
Alpha-Lipoic Acid Is an Effective Nutritive Antioxidant for Healthy Adult Dogs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Experimental Section
2.1. Study Design
2.2. Test Animals and Feeding
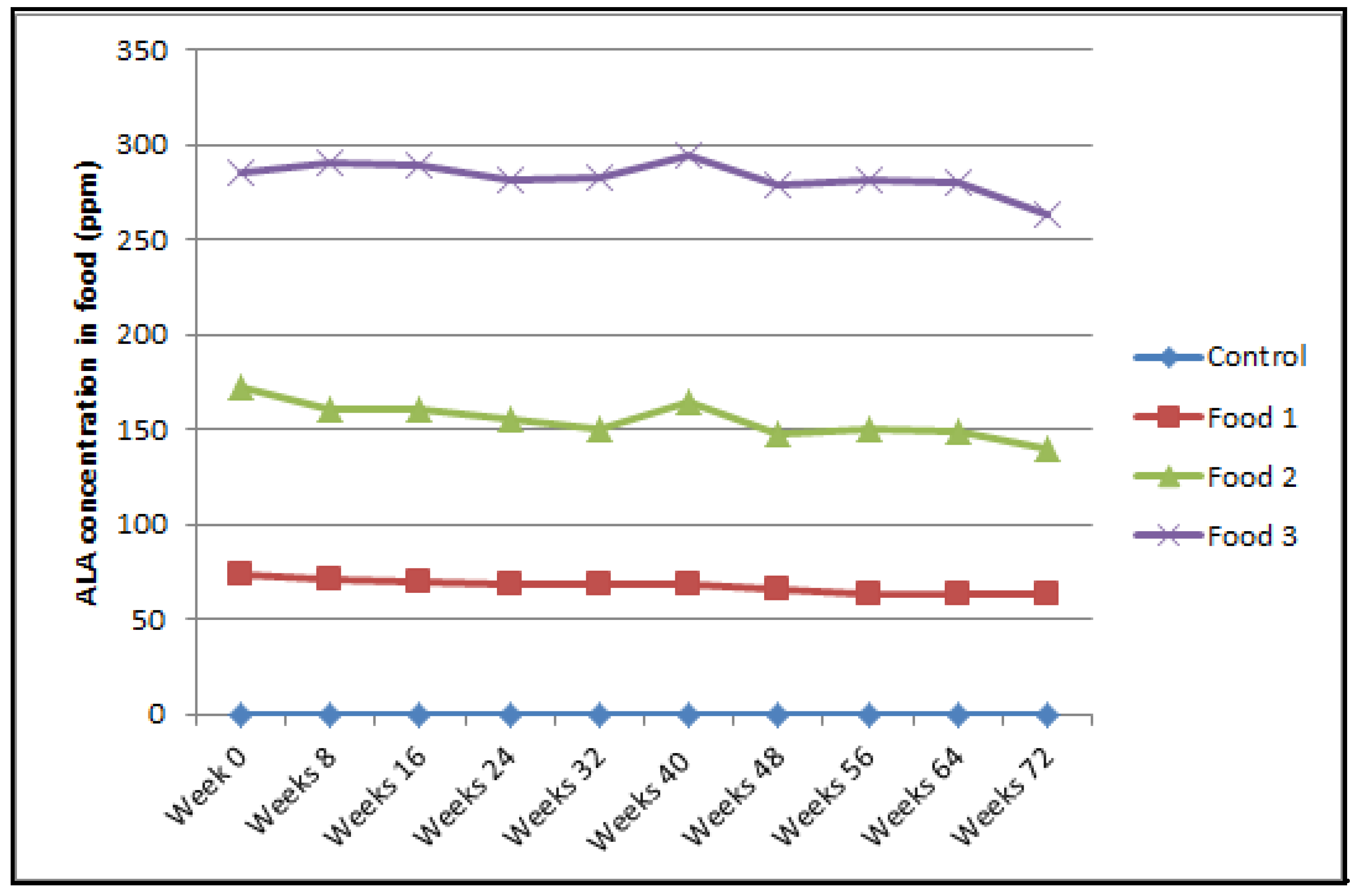
2.3. Concentrations of Alpha-Lipoic Acid in Food
2.4. Glutathione Analytical Methods
2.5. Statistical Analysis and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Ingredient | Food | |||
|---|---|---|---|---|
| Control | Food 1 | Food 2 | Food 3 | |
| Barley | 11.9 | 11.9 | 11.9 | 11.9 |
| Calcium carbonate | 1.0 | 1.0 | 1.0 | 1.0 |
| Chicken Meal | 11.9 | 11.9 | 11.9 | 11.9 |
| Choline chloride | 0.20 | 0.20 | 0.20 | |
| Corn gluten meal | 6.1 | 6.0 | 5.7 | 5.4 |
| Corn | 11.9 | 11.9 | 11.9 | 11.9 |
| Fish oil | 1.8 | 1.8 | 1.8 | 1.8 |
| Lactic acid | 1.2 | 1.2 | 1.2 | |
| Alpha Lipoic acid, 5% | 0.20 | 0.40 | 0.78 | |
| Mineral Premix, | 0.03 | 0.04 | 0.04 | 0.04 |
| Flavor, | 6.00 | 6.00 | 6.00 | 6.00 |
| Pork Fat | 5.0 | 5.0 | 5.0 | 5.0 |
| Potassium chloride | 0.41 | 0.41 | 0.41 | 0.41 |
| Brewers Rice | 11.9 | 11.9 | 11.9 | 11.9 |
| Brown Rice | 5.2 | 5.2 | 5.2 | 5.2 |
| Iodized Sodium chloride | 0.36 | 0.36 | 0.36 | 0.36 |
| Sorghum | 5.0 | 5.0 | 5.0 | 5.0 |
| Soybean Oil | 3.3 | 3.3 | 3.3 | 3.3 |
| Soybean | 5.0 | 5.0 | 5.0 | 5.0 |
| Vitamin Premix | 0.06 | 0.06 | 0.06 | 0.06 |
| Wheat | 11.9 | 11.9 | 11.9 | 11.9 |
| Nutrient | Food | |||
|---|---|---|---|---|
| Control | Food 1 | Food 2 | Food 3 | |
| alpha-Lipoic acid, ppm | 0.0 | 69.1 | 158.2 | 286.1 |
| Ash, % | 5.0 | 4.8 | 5.1 | 5.2 |
| Diet Metabolic Energy, kcal/kg | 3881 | 3814 | 3908 | 3836 |
| Chloride, % | 0.58 | 0.60 | 0.61 | 0.63 |
| Fat Crude, % | 15 | 15 | 15 | 15 |
| Polyunsaturated fatty acids, % | 8.7% | 8.6% | 8.5% | 9.1% |
| Fiber Crude, % | 2.7 | 2.7 | 2.6 | 2.7 |
| Fiber Total Dietary, % | 8.3 | 8.8 | 9.4 | 9.3 |
| Magnesium, % | 0.10 | 0.09 | 0.10 | 0.10 |
| Moisture, % | 9.4 | 9.3 | 9.4 | 9.0 |
| Phosphorus, % | 0.66 | 0.64 | 0.66 | 0.67 |
| Potassium, % | 0.59 | 0.55 | 0.58 | 0.60 |
| Protein Crude, % | 20 | 21 | 20 | 20 |
| Sodium, % | 0.3 | 0.28 | 0.31 | 0.3 |
| Selenium, ppm | 0.51 | 0.45 | 0.45 | 0.56 |
| Vitamin C as AscorbicAcid, ppm | 18 | <17.2 | <17.2 | <17.2 |
| Vitamin E—Total Tocopherols, IU/kg | 108 | 111 | 98 | 110 |
| Analyte | Time Point | Control | Food 1 | Food 2 | Food 3 |
|---|---|---|---|---|---|
| Alb/Glob Normal range 1.1–2.0 | P1 | 1.6 ± 0.24 | 1.5 ± 0.22 | 1.5 ± 0.24 | 1.5 ± 0.24 |
| Month 0 | 1.7 ± 0.27 | 1.6 ± 0.23 | 1.6 ± 0.26 | 1.6 ± 0.48 | |
| Month 6 | 1.6 ± 0.43 | 1.6 ± 0.28 | 1.6 ± 0.27 | 1.6 ± 0.25 | |
| Albumin (g/dL) Normal range 2.9–4.0 | P1 | 3.6 ± 0.33 | 3.7 ± 0.29 | 3.6 ± 0.27 | 3.6 ± 0.28 |
| Month 0 | 3.8 ± 0.29 | 3.9 ± 0.23 | 3.8 ± 0.29 | 3.9 ± 0.25 | |
| Month 6 | 3.7 ± 0.9 | 3.9 ± 0.28 | 3.8 ± 0.35 | 3.9 ± 0.23 | |
| ALP (U/L) Normal range 30–439 | P1 | 102 ± 102 | 78 ± 102 | 72 ± 117 | 72 ± 68 |
| Month 0 | 149 ± 206 | 116 ± 167 | 106 ± 200 | 96 ± 130 | |
| Month 6 | 194 ± 285 | 150 ± 238 | 124 ± 233 | 138 ± 210 | |
| ALT (U/L) Normal range 22–268 | P1 | 52 ± 54 | 33 ± 11.1 | 38 ± 11.8 | 43 ± 32 |
| Month 0 | 37 ± 10.4 | 36 ± 21 | 45 ± 39 | 36 ± 23 | |
| Month 6 | 55 ± 68 | 37 ± 13.1 | 40 ± 23 | 38 ± 22 | |
| Total Bilirubin (mg/dL) Normal range 0.0–35.1 | P1 | 0.06 ± 0.05 | 0.08 ± 0.06 | 0.06 ± 0.05 | 0.07 ± 0.05 |
| Month 0 | 0.09 ± 0.04 | 0.09 ± 0.04 | 0.09 ± 0.05 | 0.07 ± 0.05 | |
| Month 6 | 0.09 ± 0.03 | 0.1 ± 0.04 | 0.09 ± 0.05 | 0.09 ± 0.05 | |
| BUN (mg/dL) Normal range 6.40–20.50 | P1 | 13.7 ± 2.9 | 13.8 ± 2.3 | 13.4 ± 3.6 | 14 ± 2.9 |
| Month 0 | 12.2 ± 3.2 | 12.1 ± 2.3 | 11.6 ± 2.8 | 12.2 ± 2.3 | |
| Month 6 | 13.2 ± 3.5 | 13.1 ± 2.4 | 11.9 ± 2.1 | 13.8 ± 2.6 | |
| Bun/Creat. Normal range 10.30–33.60 | P1 | 20 ± 5.9 | 21 ± 4.5 | 21 ± 5.6 | 22 ± 4.5 |
| Month 0 | 19.2 ± 5.4 | 19 ± 4 | 18.8 ± 3.9 | 20 ± 4.6 | |
| Month 6 | 20 ± 5.2 | 20 ± 3.9 | 19.9 ± 5.1 | 22 ± 5.3 | |
| Calcium (mg/dL) Normal range 8.7–12.0 | P1 | 10.3 ± 0.4 | 10.2 ± 0.49 | 10.2 ± 0.37 | 10.2 ± 0.55 |
| Month 0 | 10.3 ± 0.44 | 10.6 ± 0.49 | 10.3 ± 0.4 | 10.4 ± 0.61 | |
| Month 6 | 10.4 ± 0.52 | 10.4 ± 0.49 | 10.3 ± 0.4 | 10.2 ± 0.61 | |
| Chloride (mmol/L) Normal range 105–115 | P1 | 110 ± 2.5 | 110 ± 2.1 | 110 ± 1.9 | 109 ± 2 |
| Month 0 | 112 ± 2.5 | 113 ± 2.4 | 113 ± 2.2 | 112 ± 2 | |
| Month 6 | 108 ± 1.9 | 109 ± 1.7 | 106 ± 10.6 | 108 ± 3.3 | |
| Cholesterol (mg/dL) Normal range 133–401 | P1 | 203 ± 44 | 202 ± 38 | 202 ± 32 | 186 ± 36 |
| Month 0 | 215 ± 64 | 212 ± 50 | 215 ± 46 | 195 ± 37 | |
| Month 6 | 213 ± 61 | 208 ± 48 | 219 ± 52 | 191 ± 37 | |
| Creatinine (mg/dL) Normal range 0.41–0.82 | P1 | 0.71 ± 0.16 | 0.69 ± 0.1 | 0.68 ± 0.19 | 0.65 ± 0.11 |
| Month 0 | 0.66 ± 0.14 | 0.66 ± 0.1 | 0.64 ± 0.21 | 0.62 ± 0.12 | |
| Month 6 | 0.66 ± 0.13 | 0.66 ± 0.18 | 0.63 ± 0.19 | 0.64 ± 0.13 | |
| Globulin (g/dL) Normal range 1.9–2.9 | P1 | 2.4 ± 0.27 | 2.5 ± 0.28 | 2.4 ± 0.29 | 2.5 ± 0.44 |
| Month 0 | 2.3 ± 0.25 | 2.5 ± 0.29 | 2.4 ± 0.29 | 2.6 ± 0.8 | |
| Month 6 | 2.6 ± 0.94 | 2.4 ± 0.29 | 2.4 ± 0.32 | 2.5 ± 0.37 | |
| Glucose (mg/dL Normal range) 58–96 | P1 | 84 ± 11.3 | 83 ± 8.1 | 85 ± 9 | 82 ± 9.5 |
| Month 0 | 84 ± 8.1 | 76 ± 10 | 79 ± 9.9 | 80 ± 7.5 | |
| Month 6 | 87 ± 10 | 85 ± 10 | 85 ± 11 | 86 ± 10.1 | |
| Magnesium (mg/dL) Normal range 1.7–2.3 | P1 | 2 ± 0.13 | 2 ± 0.13 | 2.1 ± 0.19 | 2.1 ± 0.13 |
| Month 0 | 2.1 ± 0.14 | 2 ± 0.1 | 2.1 ± 0.15 | 2.1 ± 0.17 | |
| Month 6 | 1.9 ± 0.13 | 1.9 ± 0.2 | 2 ± 0.13 | 2 ± 0.15 | |
| NA:K Normal range 28–37 | P1 | 30 ± 1.6 | 31 ± 1 | 31 ± 2.6 | 31 ± 1.8 |
| Month 0 | 31 ± 2.4 | 31 ± 1.6 | 31 ± 1.7 | 31 ± 1.4 | |
| Month 6 | 31 ± 1.7 | 32 ± 2.6 | 31 ± 2 | 32 ± 3 | |
| Phosphorus (mg/dL) Normal range 2.9–6.1 | P1 | 4.4 ± 0.73 | 4.2 ± 0.68 | 4.5 ± 0.57 | 4.2 ± 0.72 |
| Month 0 | 4.1 ± 0.55 | 4.1 ± 0.65 | 4.3 ± 0.55 | 4 ± 0.39 | |
| Month 6 | 4.3 ± 0.47 | 4 ± 0.57 | 4.3 ± 0.71 | 4.1 ± 0.56 | |
| Potassium (mmol/L) Normal range 4–5.3 | P1 | 4.9 ± 0.25 | 4.8 ± 0.19 | 4.9 ± 0.42 | 4.8 ± 0.24 |
| Month 0 | 4.8 ± 0.38 | 4.8 ± 0.23 | 4.8 ± 0.26 | 4.7 ± 0.23 | |
| Month 6 | 4.9 ± 0.24 | 4.7 ± 0.35 | 4.8 ± 0.53 | 4.7 ± 0.39 | |
| Total Protein (g/dL) Normal range 5.1–6.5 | P1 | 6 ± 0.41 | 6.1 ± 0.38 | 6.1 ± 0.36 | 6.2 ± 0.49 |
| Month 0 | 6.1 ± 0.32 | 6.3 ± 0.29 | 6.1 ± 0.41 | 6.3 ± 0.33 | |
| Month 6 | 6.3 ± 0.44 | 6.3 ± 0.3 | 6.2 ± 0.43 | 6.4 ± 0.41 | |
| Sodium (mmol/L) Normal range 144–150 | P1 | 149 ± 1.3 | 149 ± 1.6 | 150 ± 1.8 | 148 ± 1.7 |
| Month 0 | 149 ± 2.1 | 150 ± 2 | 149 ± 1.4 | 148 ± 1.8 | |
| Month 6 | 149 ± 1.8 | 150 ± 15 | 147 ± 13.6 | 149 ± 1.9 | |
| Triglycerides (mg/dL) Normal range 34–429 | P1 | 107 ± 87 | 124 ± 107 | 78 ± 41 | 80 ± 59 |
| Month 0 | 94 ± 78 | 89 ± 53 | 88 ± 50 | 64 ± 17.1 | |
| Month 6 | 98 ± 75 | 91 ± 61 | 85 ± 63 | 67 ± 24 |
| Analyte | Phase | Control | Food 1 | Food 2 | Food 3 |
|---|---|---|---|---|---|
| Basophils (%) | P1 | 0.19 ± 0.12 | 0.09 ± 0.1 | 0.15 ± 0.09 | 0.14 ± 0.10 |
| Month 0 | 0.12 ± 0.06 | 0.09 ± 0.08 | 0.11 ± 0.10 | 0.11 ± 0.07 | |
| Month 6 | 0.09 ± 0.09 | 0.1 ± 0.08 | 0.11 ± 0.15 | 0.09 ± 0.07 | |
| Absolute Basophils (k/µL) Normal range 0.00–0.10 | P1 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 |
| Month 0 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | |
| Month 6 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | |
| Eosinophils (%) | P1 | 5.3 ± 2.5 | 6.2 ± 4.3 | 5.5 ± 2.4 | 6.7 ± 5.1 |
| Month 0 | 4.7 ± 2.4 | 5.3 ± 3 | 4.1 ± 1.6 | 7.2 ± 5.9 | |
| Month 6 | 4.4 ± 2.2 | 4.6 ± 2.6 | 3.8 ± 1.5 | 6.7 ± 4.8 | |
| Absolute Eosinophils (k/µL) Normal range 0.00–1.40 | P1 | 0.44 ± 0.31 | 0.5 ± 0.38 | 0.44 ± 0.24 | 0.63 ± 0.66 |
| Month 0 | 0.38 ± 0.19 | 0.43 ± 0.28 | 0.34 ± 0.18 | 0.68 ± 0.78 | |
| Month 6 | 0.33 ± 0.16 | 0.36 ± 0.21 | 0.31 ± 0.18 | 0.59 ± 0.55 | |
| HCT (%) Normal range 33.0–58.7 | P1 | 52 ± 3.7 | 50 ± 3.7 | 51 ± 4.3 | 51 ± 3.7 |
| Month 0 | 49 ± 4.5 | 50 ± 5.2 | 49 ± 3.3 | 50 ± 4.1 | |
| Month 6 | 49 ± 3.3 | 49 ± 4.6 | 49 ± 5.4 | 48 ± 4.7 | |
| HGB (g/dL) Normal range 10.5–20.1 | P1 | 17.2 ± 1.3 | 16.6 ± 1.2 | 17 ± 1.5 | 17 ± 1.3 |
| Month 0 | 16.8 ± 1.6 | 16.9 ± 1.7 | 16.7 ± 1.3 | 16.9 ± 1.5 | |
| Month 6 | 16.3 ± 1.2 | 16.2 ± 1.6 | 16.4 ± 1.8 | 16 ± 1.7 | |
| Lymphocytes (%) | P1 | 27 ± 6 | 26 ± 7 | 26 ± 6 | 26 ± 6 |
| Month 0 | 24 ± 6 | 25 ± 7 | 24 ± 6 | 24 ± 6 | |
| Month 6 | 23 ± 6 | 22 ± 7 | 23 ± 7 | 22 ± 5 | |
| Absolute Lymphocytes (k/µL) Normal range 0.30–3.90 | P1 | 2.1 ± 0.4 | 2.1 ± 0.9 | 1.9 ± 0.5 | 2.2 ± 0.7 |
| Month 0 | 2 ± 0.6 | 2.1 ± 0.9 | 1.9 ± 0.5 | 2.1 ± 0.8 | |
| Month 6 | 1.8 ± 0.7 | 1.8 ± 0.9 | 1.7 ± 0.5 | 1.9 ± 0.7 | |
| MCH (pg) Normal range 21.0–27.0 | P1 | 23 ± 0.87 | 23 ± 0.71 | 23 ± 0.84 | 23 ± 0.87 |
| Month 0 | 23 ± 1.00 | 23 ± 1.00 | 23 ± 0.92 | 23 ± 0.77 | |
| Month 6 | 23 ± 0.95 | 23 ± 0.83 | 23 ± 0.95 | 23 ± 0.79 | |
| MCHC (g/dL)Normal range 30.1–41.9 | P1 | 33 ± 0.54 | 33 ± 0.86 | 33 ± 0.65 | 33 ± 0.56 |
| Month 0 | 34 ± 0.67 | 34 ± 1.20 | 34 ± 0.70 | 34 ± 0.65 | |
| Month 6 | 33 ± 0.64 | 33 ± 0.69 | 33 ± 0.7 | 33 ± 0.68 | |
| MCV (fL) Normal range 63.0–78.3 | P1 | 69 ± 2.9 | 70 ± 3.1 | 69 ± 3.1 | 69 ± 3.2 |
| Month 0 | 68 ± 3.0 | 68 ± 2.9 | 69 ± 3.1 | 69 ± 2.8 | |
| Month 6 | 68 ± 2.8 | 69 ± 2.7 | 69 ± 3.4 | 69 ± 2.9 | |
| Monocytes (%) | P1 | 6.1 ± 1.3 | 5.8 ± 1.8 | 6.7 ± 2.7 | 5.6 ± 1.6 |
| Month 0 | 6 ± 1.1 | 6.4 ± 1.6 | 6.8 ± 2.2 | 6.3 ± 1.6 | |
| Month 6 | 5.4 ± 1.4 | 5.5 ± 1.8 | 6 ± 2.3 | 5.8 ± 1.5 | |
| Absolute Monocytes (k/µL) Normal range 0.00–1.40 | P1 | 0.47 ± 0.12 | 0.48 ± 0.23 | 0.49 ± 0.18 | 0.5 ± 0.23 |
| Month 0 | 0.5 ± 0.12 | 0.53 ± 0.22 | 0.53 ± 0.23 | 0.54 ± 0.16 | |
| Month 6 | 0.42 ± 0.15 | 0.45 ± 0.17 | 0.46 ± 0.23 | 0.49 ± 0.16 | |
| Neutrophils (%) | P1 | 61 ± 5 | 62 ± 9 | 61 ± 7 | 62 ± 8 |
| Month 0 | 65 ± 7 | 63 ± 8 | 65 ± 7 | 63 ± 9 | |
| Month 6 | 67 ± 7 | 68± 8 | 68 ± 9 | 66 ± 8 | |
| Absolute Neutrophils (k/µL) Normal range 2.5–15.7 | P1 | 4.8 ± 1.4 | 5.1 ± 1.5 | 4.7 ± 1.6 | 5.4 ± 2.0 |
| Month 0 | 5.4 ± 1.1 | 5.3 ± 1.9 | 5.3 ± 1.7 | 5.4 ± 1.4 | |
| Month 6 | 5.2 ± 1.4 | 5.7 ± 2.1 | 5.4 ± 2.0 | 5.6 ± 1.7 | |
| Platelets (k/µL) Normal range 140–540 | P1 | 274 ± 73 | 270 ± 83 | 305 ± 95 | 315 ± 78 |
| Month 0 | 300 ± 86 | 265 ± 103 | 319 ± 104 | 312 ± 83 | |
| Month 6 | 294 ± 103 | 277 ± 98 | 343 ± 113 | 313 ± 110 | |
| RBC (M/µL) Normal range 4.48–8.53 | P1 | 7.6 ± 0.67 | 7.2 ± 0.51 | 7.5 ± 0.66 | 7.4 ± 0.63 |
| Month 0 | 7.3 ± 0.68 | 7.3 ± 0.81 | 7.2 ± 0.63 | 7.2 ± 0.75 | |
| Month 6 | 7.2 ± 0.58 | 7.1 ± 0.76 | 7.2 ± 0.81 | 7 ± 0.85 | |
| RDW (fL) Normal range 12.9–21.0 | P1 | 36 ± 1.2 | 37 ± 7.4 | 36 ± 1.4 | 36 ± 1.5 |
| Month 0 | 36 ± 1.3 | 37 ± 6.6 | 35 ± 1.4 | 35 ± 1.5 | |
| Month 6 | 36 ± 1.4 | 36 ± 1.4 | 36 ± 1.7 | 35 ± 1.3 | |
| Reticulocytes (%) Normal range 0.31–1.58 | P1 | 0.69 ± 0.41 | 0.64 ± 0.27 | 0.58 ± 0.23 | 0.98 ± 0.48 |
| Month 0 | 0.70 ± 0.21 | 0.83 ± 0.42 | 0.58 ± 0.2 | 0.80 ± 0.36 | |
| Month 6 | 0.79 ± 0.31 | 0.77 ± 0.35 | 0.71 ± 0.34 | 0.77 ± 0.38 | |
| Absolute Reticulocytes (M/ul) | P1 | 0.05 ± 0.03 | 0.05 ± 0.02 | 0.04 ± 0.02 | 0.07 ± 0.04 |
| Month 0 | 0.05 ± 0.02 | 0.06 ± 0.03 | 0.04 ± 0.02 | 0.06 ± 0.03 | |
| Month 6 | 0.06 ± 0.02 | 0.06 ± 0.03 | 0.05 ± 0.03 | 0.05 ± 0.03 | |
| WBC (ku/µL Normal range) 4.0–18.2 | P1 | 7.8 ± 1.8 | 8.2 ± 2.1 | 7.6 ± 2.2 | 8.7 ± 2.8 |
| Month 0 | 8.3 ± 1.4 | 8.3 ± 2.7 | 8.1 ± 2.2 | 8.7 ± 2.3 | |
| Month 6 | 7.8 ± 1.9 | 8.3 ± 2.6 | 7.9 ± 2.6 | 8.5 ± 2.5 |



References
- Reed, L.J. A trail of research from lipoic acid to alpha-keto acid dehydrogenase complexes. J. Biol. Chem. 2001, 276, 38329–38336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreau, J.P. Biosynthesis of lipoic acid via unsaturated fatty acids. Methods Enzymol. 1979, 62, 152–158. [Google Scholar] [PubMed]
- Kramer, K.; Packer LR-a-LAIn: Kramer, K.; Hoppe, P.; Packer, L. (Eds.) Nutraceuticals in Health and Disease Prevention; Marcel Dekker, Inc.: New York, NY, USA, 2001; pp. 129–164. [Google Scholar]
- Reed, L.J. Multienzyme complexes. Acc. Chem. Res. 1974, 7, 40–46. [Google Scholar] [CrossRef]
- Ames, B.N. Micronutrients prevent cancer and delay aging. Toxicol. Lett. 1998, 102–103, 5–18. [Google Scholar] [CrossRef]
- Gorąca, A.; Huk-Kolega, H.; Piechota, A.; Kleniewska, P.; Ciejka, E.; Skibska, B. Lipoic acid-biological activity and therapeutic potential. Pharmacol. Rep. 2011, 63, 849–858. [Google Scholar] [CrossRef]
- El Barky, A.R.; Hussein, S.A.; Mohamed, T.M. The Potent Antioxidant Alpha Lipoic Acid. J. Plant Chem. Ecophysiol. 2017, 2, 1016. [Google Scholar]
- Ross, S.M. Clinical applications of lipoic acid in type II diabetes mellitus. Holist. Nurs. Pract. 2006, 20, 305–306. [Google Scholar] [CrossRef]
- Hendler, S.S.; Rorvik, D.R. (Eds.) PDR for Nutritional Supplements; Medical Economics Company, Inc.: Montvale, NJ, USA, 2001. [Google Scholar]
- Foster, T.S. Efficacy and safety of alpha-lipoic acid supplementation in the treatment of symptomatic diabetic neuropathy. Diabetes Educ. 2007, 33, 111–117. [Google Scholar] [CrossRef]
- Jacob, S.; Henriksen, E.J.; Schiemann, A.L.; Simon, I.; Clancy, D.E.; Tritschler, H.J.; Jung, W.I.; Augustin, H.J.; Dietze, G.J. Enhancement of glucose disposal in patients with type 2 diabetes by alpha-lipoic acid. Arzneimittelforschung 1995, 45, 872–874. [Google Scholar]
- Jacob, S.; Rett, K.; Henriksen, E.J.; Haring, H.U. Thioctic acid—Effects on insulin sensitivity and glucose-metabolism. Biofactors 1999, 10, 169–174. [Google Scholar] [CrossRef]
- Akbari, M.; Ostadmohammadi, V.; Lankarani, K.B.; Tabrizi, R.; Kolahdooz, F.; Khatibi, S.R.; Asemi, Z. The effects of alpha-lipoic acid supplementation on glucose control and lipid profiles among patients with metabolic diseases: A systematic review and meta-analysis of randomized controlled trials. Metabolism 2018, 87, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Nowak, H.; Kempler, P.; Vargha, P.; Low, P.A. Treatment of symptomatic diabetic polyneuropathy with the antioxidant alpha-lipoic acid: A meta-analysis. Diabet. Med. 2004, 21, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Hanefeld, M.; Ruhnau, K.J.; Hasche, H.; Lobisch, M.; Schütte, K.; Kerum, G.; Malessa, R. Treatment of symptomatic diabetic polyneuropathy with the antioxidant alpha-lipoic acid: A 7-month multicenter randomized controlled trial (ALADIN III Study). ALADIN III Study Group. Alpha-lipoic acid in diabetic neuropathy. Diabetes Care 1999, 22, 1296–1301. [Google Scholar]
- Ziegler, D.; Low, P.A.; Litchy, W.J.; Boulton, A.J.M.; Vinik, A.I.; Freeman, R.; Samigullin, R.; Tritschler, H.; Munzel, U.; Maus, J.; et al. Efficacy and safety of antioxidant treatment with alpha-lipoic acid over 4 years in diabetic polyneuropathy: The NATHAN 1 trial. Diabetes Care 2011, 34, 2054–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streeper, R.S.; Henriksen, E.J.; Jacob, S.; Hokama, J.Y.; Fogt, D.L.; Tritschler, H.J. Differential effects of lipoic acid stereoisomers on glucose metabolism in insulin resistant skeletal muscle. Am. J. Physiol. 1997, 273, E185–E191. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Streeper, R.S.; Fogt, D.L.; Hokama, J.Y.; Tritschler, H.J.; Dietze, G.J.; Henriksen, E.J. The antioxidant alpha-lipoic acid enhances insulin-stimulated glucose metabolism in insulin-resistant rat skeletal muscle. Diabetes 1996, 45, 1024–1029. [Google Scholar] [CrossRef]
- Vasdev, S.; Ford, C.A.; Parai, S.; Longerich, L.; Gadag, V. Dietary alpha-lipoic acid supplementation lowers blood pressure in spontaneously hypertensive rats. J. Hypertens. 2000, 18, 567–573. [Google Scholar] [CrossRef]
- Suh, J.H.; Shigeno, E.T.; Morrow, J.D.; Cox, B.; Rocha, A.E.; Frei, B.; Hagen, T.M. Oxidative stress in the aging rat heart is reversed by dietary supplementation with (R)-(alpha)-lipoic acid. FASEB J. 2001, 15, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Zicker, S.C.; Hagen, T.M.; Joisher, N.; Golder, C.; Joshi, D.K.; Miller, E.P. Safety of long-term feeding of dl-α-lipoic acid and its effect of reduced glutathione:oxidized glutathione ratios in Beagles. Vet. Ther. 2002, 3, 167–176. [Google Scholar]
- Chaudhary, P.; Marracci, G.H.; Bourdette, D.N. Lipoic acid inhibits expression of ICAM-1 and VCAM-1 by CNS endothelial cells and T cell migration into the spinal cord in experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2006, 175, 87–96. [Google Scholar] [CrossRef]
- Lee, E.Y.; Lee, C.K.; Lee, K.U.; Park, J.Y.; Cho, K.J.; Cho, Y.S.; Lee, H.R.; Moon, S.H.; Moon, H.B.; Yoo, B. Alpha-lipoic acid suppresses the development of collagen-induced arthritis and protects against bone destruction in mice. Rheumatol. Int. 2007, 27, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Gupta, Y.K. Effect of alpha lipoic acid on intracerebroventricular streptozotocin model of cognitive impairment in rats. Eur. Neuropsychopharmacol. 2003, 13, 241–247. [Google Scholar] [CrossRef]
- Milgram, N.W.; Zicker, S.C.; Head, E.; Muggenburg, B.A.; Murphey, H.; Ikeda-Douglas, C.J.; Cotman, C.W. Dietary enrichment counteracts age-associated cognitive dysfunction in canines. Neurobiol. Aging 2002, 23, 737–745. [Google Scholar] [CrossRef]
- Milgram, N.W.; Head, E.; Zicker, S.C.; Ikeda-Douglas, C.; Murphey, H.; Muggenberg, B.A.; Siwak, C.T.; Tapp, P.D.; Lowry, S.R.; Cotman, C.W. Long-term treatment with antioxidants and a program of behavioral enrichment reduces age-dependent impairment in discrimination and reversal learning in beagle dogs. Exp. Gerontol. 2004, 39, 753–765. [Google Scholar] [CrossRef]
- Sumathi, R.; Baskaran, G.; Varalakshmi, P. Relationship between glutathione and DL a-Lipoic Acid against cadmium-induced hepatotoxicity. Jpn. J. Med. Sci. Biol. 1996, 49, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Kagan, V.E.; Shvedova, A.; Serbinova, E.; Khan, S.; Swanson, C.; Powell, R.; Packer, L. Dihydrolipoic Acid--A Universal Antioxidant Both in the Membrane and in the Aqueous Phase. Reduction of Peroxyl, Ascorbyl and Chromanoxyl Radicals. Biochem. Pharmacol. 1992, 44, 1637–1649. [Google Scholar] [CrossRef]
- Biewenga, G.P.; Haenen, G.R.M.M.; Bas, A. The Pharmacology of the Antioxidant Lipoic Acid. Gen. Pharmacol. 1997, 29, 315–331. [Google Scholar] [CrossRef]
- Spector, A.; Huang, R.-R.C.; Yan, G.-Z.; Wang, R.-R. Thioredoxin fragment 31-36 is reduced by dihydrolipoamide and reduces oxidized protein. Biochem. Biophys. Res. Commun. 1988, 150, 156–162. [Google Scholar] [CrossRef]
- Loftin, E.G.; Herold, L.V. Therapy and outcome of suspected alpha lipoic acid toxicity in two dogs. J. Vet. Emerg. Crit. Care 2009, 19, 501–506. [Google Scholar] [CrossRef]
- Sigel, H.; Prijs, B.; McCormick, D.B.; Shih, J.C.H. Stability of binary and ternary complexes of a-lipoate and lipoate derivatives with Mn 2+, Cu 2+, and Zn 2+ in solution. Arch. Biochem. Biophys. 1978, 187, 208–214. [Google Scholar] [CrossRef]
- Ou, P.; Tritschler, H.J.; Wolff, S.P. Thioctic (Lipoic) acid: A therapeutic metal-chelating antioxidant? Biochem. Pharmacol. 1995, 50, 123–126. [Google Scholar] [CrossRef]
- Scott, B.C.; Aruoma, O.I.; Evans, P.J.; O’Neill, C.; van der Vliet, A.; Cross, C.E.; Tritschler, H.; Halliwell, B. Lipoic and dihydrolipoic acids as antioxidants: A critical evaluation. Free Radic. Res. 1994, 20, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Sumathi, R.; Devi, V.K.; Varalakshmi, P. DL-Lipoic acid protection against cadmium induced tissue lipid peroxidation. Med. Sci. Res. 1994, 22, 23–25. [Google Scholar]
- Teichert, J.; Kern, J.; Tritschler, H.J.; Ulrich, H.; Preiss, R. Investigations on the pharmacokinetics of a-LA in healthy volunteers. Int. J. Clin. Pharmacol. Ther. 1998, 36, 625–628. [Google Scholar] [PubMed]
- Zicker, S.C.; Avila, A.; Joshi, D.K.; Gross, K.L. Pharmacokinetics of orally administered DL-a-LA in dogs. Am. J. Vet. Res. 2010, 71, 1377–1383. [Google Scholar] [CrossRef]
- Packer, L.; Witt, E.H.; Tritschler, H.J. a-LA as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Hill, A.S.; Werner, J.A.; Rogers, Q.R.; O’Neill, S.L.; Christopher, M.M. Lipoic acid is 10 times more toxic in cats than reported in humans, dogs or rats. J. Anim. Physiol. Anim. Nutr. 2004, 88, 150–156. [Google Scholar] [CrossRef]
- Peter, G.; Barbe, H.O. Untersuchungen zur absorption und verteilung der chiocrsaure als grundlage der klinischenwirksamkeit bei der behandlung der diaberischen polyneuroparhie [Studies on the absorption and distribution of thioctic acid as a basis for the clinical efficacy in the treatment of diabetic polyneuropathy]. Diabetes Stoffwechs. 1996, 5 (Suppl. 3), 12–16. [Google Scholar]
- Grunert, R.R. The effect of di-a-Lipoic Acid on heavy metal intoxication in mice and dogs. Arch. Biochem. Biophys. 1960, 86, 1990–1994. [Google Scholar] [CrossRef]
- Paetau-Robinson, I.; Brejda, J.J.; Zicker, S.C. Long-term feeding of dl-α-lipoic acid to dogs is safe. Intern. J. Appl. Res. Vet. Med. 2013, 11, 100–109. [Google Scholar]
- Sen, C.K. Nutritional biochemistry of cellular glutathione. J. Nutr. Biochem. 1997, 8, 660–672. [Google Scholar] [CrossRef]
- Sen, C.K. Oxidants and antioxidants in exercise. J. Appl. Physiol. 1995, 79, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Werner, P.; Cohen, G. Glutathione disulfide (GSSG) as a marker of oxidative injury to brain mitochondria. Ann. N. Y. Acad. Sci. 1993, 28, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.M.; Jones, J.D.; Baker, J.; Pullen, R.G. Measurement of glutathione as a marker of oxidative stress in CNS reaggregate cultures. Biochem. Soc. Trans. 1996, 24, 452S. [Google Scholar] [CrossRef]
- Neefjes, V.M.; Evelo, C.T.; Baars, L.G.; Blanco, C.E. Erythrocyte glutathione S transferase as a marker of oxidative stress at birth. Arch. Dis. Child. Fetal. Neonatal. Ed. 1999, 81, F130–F133. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Tritschler, H.J.; Packer, L. a-LA increases intracellular glutathione in a human T-lymphocyte Jurkat cell line. Biochem. Biophys. Res. Commun. 1995, 207, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K.; Roy, S.; Han, D.; Packer, L. Regulation of cellular thiols in human lymphocytes by a-LA: A flow cytometric analysis. Free Radic. Biol. Med. 1997, 22, 1241–1257. [Google Scholar] [CrossRef]
- Association of American Feed Control Officials. Official Publication of American Feed Control Officials Incorporated; Association of American Feed Control Officials: Champaign, IL, USA, 2016; pp. 153–163. [Google Scholar]
- Mohammadi, V.; Khalili, M.; Eghtesadi, S.; Dehghani, S.; Jazayeri, S.; Aghababaee, S.K.; Sabour, H.; Saberi, H.; Eghtesadi, M.; Gohari, M.R. The effect of alpha-Lipoic Acid (a-LA) supplementation on cardiovascular risk factors in men with chronic spinal cord injury: A clinical trial. Spinal Cord 2015, 53, 621–624. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Guo, Y.; Chan, L.; Guan, X. α-Lipoic acid increases energy expenditure by enhancing adenosine monophosphate–activated protein kinase–peroxisome proliferator-activated receptor-γ coactivator-1α signaling in the skeletal muscle of aged mice. Metabolism 2010, 59, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Huerta, A.E.; Navas-Carretero, S.; Prieto-Hontoria, P.L.; Martínez, J.A.; Moreno-Aliaga, M.J. Effects of α-lipoic acid and eicosapentaenoic acid in overweight and obese women during weight loss. Obesity 2015, 23, 313–321. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, P.; Wu, N.; He, B.; Lu, Y.; Li, S.; Liu, Y.; Zhao, S.; Liu, L.; Li, Y. Amelioration of Lipid Abnormalities by α-Lipoic acid Through Antioxidative and Anti-Inflammatory Effects. Obesity 2011, 19, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Handelman, G.; Marcocci, L.; Sen, C.K.; Roy, S.; Kobuchi, H.; Tritschler, H.J.; Flohé, L.; Packer, L. Lipoic acid increases de novo synthesis of glutathione by improving cystine utilization. Biofactors 1997, 6, 321–338. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Atalay, M.; Laaksonen, D.E.; Gul, M.; Roy, S.; Sen, C.K. a-Lipoic Acid Supplementation: Tissue Glutathione Homeostasis at Rest and After Exercise. J. Appl. Physiol. 1985, 86, 1191–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deneke, S.M.; Fanburg, B.L. Regulation of cellular glutathione. Am. J. Physiol. 1989, 257, L163–L173. [Google Scholar] [CrossRef]
- Srivastava, S.K. Metabolism of red cell glutathione. Exp. Eye Res. 1971, 11, 294–305. [Google Scholar] [CrossRef]
- Anderson, M.E.; Meister, A. Dynamic State of Glutathione in Blood Plasma. J. Biol. Chem. 1980, 255, 9530–9533. [Google Scholar] [CrossRef]
- Griffith, O.W.; Meister, A. Glutathione: Interorgan translocation, turnover, and metabolism. Proc. Natl. Acad. Sci. USA 1979, 76, 268–272. [Google Scholar] [CrossRef] [Green Version]
- Griffith, O.W.; Novogrodsky, A.; Meister, A. Translocation of glutathione from lymphoid cells that have markedly different gamma-glutamyl transpeptidase activities. Proc. Natl. Acad. Sci. USA 1979, 76, 2249–2252. [Google Scholar] [CrossRef] [Green Version]
- Meister, A.; Griffith, O.W.; Novogrodsky, A.; Tate, S.S. Direct evidence for inter-organ transport of glutathione and that the non-filtration renal mechanism for glutathione utilization involves γ-glutamyl transpeptidase. Ciba Found. Symp. 1980, 72, 135–161. [Google Scholar]
- Wang, L.; Ahn, Y.J.; Asmis, R. Sexual dimorphism in glutathione metabolism and glutathione-dependent responses. Redox Biol. 2020, 31, 101410. [Google Scholar] [CrossRef]
| Patient Characteristics | Overall | By Group | |||
|---|---|---|---|---|---|
| Control | Food 1 | Food 2 | Food 3 | ||
| Number of Animals | 80 | 20 | 20 | 20 | 20 |
| Male | 3 | 1 | 1 | 1 | 0 |
| Female | 1 | 0 | 1 | 0 | 0 |
| Neutered | 41 | 9 | 9 | 9 | 14 |
| Spayed | 35 | 10 | 9 | 10 | 6 |
| Mean age (yr) | 10.2 ± 1.8 | 9.8 ± 2.1 | 10.0 ± 1.6 | 10.6 ± 1.8 | 10.5 ± 1.7 |
| Mean weight (kg) | 14.6 ± 1.0 | 16.3 ± 1.0 | 14.7 ± 1.0 | 14.3 ± 1.0 | 12.9 ± 1.0 |
| Mean serum Vitamin E (μg/mL) | 33.8 ± 8.2 | 35.0 ± 8.8 | 33.9 ± 9.0 | 34.0 ± 8.1 | 32.2 ± 7.1 |
| Test | Food Group (If Applicable) | Description of Adverse Event | No of Animals |
|---|---|---|---|
| Phase 1 | N/A | Loose stool observed and resolved | 1 |
| Phase 1 | N/A | Perineal hernia, not study-related | 1 |
| Phase 1 | N/A | Hemangiosarcoma, not study-related | 1 |
| Phase 2 | Food 1 | Vomiting/anorexia—hepatopathy, resolved, completed study | 1 |
| Phase 2 | Food 3 | Coughing × 2 days—age-related cardiac disease, not food-related. Started on diuretics, completed study | 1 |
| Phase 2 | Food 3 | Anima died, due to anaplastic sarcoma, not study-related | 1 |
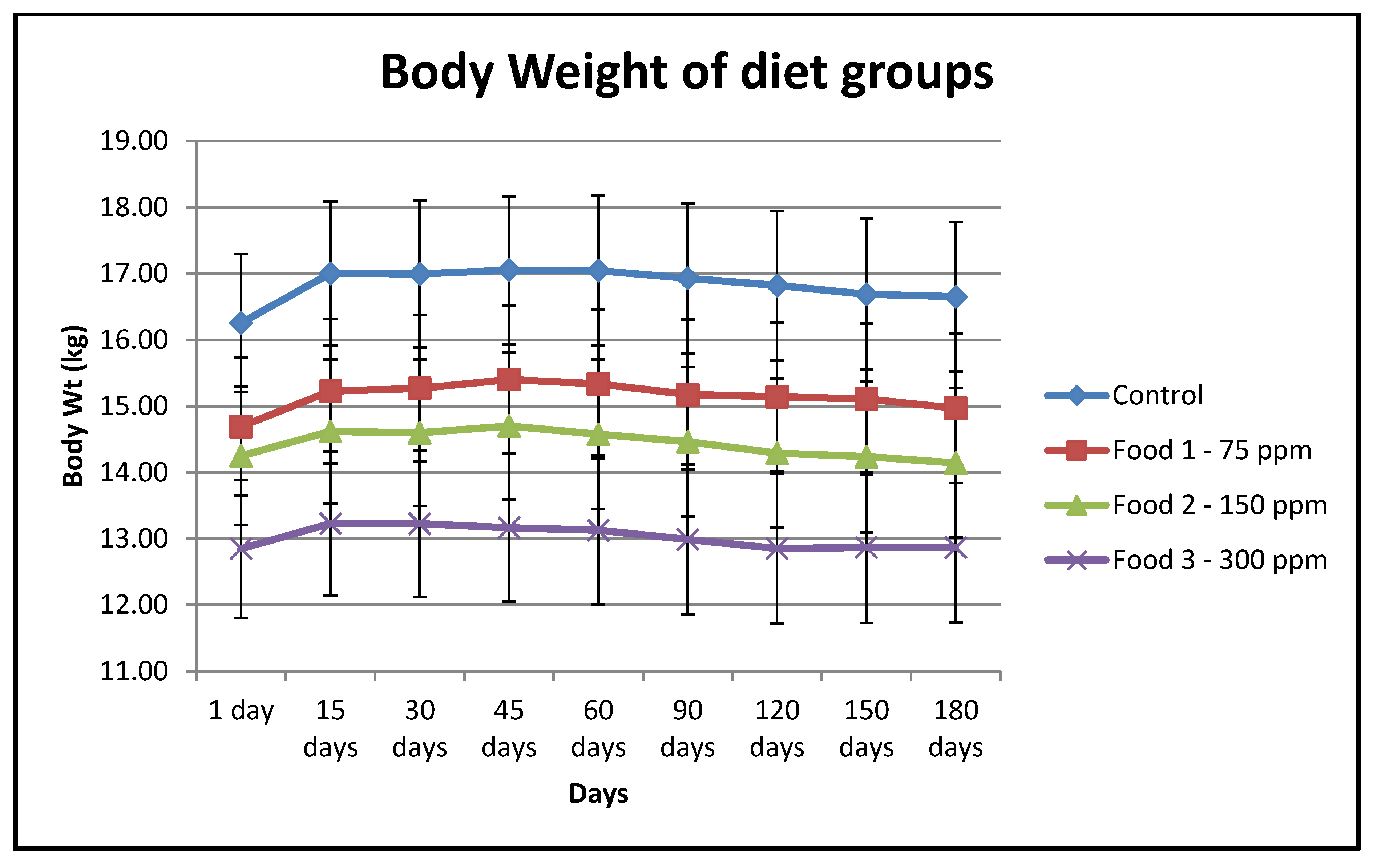
| Food | n | Mean Body wt (In kgs) of Dogs in Each Group on Day 1 | SE | Mean Body wt (In kgs) of Dogs in Each Group on Day 180 | SE | Adjusted p-Value of Mean Body wt Day 1 vs. 180 |
|---|---|---|---|---|---|---|
| Control | 20 | 16.26 | 1.04 | 16.65 | 1.13 | 0.53 |
| Food 1 | 20 | 14.69 | 1.04 | 14.97 | 1.13 | 0.68 |
| Food 2 | 20 | 14.25 | 1.04 | 14.14 | 1.13 | 0.96 |
| Food 3 | 19 | 12.85 | 1.04 | 12.87 | 1.13 | 1.00 |
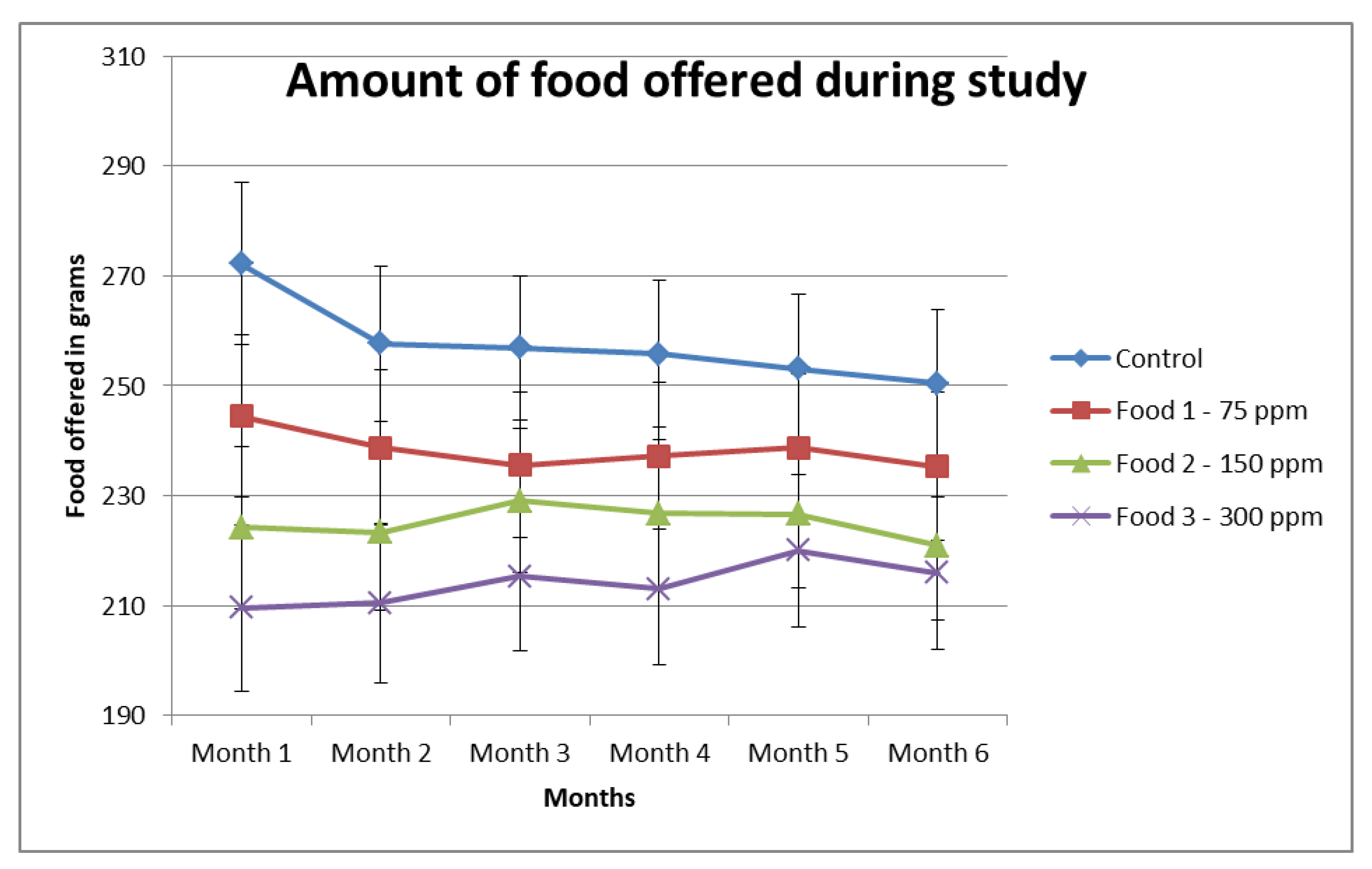
| Food | Period/ 30 Days | n | Mean (g) | SE | Difference vs. Period 1 | Adjusted p-Value vs. Period 1 |
|---|---|---|---|---|---|---|
| Control | 1 | 20 | 272.2 | 14.8 | ||
| Control | 2 | 20 | 257.7 | 14.1 | −14.5 | 0.22 |
| Control | 3 | 20 | 256.9 | 13.1 | −15.4 | 0.28 |
| Control | 4 | 20 | 255.8 | 13.4 | −16.4 | 0.27 |
| Control | 5 | 20 | 253.1 | 13.5 | −19.2 | 0.27 |
| Control | 6 | 20 | 250.4 | 13.5 | −21.8 | 0.27 |
| Food 1 | 1 | 20 | 244.4 | 14.8 | ||
| Food 1 | 2 | 20 | 238.7 | 14.1 | −5.7 | 0.49 |
| Food 1 | 3 | 20 | 235.6 | 13.1 | −8.8 | 0.46 |
| Food 1 | 4 | 20 | 237.2 | 13.4 | −7.2 | 0.55 |
| Food 1 | 5 | 20 | 238.7 | 13.5 | −5.7 | 0.71 |
| Food 1 | 6 | 20 | 235.3 | 13.5 | −9.1 | 0.57 |
| Food 2 | 1 | 20 | 224.2 | 14.8 | ||
| Food 2 | 2 | 20 | 223.3 | 14.1 | −0.9 | 0.99 |
| Food 2 | 3 | 20 | 229.1 | 13.1 | 4.9 | 0.68 |
| Food 2 | 4 | 20 | 226.8 | 13.4 | 2.6 | 0.90 |
| Food 2 | 5 | 20 | 226.7 | 13.5 | 2.5 | 0.94 |
| Food 2 | 6 | 20 | 220.9 | 13.5 | −3.3 | 0.91 |
| Food 3 | 1 | 20 | 215.1 | 14.9 | ||
| Food 3 | 2 | 20 | 213.5 | 14.1 | −1.6 | 0.93 |
| Food 3 | 3 | 20 | 219.9 | 13.2 | 4.8 | 0.71 |
| Food 3 | 4 | 20 | 214.0 | 13.4 | −1.1 | 0.99 |
| Food 3 | 5 | 19 | 220.9 | 13.5 | 5.8 | 0.75 |
| Food 3 | 6 | 19 | 216.8 | 13.5 | 1.8 | 0.99 |
| Slope | 95% Confidence Interval | |||||
|---|---|---|---|---|---|---|
| Matrix | Analyte | Estimate | SE | p-Value | Lower | Upper |
| Plasma | GSH | −0.011 | 0.037 | 0.774 | −0.084 | 0.063 |
| Plasma | GSSG | −0.014 | 0.041 | 0.726 | −0.095 | 0.066 |
| Plasma | GSH/GSSG Ratio | 0.005 | 0.031 | 0.880 | −0.057 | 0.067 |
| Plasma | Total glutathione | −0.014 | 0.036 | 0.709 | −0.086 | 0.059 |
| RBC Lysate | GSH | 0.049 | 0.021 | 0.024 | 0.01 | 0.092 |
| RBC Lysate | GSSG | 0.043 | 0.034 | 0.203 | −0.024 | 0.110 |
| RBC Lysate | GSH/GSSG Ratio | 0.006 | 0.029 | 0.846 | −0.052 | 0.063 |
| RBC Lysate | Total glutathione | 0.047 | 0.021 | 0.029 | 0.005 | 0.089 |
| z | Slope | 95% Confidence Interval | ||||
|---|---|---|---|---|---|---|
| Matrix | Analyte | Estimate | SE | p Value | Lower | Upper |
| Plasma | GSH | 0.021 | 0.054 | 0.694 | −0.086 | 0.129 |
| Plasma | GSSG | 0.008 | 0.041 | 0.843 | −0.073 | 0.089 |
| Plasma | GSH/GSSG Ratio | 0.013 | 0.046 | 0.775 | −0.079 | 0.106 |
| Plasma | Total glutathione | 0.016 | 0.049 | 0.74 | −0.080 | 0.113 |
| RBC Lysate | GSH | 0.049 | 0.021 | 0.022 | 0.007 | 0.092 |
| RBC Lysate | GSSG | 0.064 | 0.031 | 0.044 | 0.002 | 0.126 |
| RBC Lysate | GSH/GSSG Ratio | −0.015 | 0.108 | 0.891 | −0.229 | 0.200 |
| RBC Lysate | Total glutathione | 0.050 | 0.022 | 0.023 | 0.007 | 0.093 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anthony, R.M.; MacLeay, J.M.; Jewell, D.E.; Brejda, J.J.; Gross, K.L. Alpha-Lipoic Acid Is an Effective Nutritive Antioxidant for Healthy Adult Dogs. Animals 2021, 11, 274. https://doi.org/10.3390/ani11020274
Anthony RM, MacLeay JM, Jewell DE, Brejda JJ, Gross KL. Alpha-Lipoic Acid Is an Effective Nutritive Antioxidant for Healthy Adult Dogs. Animals. 2021; 11(2):274. https://doi.org/10.3390/ani11020274
Chicago/Turabian StyleAnthony, Reshma M., Jennifer M. MacLeay, Dennis E. Jewell, John J. Brejda, and Kathy L. Gross. 2021. "Alpha-Lipoic Acid Is an Effective Nutritive Antioxidant for Healthy Adult Dogs" Animals 11, no. 2: 274. https://doi.org/10.3390/ani11020274
APA StyleAnthony, R. M., MacLeay, J. M., Jewell, D. E., Brejda, J. J., & Gross, K. L. (2021). Alpha-Lipoic Acid Is an Effective Nutritive Antioxidant for Healthy Adult Dogs. Animals, 11(2), 274. https://doi.org/10.3390/ani11020274