standard deviation (SD), and range for pellet, lucerne chaff and nutrients intakes, empty rumen weight, and rumen volume of the lambs included in the study (DMI, dry matter intake; ME, metabolisable energy; CP, crude protein; ADF, acid detergent fibre; NDF, neutral detergent fibre).
| Parameter | Mean (n = 22) | SD | Range |
|---|
| Total DMI from pellets, kg | 7.60 | 3.31 | 1.76–12.98 |
| Total DMI from lucerne chaff, kg | 0.29 | 0.16 | 0.02–0.58 |
| Total DMI, kg | 7.89 | 3.37 | 2.01–13.47 |
| Total ME intake, MJ | 99.17 | 42.72 | 24.74–171.10 |
| Total CP intake, g | 1616.97 | 697.34 | 404.56–2800.56 |
| Combined CP:ME intake, g/MJ | 16.31 | 0.08 | 16.19–16.38 |
| Organic matter intake from pellets and lucerne chaff 1, kg | 7.45 | 3.19 | 1.90–12.74 |
| Total ADF intake, g | 661.24 | 287.05 | 197.08–1254.23 |
| Total NDF intake, g | 1698.45 | 825.25 | 525.26–3380.44 |
| Total hemicellulose intake 2, g | 1037.21 | 547.57 | 312.37–2126.21 |
| Empty rumen weight, g | 238.46 | 68.10 | 125.30–404.60 |
| Rumen volume, mL | 1099.00 | 327.88 | 552.59–1917.81 |
Table 3.
Effect of three lamb-rearing treatments (HFP57, high fibre concentrate pellets and milk feeding to 57 d of age; HFP42, high fibre concentrate pellets and early weaning at 42 d of age; LFP42, low fibre concentrate pellets and early weaning at 42 d of age) on volatile fatty acid (VFA) content of rumen fluid from artificially reared lambs at slaughter (SE, standard error).
Table 3.
Effect of three lamb-rearing treatments (HFP57, high fibre concentrate pellets and milk feeding to 57 d of age; HFP42, high fibre concentrate pellets and early weaning at 42 d of age; LFP42, low fibre concentrate pellets and early weaning at 42 d of age) on volatile fatty acid (VFA) content of rumen fluid from artificially reared lambs at slaughter (SE, standard error).
| Parameter | Treatment LS Means | Pooled SE | p-Value |
|---|
| HFP57 * | HFP42 * | LFP42 * |
|---|
| Acetic, mg/g | 0.85 (1.41) | 0.78 (1.29) | 0.93 (1.56) | 0.100 | 0.33 |
| Propionic, mg/g | 0.38 (0.51) | 0.38 (0.52) | 0.42 (0.57) | 0.041 | 0.48 |
| n-Butyric, mg/g | 0.13 a (0.15) | 0.17 b (0.20) | 0.19 b (0.22) | 0.021 | 0.019 |
| iso-Butyric, mg/g | 0.06 (0.073) | 0.07 (0.080) | 0.08 (0.090) | 0.008 | 0.18 |
| n-Valeric, mg/g | 0.057 a (0.056) | 0.063 a (0.063) | 0.087 b (0.085) | 0.0106 | 0.018 |
| iso-Valeric, mg/g | 0.08 (0.075) | 0.08 (0.081) | 0.10 (0.097) | 0.021 | 0.29 |
| n-Caproic, mg/g | 0.03 (0.026) | 0.03 (0.031) | 0.05 (0.046) | 0.011 | 0.06 |
| Acetic to propionic ratio 1 | 2.22 (2.75) | 1.99 (2.46) | 2.19 (2.74) | 0.116 | 0.11 |
| Total VFA, mg/g 2 | 1.58 (22.9) | 1.58 (22.6) | 1.86 (26.7) | 0.172 | 0.19 |
Table 4.
Effect of three lamb-rearing treatments (HFP57, high fibre concentrate pellets and milk feeding to 57 d of age; HFP42, high fibre concentrate pellets and early weaning; LFP42, low fibre concentrate pellets and early weaning) on papillae height (µm), width (µm), density (papillae number/cm), muscle layer thickness (µm, MLT), longer (>500 µm) papillae percentage and papillae surface length to straight tissue length (PSL/STL) ratio at the rumen dorsal and ventral sites (D, dorsal rumen site; V, ventral rumen site; SE, standard error).
Table 4.
Effect of three lamb-rearing treatments (HFP57, high fibre concentrate pellets and milk feeding to 57 d of age; HFP42, high fibre concentrate pellets and early weaning; LFP42, low fibre concentrate pellets and early weaning) on papillae height (µm), width (µm), density (papillae number/cm), muscle layer thickness (µm, MLT), longer (>500 µm) papillae percentage and papillae surface length to straight tissue length (PSL/STL) ratio at the rumen dorsal and ventral sites (D, dorsal rumen site; V, ventral rumen site; SE, standard error).
| Variable | Back Transformed Log LSmeans (logLSmeans) 1 | Pooled Log SE | Back Transformed Log LSmeans (logLSmeans) 1 | Pooled Log SE | Back Transformed Log LSmeans (logLSmeans) 1 | Pooled Log SE | p-Value |
|---|
| Treatment * Rumen Site |
|---|
| Treatment | Rumen site | HFP57 | HFP42 | LFP42 |
|---|
| HFP57 | HFP42 | LFP42 | D | V | D | V | D | V | D | V | Treat-ment | Rumen Site | Treatment * Rumen site |
|---|
| Papillae height | 648.3 | 748.7 | 796.5 | 0.04 | 649.2 a | 817.3 b | 0.02 | 569.4 | 738.2 | 675.8 | 829.5 | 711.4 | 891.9 | 0.04 | 0.23 | <0.0001 | 0.8651 |
| (2.8) | (2.9) | (2.9) | (2.8) | (2.9) | (2.8) | (2.9) | (2.8) | (2.9) | (2.9) | (3.0) |
| Papillae width | 419.6 | 421.4 | 437.7 | 0.02 | 442.7 b | 410.2 a | 0.01 | 478.0 b | 368.3 a | 414.0 ab | 428.7 b | 438.3 b | 437.1 b | 0.02 | 0.81 | 0.0006 | <0.0001 |
| (2.6) | (2.6) | (2.6) | (2.7) | (2.6) | (2.68) | (2.57) | (2.62) | (2.57) | (2.64) | (2.64) |
| Papillae density | 16.2 | 13.7 | 14.2 | 0.04 | 13.4 a | 16.1 b | 0.02 | 13.5 a | 19.5 b | 13.3 a | 14.1 a | 13.3 a | 15.1 a | 0.04 | 0.33 | <0.0001 | 0.0055 |
| (1.2) | (1.1) | 1.2) | (1.1) | (1.2) | (1.1) | (1.3) | (1.1) | (1.2) | (1.1) | (1.2) |
| MLT | 1112.8 | 834.1 | 874.0 | 0.04 | 1006.0 b | 864.6 a | 0.02 | 1224.1 c | 1011.6 a | 913.3 a | 761.9 b | 910.8 ab | 838.7 ab | 0.04 | 0.08 | <0.0001 | 0.0033 |
| (3.1) | (2.9) | (2.9) | | (3.0) | (2.9) | | (3.09) | (3.00) | (2.96) | (2.88) | (2.96) | (2.92) | | | | |
| Papillae > 500 µm% 2 | 63.0 | 64.5 | 68.1 | 0.03 | 59.0 b | 71.9 a | 0.02 | 57.0 | 69.6 | 58.8 | 70.7 | 61.3 | 75.7 | 0.03 | 0.67 | <0.0001 | 0.9696 |
| (1.8) | (1.8) | (1.8) | (1.2) | (1.9) | (1.76) | (1.84) | (1.77) | (1.85) | (1.79) | (1.88) |
| PSL/STL 3 | 4.0 | 4.2 | 4.3 | 0.04 | 3.6 a | 4.9 b | 0.03 | 3.2 | 4.9 | 3.8 | 4.7 | 3.7 | 5.0 | 0.04 | 0.82 | <0.0001 | 0.1232 |
| (0.6) | (0.6) | (0.6) | (0.6) | (0.7) | (0.51) | (0.69) | (0.58) | (0.67) | (0.57) | (0.70) |
Table 5.
Effect of three lamb-rearing treatments (HFP57, high fibre concentrate pellets and milk feeding to 57 d of age; HFP42, high fibre concentrate pellets and early weaning at 42 d of age; LFP42, low fibre concentrate pellets and early weaning at 42 d of age) on lamb rumen dorsal wall thickness at five and seven weeks of age as measured from Ultrasound scanned images (SE, standard error).
Table 5.
Effect of three lamb-rearing treatments (HFP57, high fibre concentrate pellets and milk feeding to 57 d of age; HFP42, high fibre concentrate pellets and early weaning at 42 d of age; LFP42, low fibre concentrate pellets and early weaning at 42 d of age) on lamb rumen dorsal wall thickness at five and seven weeks of age as measured from Ultrasound scanned images (SE, standard error).
| Lambs Age | Treatment | Pooled SE | p-Value |
|---|
| HFP57 | HFP42 | LFP42 | Treatment | Lamb 1 | Image 2 |
|---|
| 5 weeks (36 ± 1.2 d of age), cm | 0.25 | 0.20 | 0.22 | 0.04 | 0.7299 | <0.0001 | 0.0014 |
| 7 weeks (49 ± 1.2 d of age), cm | 0.33 a | 0.48 b | 0.56 b | 0.05 | 0.0068 | <0.0001 | 0.0204 |
Table 6.
Pearson correlation coefficients of dietary factors and rumen physical development parameters of artificially reared lambs exposed to three rearing treatments (DMI, dry matter intake; ME, metabolisable energy; CP, crude protein; ADF, acid detergent fibre; NDF, neutral detergent fibre).
Table 6.
Pearson correlation coefficients of dietary factors and rumen physical development parameters of artificially reared lambs exposed to three rearing treatments (DMI, dry matter intake; ME, metabolisable energy; CP, crude protein; ADF, acid detergent fibre; NDF, neutral detergent fibre).
| Parameter | Empty Rumen Weight, g | Rumen Volume, mL | Papillae Height-Dorsal, μm | Papillae Height-Ventral, μm | Papillae Width-Dorsal, μm | Papillae Width-Ventral, μm | Muscle Layer Thickness-Dorsal, μm | Muscle Layer Thickness-Ventral, μm |
|---|
| Total DMI from pellets, kg | 0.85 ** | 0.51 * | 0.58 * | 0.53 * | −0.05 | 0.30 | −0.19 | −0.16 |
| Total DMI from lucerne chaff, kg | 0.35 | 0.55 * | −0.03 | −0.05 | 0.31 | 0.24 | 0.21 | −0.14 |
| Total DMI, kg | 0.86 ** | 0.53 * | 0.54 * | 0.51 * | −0.03 | 0.30 | −0.18 | −0.19 |
| Total ME intake, MJ | 0.86 ** | 0.51 * | 0.53 * | 0.52 * | −0.03 | 0.29 | −0.18 | −0.16 |
| Total CP intake, g | 0.86 ** | 0.51 * | 0.53 * | 0.52 * | −0.03 | 0.29 | −0.18 | −0.15 |
| Combined CP:ME intake, g/MJ | −0.07 | −0.36 | −0.25 | −0.09 | 0.08 | −0.26 | 0.20 | 0.23 |
| Organic matter intake from pellets and lucerne chaff 1, kg | 0.86 ** | 0.53 * | 0.54 * | 0.51 * | −0.03 | 0.30 | −0.18 | −0.16 |
| Total ADF intake, g | 0.81 ** | 0.46 * | 0.39 | 0.39 | 0.10 | 0.20 | −0.05 | −0.07 |
| Total NDF intake, g | 0.77 ** | 0.34 | 0.38 | 0.41 * | 0.05 | 0.13 | −0.07 | −0.03 |
| Total hemicellulose intake 2, g | 0.74 ** | 0.28 | 0.36 | 0.42 | 0.03 | 0.09 | −0.08 | −0.004 |
Table 7.
Pearson correlation coefficients of dietary factors and iso-butyric, and iso-valeric content of rumen fluid of artificially reared lambs exposed to three rearing treatments (DMI, dry matter intake; ME, metabolisable energy; CP, crude protein; ADF, acid detergent fibre; NDF, neutral detergent fibre).
Table 7.
Pearson correlation coefficients of dietary factors and iso-butyric, and iso-valeric content of rumen fluid of artificially reared lambs exposed to three rearing treatments (DMI, dry matter intake; ME, metabolisable energy; CP, crude protein; ADF, acid detergent fibre; NDF, neutral detergent fibre).
| Variable | Iso-Butyric, mg/g | Iso-Valeric, mg/g |
|---|
| Total DMI from pellets, kg | 0.54 * | 0.49 * |
| Total DMI from lucerne chaff, kg | 0.45 * | 0.43 * |
| Total DMI, kg | 0.55 * | 0.50 * |
| Total ME intake, MJ | 0.54 * | 0.49 * |
| Total CP intake, g | 0.53 | 0.49 * |
| Organic matter intake from pellets and lucerne chaff 1, kg | 0.55 * | 0.50 * |
| Total ADF intake, g | 0.48 * | 0.45 * |
| Total NDF intake, g | 0.39 | 0.37 |
| Total hemicellulose intake 2, g | 0.34 | 0.32 |


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}