
Population Density and Driving Factors of North China Leopards in Tie Qiao Shan Nature Reserve


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
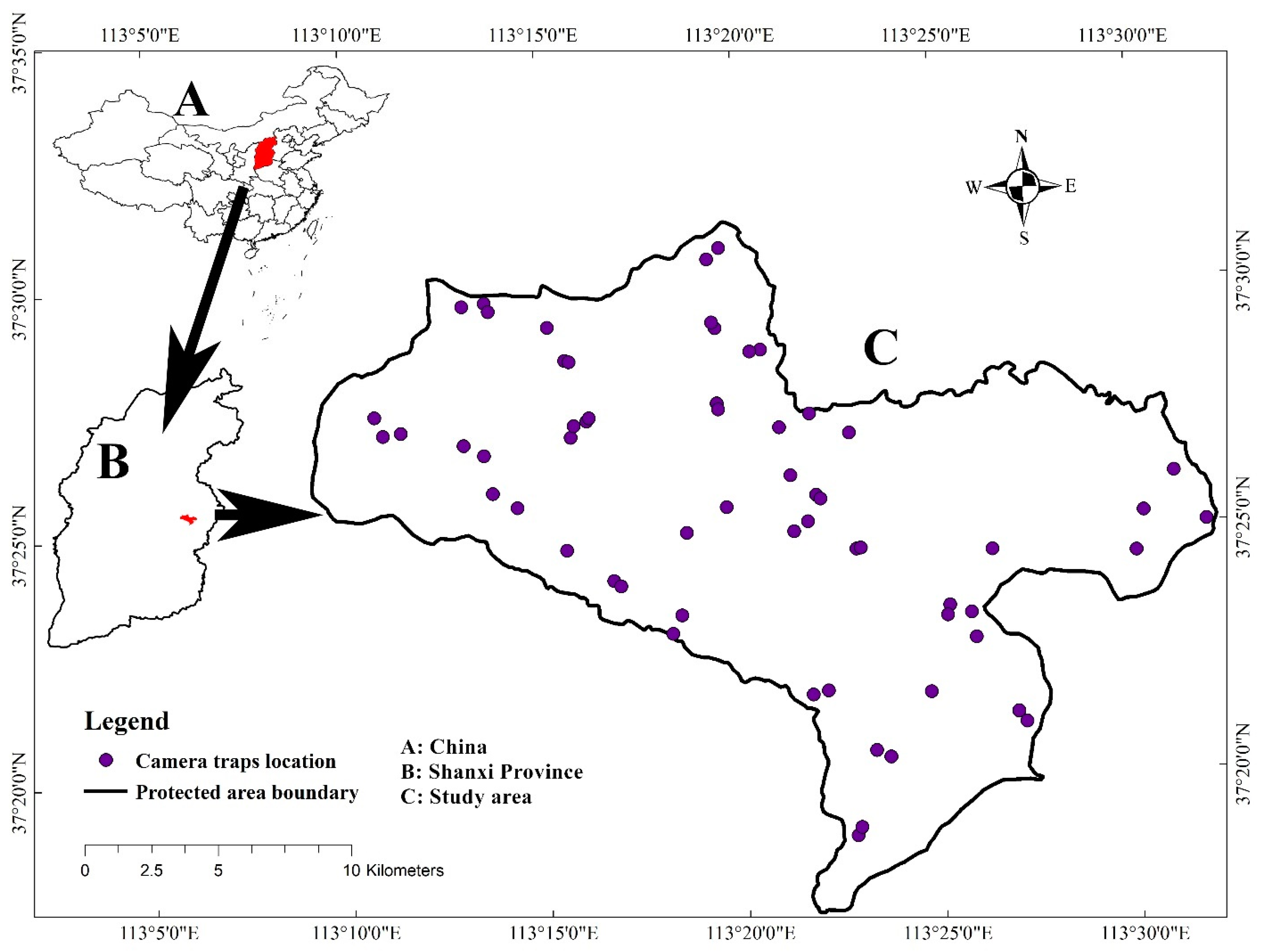
2.1. Study Area
2.2. Camera Trap Erection
2.3. Data Analysis
3. Results
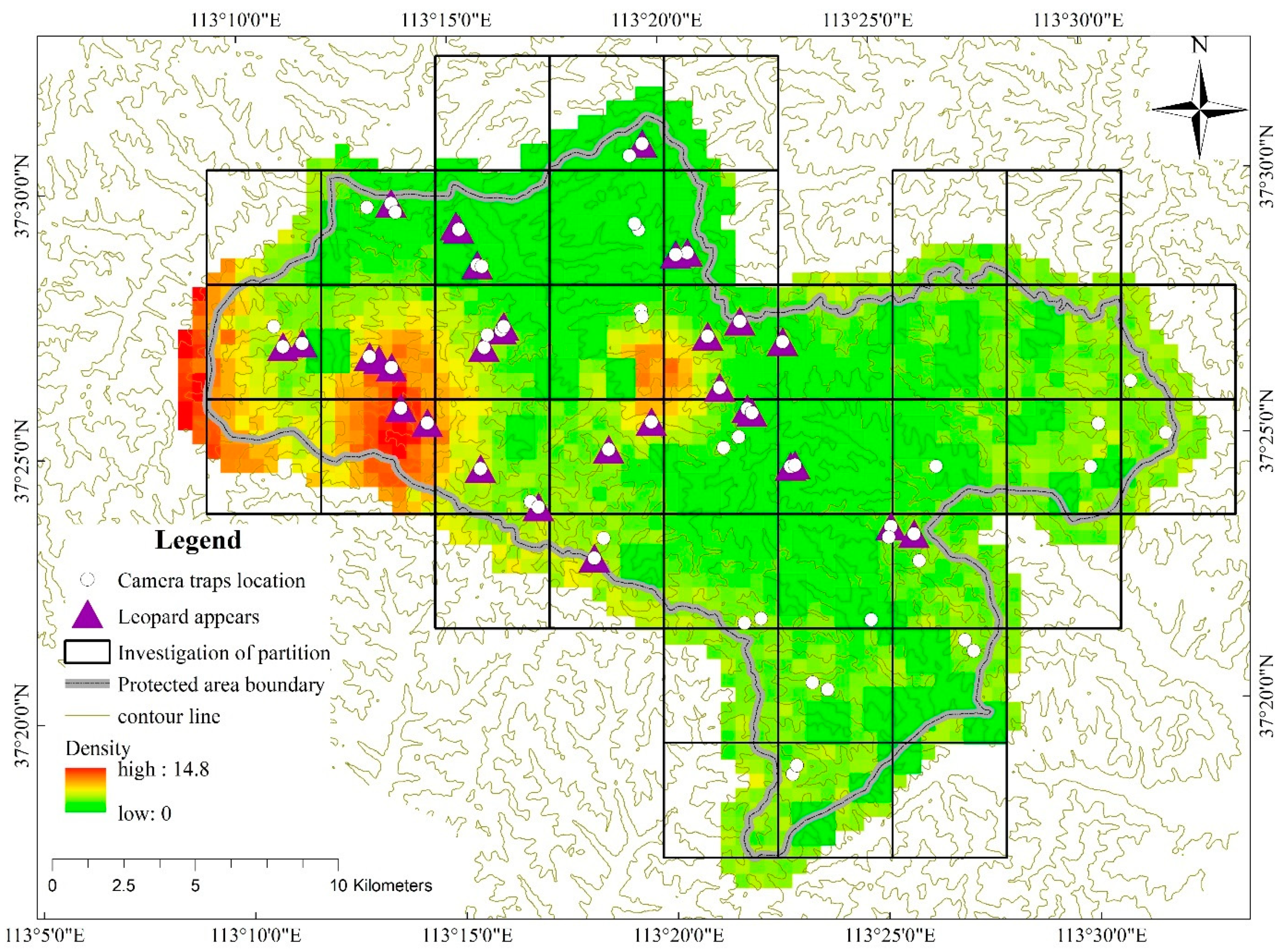
3.1. Population Density and Spatial Distribution of North China Leopard
3.2. The Relative Abundance Index and Distribution of Wild Boar and Roe Deer
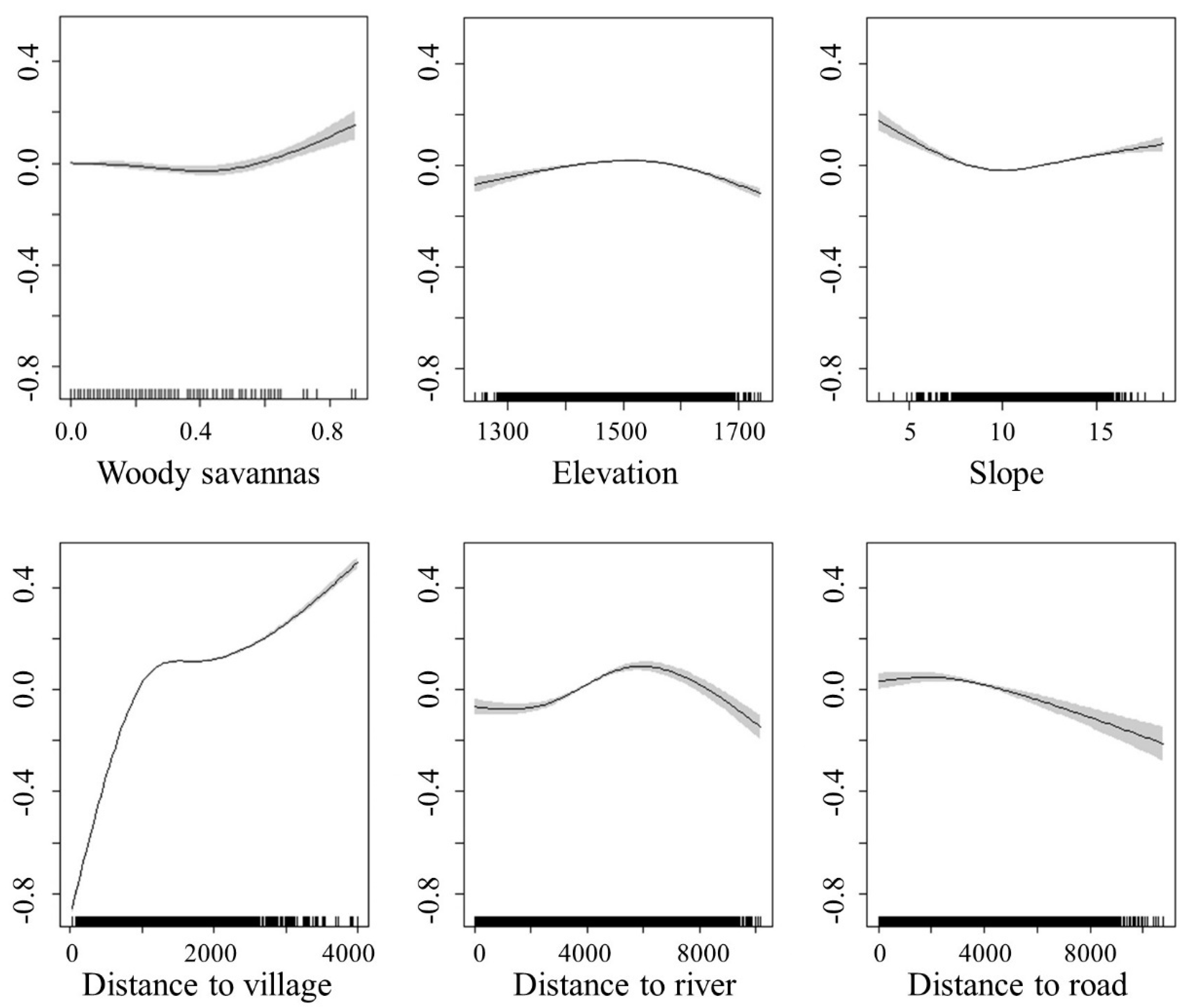
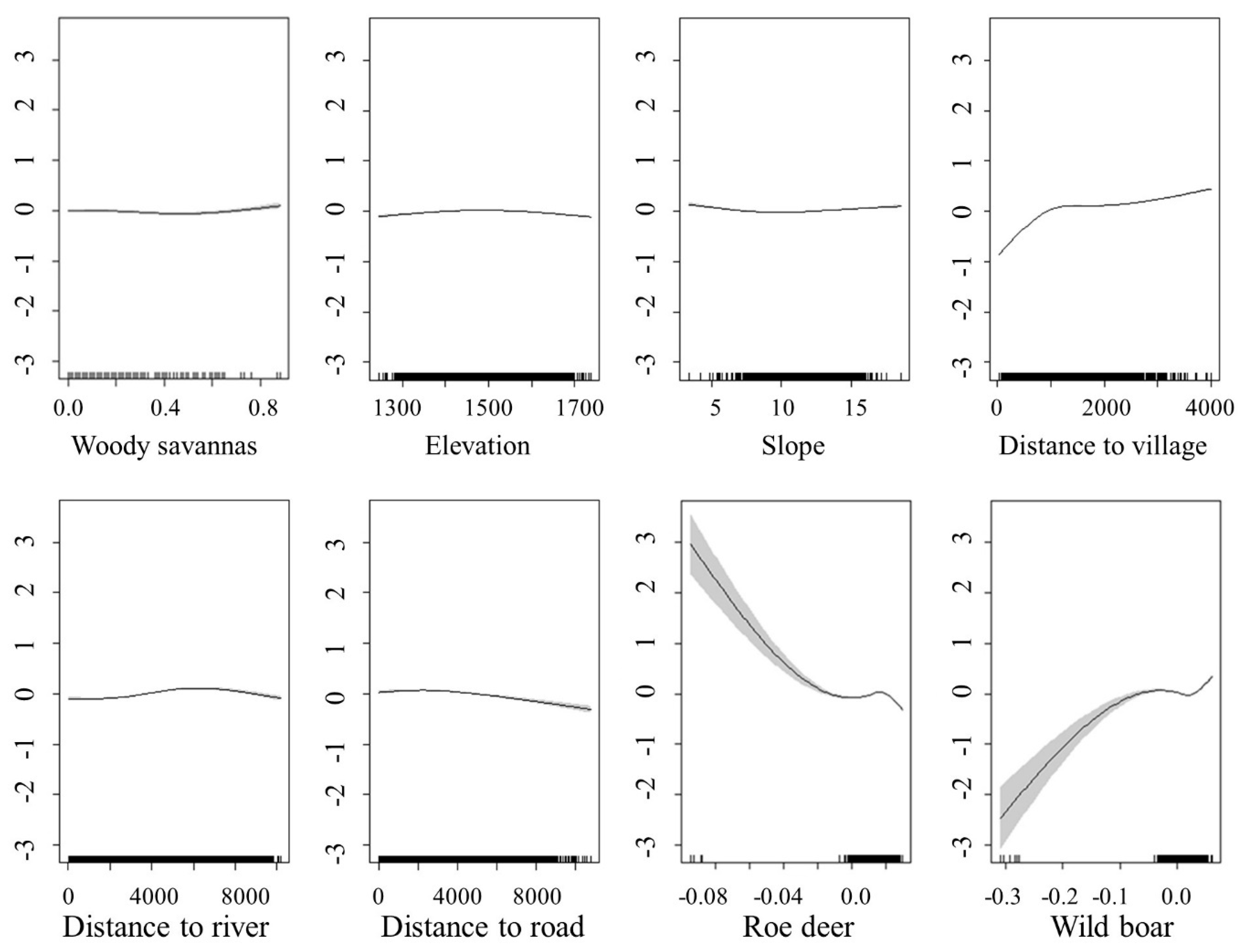
3.3. The Relationship between the Spatial Distribution of North China Leopard Density and Its Prey and Other Environmental Factors
4. Discussion
4.1. Population Size and Density Distribution of North China Leopard
4.2. The Relationship between the Distribution of North China Leopard Prey and the Population Density Distribution of North China Leopard
4.3. The Relationship between Environmental Factors and Human Factors and the Population Density Distribution of North China Leopards
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobson, A.P.; Gerngross, P.; Lemeris, J.R., Jr.; Schoonover, R.F.; Anco, C.; Breitenmoser-Wursten, C.; Durant, S.M.; Farhadinia, M.S.; Henschel, P.; Kamler, J.F.; et al. Leopard (Panthera Pardus) Status, Distribution, and The Research Efforts Across Its Range. PeerJ 2016, 4, e1974. [Google Scholar] [CrossRef] [Green Version]
- Simcharoen, S.; Barlow, A.C.D.; Simcharoen, A.; Smith, J.L.D. Home Range Size and Daytime Habitat Selection of Leopards in Huai Kha Khaeng Wildlife Sanctuary, Thailand. Biol. Conserv. 2008, 141, 2242–2250. [Google Scholar] [CrossRef]
- Mondal, K.; Bhattacharjee, S.; Gupta, S.; Sankar, K.; Qureshi, Q. Home Range and Resource Selection of ‘Problem’ Leopards Trans-Located to Forested Habitat. Curr. Sci. 2013, 105, 338–345. [Google Scholar]
- Laguardia, A.; Kamler, J.F.; Li, S.; Zhang, C.; Zhou, Z.; Shi, K. The Current Distribution and Status of Leopards Panthera Pardus in China. Oryx 2015, 51, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Hale, R.; Swearer, S.E. When Good Animals Love Bad Restored Habitats: How Maladaptive Habitat Selection Can Constrain Restoration. J. Appl. Ecol. 2017, 54, 1478–1486. [Google Scholar] [CrossRef]
- Morris, D.W. Toward an Ecological Synthesis: A Case for Habitat Selection. Oecologia 2003, 136, 1–13. [Google Scholar] [CrossRef]
- Mondal, K.; Gupta, S.; Qureshi, Q.; Sankar, K. Prey Selection and Food Habits of Leopard (Panthera pardus Fusca) in Sariska Tiger Reserve, Rajasthan, India. Mammalia 2011, 75, 201–205. [Google Scholar] [CrossRef]
- Hayward, M.W.; Henschel, P.; O’Brien, J.; Hofmeyr, M.; Balme, G.; Kerley, G.I.H. Prey Preferences of the Leopard (Panthera Pardus). J. Zool. 2006, 270, 298–313. [Google Scholar] [CrossRef]
- Aryal, A.; Brunton, D.; Ji, W.H.; Raubenheimer, D. Blue Sheep in The Annapurna Conservation Area, Nepal: Habitat Use, Population Biomass and Their Contribution to The Carrying Capacity of Snow Leopards. Integr. Zool. 2014, 9, 34–45. [Google Scholar] [CrossRef]
- Cushman, S.A. Effects of Habitat Loss and Fragmentation on Amphibians: A Review and Prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, D.F.; Oliveira, E.S.; Alves, R.R.N. Conflicts between Humans and Terrestrial Vertebrates: A Global Review. Trop. Conserv. Sci. 2018, 11, 15. [Google Scholar] [CrossRef]
- Athreya, V.; Odden, M.; Linnell, J.D.C.; Krishnaswamy, J.; Karanth, U. Big Cats in Our Backyards: Persistence of Large Carnivores in A Human Dominated Landscape in India. PLoS ONE 2013, 8, e57872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- du Preez, B.; Loveridge, A.J.; Macdonald, D.W. Making The Best of Camera-Trap Surveys in an Imperfect World: A Reply to Balme et al. Biol. Conserv. 2014, 179, 146–147. [Google Scholar] [CrossRef]
- Hiby, L.; Lovell, P.; Patil, N.; Kumar, N.S.; Gopalaswamy, A.M.; Karnath, K.U. A tiger Cannot Change Its Stripes: Using A Three-Dimensional Model to Match Images of Living Tigers and Tiger Skins. Biol. Lett. 2009, 5, 383–386. [Google Scholar] [CrossRef]
- Duarte, A. Camera Trapping for Wildlife Research. J. Wildl. Manag. 2017, 81, 1125. [Google Scholar] [CrossRef]
- Silver, S.C.; Ostro, L.E.T.; Marsh, L.K.; Maffei, L.; Noss, A.J.; Kelly, M.J.; Wallace, R.B.; Gomez, H.; Ayala, G. The Use of Camera Traps for Estimating Jaguar Panthera Onca Abundance and Density Using Capture/Recapture Analysis. Oryx 2004, 38, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, T.V.; Kovach, A.I. Spatially Explicit Abundance Estimation of a Rare Habitat Specialist: Implications for Secr Study Design. Ecosphere 2018, 9, 17. [Google Scholar] [CrossRef]
- Gopalaswamy, A.M.; Royle, J.A.; Hines, J.E.; Singh, P.; Jathanna, D.; Kumar, N.S.; Karanth, K.U. Program SPACECAP: Software for Estimating Animal Density Using Spatially Explicit Capture-Recapture Models. Methods Ecol. Evol. 2012, 3, 1067–1072. [Google Scholar] [CrossRef]
- Hopkins, H.L.; Kennedy, L.L. An Assessment of Indices of Relative and Absolute Abundance for Monitoring Populations of Small Mammals. Wildl. Soc. Bull. 2004, 32, 1289–1296. [Google Scholar] [CrossRef]
- Jenks, K.E.; Chanteap, P.; Damrongchainarong, K.; Cutter, P.; Cutter, P.; Redford, T.; Lynam, A.J.; Howard, J.; Leimgruber, P. Using relative Abundance Indices from Camera-Trapping to Test Wildlife Conservation Hypotheses—An Example from Khao Yai National Park, Thailand. Trop. Conserv. Sci. 2011, 4, 113–131. [Google Scholar] [CrossRef] [Green Version]
- Stasinopoulos, D.M.; Rigby, R.A. Generalized Additive Models for Location Scale and Shape (GAMLSS) in R. J. Stat. Softw. 2007, 23, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Shi, Q.; Wang, G.; Li, Z.; Sun, Q.; Hua, Y.; Jiang, G. Spatial Distribution Drivers of Amur Leopard Density in Northeast China. Biol. Conserv. 2015, 191, 258–265. [Google Scholar] [CrossRef]
- Yang, H.; Xie, B.; Zhao, G.; Gong, Y.; Mou, P.; Ge, J.; Feng, L. Elusive Cats in Our Backyards: Persistence of The North Chinese Leopard (Panthera Pardus Japonensis) in A Human-Dominated Landscape in Central China. Integr. Zool. 2020. [Google Scholar] [CrossRef]
- Mondal, K.; Sankar, K.; Qureshi, Q. Factors Influencing The Distribution of Leopard in A Semiarid Landscape of Western India. Acta Theriol. 2013, 58, 179–187. [Google Scholar] [CrossRef]
- Roberge, J.M.; Angelstam, P. Usefulness of the Umbrella Species Concept as A Conservation Tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Eklund, J.; Cabeza, M. Quality of Governance and Effectiveness of Protected Areas: Crucial Concepts for Conservation Planning. Ann. N. Y. Acad. Sci. 2017, 1399, 27–41. [Google Scholar] [CrossRef]
- Keuling, O.; Stier, N.; Roth, M. Commuting, Shifting or Remaining? Different Spatial Utilisation Patterns of Wild Boar Sus Scrofa L. in Forest and Field Crops during Summer. Mamm. Biol. 2009, 74, 145–152. [Google Scholar] [CrossRef]
- Acevedo, P.; Escudero, M.A.; Munoz, R.; Gortazar, C. Factors Affecting Wild Boar Abundance Across an Environmental Gradient in Spain. Acta Theriol. 2006, 51, 327–336. [Google Scholar] [CrossRef]
- Mysterud, A.; Tryjanowski, P.; Panek, M.; Pettorelli, N.; Stenseth, N.C. Inter-Specific Synchrony of Two Contrasting Ungulates: Wild Boar (Sus Scrofa) and Roe Deer (Capreolus Capreolus). Oecologia 2007, 151, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Strampelli, P.; Andresen, L.; Everatt, K.T.; Somers, M.J.; Rowcliffe, J.M. Habitat Use Responses of the African Leopard in A Human-Disturbed Region of Rural Mozambique. Mamm. Biol. 2018, 89, 14–20. [Google Scholar] [CrossRef]
- Henschel, P.; Hunter, L.T.B.; Coad, L.; Abernethy, K.A.; Muehlenberg, M. Leopard Prey Choice in The Congo Basin Rainforest Suggests Exploitative Competition with Human Bushmeat Hunters. J. Zool. 2011, 285, 11–20. [Google Scholar] [CrossRef]
- Lyngdoh, S.; Shrotriya, S.; Goyal, S.P.; Clements, H.; Hayward, M.W.; Habib, B. Prey Preferences of the Snow Leopard (Panthera Uncia): Regional Diet Specificity Holds Global Significance for Conservation. PLoS ONE 2014, 9, e88349. [Google Scholar] [CrossRef]
- Bai, D.F.; Chen, P.J.; Atzeni, L.; Cering, L.; Li, Q.; Shi, K. Assessment of Habitat Suitability of the Snow Leopard (Panthera Uncia) in Qomolangma National Nature Reserve Based on Maxent Modeling. Zool. Res. 2018, 39, 373–386. [Google Scholar] [CrossRef]
- Pu, Z.; Zhang, F.; Gu, Y.; Liang, X.; Xing, S. Conservation Status and Priority Conservation Areas of Natural Vegetation in Northeast China. Chin. J. Ecol. 2019, 38, 2821–2832. [Google Scholar]
- Macdonald, D.W.; Bothwell, H.M.; Hearn, A.J.; Cheyne, S.M.; Haidir, I.; Hunter, L.T.B.; Kaszta, Z.; Linkie, M.; Macdonald, E.A.; Ross, J.; et al. Multi-Scale Habitat Selection Modeling Identifies Threats and Conservation Opportunities for The Sunda Clouded Leopard (Neofelis Diardi). Biol. Conserv. 2018, 227, 92–103. [Google Scholar] [CrossRef]
- Penjor, U.; Macdonald, D.W.; Wangchuk, S.; Tandin, T.; Tan, C.K.W. Identifying Important Conservation Areas for The Clouded Leopard Neofelis Nebulosa in A Mountainous Landscape: Inference from Spatial Modeling Techniques. Ecol. Evol. 2018, 8, 4278–4291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haidir, I.A.; Macdonald, D.W.; Wong, W.-M.; Lubis, M.I.; Linkie, M. Population Dynamics of Threatened Felids in Response to Forest Cover Change in Sumatra. PLoS ONE 2020, 15, e0236144. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description of the Habitat Factor | Data Type | Unit |
|---|---|---|---|
| Elevation | 30 m resolution DEM conversion 0.5 km resolution elevation grid | Continuous | m |
| Slope | 30 m resolution DEM conversion 0.5 km resolution slope grid | Continuous | m |
| Aspect | 30 m resolution DEM conversion 0.5 km resolution aspect grid | Continuous | m |
| Distance to village | Distance from the central point of each pixel to the nearest village | Continuous | m |
| Distance to road | Distance from the central point of each pixel to the nearest road | Continuous | m |
| Distance to river | Distance from the central point of each pixel to the nearest river | Continuous | m |
| Net production | Difference between how much carbon dioxide is taken in by plants compared to how much is put out by them; this difference is total amount of carbon dioxide taken in by plants, called net primary productivity; extract in raster layer | Continuous | gCm−2 yr−1 |
| Mixed forest | Mixed forest area ratio of each pixel | Continuous | Ratio |
| Woody savanna | Woody savanna area ratio of each pixel | Continuous | Ratio |
| Parameter | Mean | SD | 5% | 95% |
|---|---|---|---|---|
| ψ | 4782.32 | 1689.66 | 3133.31 | 6741.81 |
| σ | 0.03 | 0.01 | 0.01 | 0.04 |
| β | 1.38 | 0.38 | 0.69 | 2.18 |
| p0 | 0.43 | 0.11 | 0.25 | 0.66 |
| N | 17.98 | 3.45 | 12 | 24 |
| D | 4.23 | 0.81 | 2.82 | 5.64 |
| p1 | 0.03 | 0.01 | 0.01 | 0.04 |
| p2 | 0.01 | 0.02 | 0.06 | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Zaman, M.; Wang, M.; Vitekere, K.; Ma, J.; Jiang, G. Population Density and Driving Factors of North China Leopards in Tie Qiao Shan Nature Reserve. Animals 2021, 11, 429. https://doi.org/10.3390/ani11020429
Zhu M, Zaman M, Wang M, Vitekere K, Ma J, Jiang G. Population Density and Driving Factors of North China Leopards in Tie Qiao Shan Nature Reserve. Animals. 2021; 11(2):429. https://doi.org/10.3390/ani11020429
Chicago/Turabian StyleZhu, Mengyan, Muhammad Zaman, Meng Wang, Kasereka Vitekere, Jianzhang Ma, and Guangshun Jiang. 2021. "Population Density and Driving Factors of North China Leopards in Tie Qiao Shan Nature Reserve" Animals 11, no. 2: 429. https://doi.org/10.3390/ani11020429