
Occurrence and Genetic Diversity of Protist Parasites in Captive Non-Human Primates, Zookeepers, and Free-Living Sympatric Rats in the Córdoba Zoo Conservation Centre, Southern Spain

 , , , , , ,
, , , , , ,  ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Epidemiological Questionnaire
2.4. DNA Extraction and Purification
2.5. Molecular Detection and Characterization of Giardia Duodenalis
2.6. Molecular Detection and Characterization of Cryptosporidium spp.
2.7. Molecular Detection and Characterization of Blastocystis sp.
2.8. Molecular Detection and Characterization of Enterocytozoon bieneusi
2.9. Molecular Differential Detection of Entamoeba histolytica and Entamoeba dispar
2.10. Molecular Detection of Balantioides coli
2.11. Molecular Detection of Troglodytella spp.
2.12. Molecular Detection of Leishmania spp.
2.13. Statistical Analysis
3. Results
3.1. Prevalence and Molecular Characterization of Enteroparasites in Captive Non-Human Primates
3.2. Prevalence and Molecular Characterization of Enteroparasites in Humans
3.3. Prevalence and Molecular Characterization of Enteroparasites and Leishmania spp. in Rats
3.4. Molecular-Based Evidence of Zoonotic Transmission
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DuPont, H.L. Persistent diarrhea: A clinical review. JAMA 2016, 315, 2712–2723. [Google Scholar] [CrossRef]
- Hemphill, A.; Müller, N.; Müller, J. Comparative pathobiology of the intestinal protozoan parasites Giardia lamblia, En-tamoeba histolytica, and Cryptosporidium parvum. Pathogens 2019, 8, 116. [Google Scholar] [CrossRef]
- Ajjampur, S.S.; Tan, K.S. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitol. Int. 2016, 65, 772–779. [Google Scholar] [CrossRef]
- Li, W.; Feng, Y.; Santin, M. Host Specificity of Enterocytozoon bieneusi and Public Health Implications. Trends Parasitol. 2019, 35, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Cacciò, S.M. Zoonotic potential of Giardia. Int. J. Parasitol. 2013, 43, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium species in humans and animals: Current understanding and research needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef]
- Li, W.; Feng, Y.; Zhang, L.; Xiao, L. Potential impacts of host specificity on zoonotic or interspecies transmission of Enterocytozoon bieneusi. Infect. Genet. Evol. 2019, 75, 104033. [Google Scholar] [CrossRef] [PubMed]
- Hublin, J.S.Y.; Maloney, J.G.; Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Widmer, G.; Köster, P.C.; Carmena, D. Cryptosporidium hominis infections in non-human animal species: Revisiting the concept of host specificity. Int. J. Parasitol. 2020, 50, 253–262. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s Box: Blastocystis Subtypes Revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef]
- Maloney, J.G.; Molokin, A.; Da Cunha, M.J.R.; Cury, M.C.; Santin, M. Blastocystis subtype distribution in domestic and captive wild bird species from Brazil using next generation amplicon sequencing. Parasite Epidemiol. Control 2020, 9, e00138. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Jiang, W.; Roellig, D.M.; Wan, Z.; Li, N.; Guo, Y.; Feng, Y.; Xiao, L. Characterizations of Enterocytozoon bieneusi at new genetic loci reveal a lack of strict host specificity among common genotypes and the existence of a canine-adapted Enterocytozoon species. Int. J. Parasitol. 2021, 51, 215–223. [Google Scholar] [CrossRef]
- Gómez, M.S.; Gracenea, M.; Gosalbez, P.; Feliu, C.; Enseñat, C.; Hidalgo, R. Detection of oocysts of Cryptosporidium in several species of monkeys and in one prosimian species at the Barcelona zoo. Parasitol. Res. 1992, 78, 619–620. [Google Scholar] [CrossRef]
- Soledad Gómez, M.; Gracenea, M.; Montoliu, I.; Feliu, C.; Monleon, A.; Fernandez, J.; Enseñat, C. Intestinal parasitism—protozoa and helminths—in primates at the Barcelona Zoo. J. Med. Primatol. 1996, 25, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.S.; Torres, J.; Gracenea, M.; Fernandez-Morán, J.; Gonzalez-Moreno, O. Further report on Cryptosporidium in Barcelona zoo mammals. Parasitol. Res. 2000, 86, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Gracenea, M.; Gómez, M.S.; Torres, J.; Carné, E.; Fernández-Morán, J. Transmission dynamics of Cryptosporidium in primates and herbivores at the Barcelona zoo: A long-term study. Veter. Parasitol. 2002, 104, 19–26. [Google Scholar] [CrossRef]
- Pérez Cordón, G.; Hitos Prados, A.; Romero, D.; Sánchez Moreno, M.; Pontes, A.; Osuna, A.; Rosales, M.J. Intestinal parasitism in the animals of the zoological garden “Peña Escrita” (Almuñecar, Spain). Vet. Parasitol. 2008, 156, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Díaz, R.A.; Sansano-Maestre, J.; Martínez-Herrero, M.D.C.; Ponce-Gordo, F.; Gómez-Muñoz, M.T. Occurrence and genetic characterization of Giardia duodenalis from captive nonhuman primates by multi-locus sequence analysis. Parasitol. Res. 2011, 109, 539–544. [Google Scholar] [CrossRef]
- Ponce-Gordo, F.; García-Rodríguez, J.J. Balantioides coli. Res. Veter. Sci. 2020, S0034–5288, 31066–31073. [Google Scholar] [CrossRef]
- Pomajbíková, K.; Petrželková, K.J.; Profousová, I.; Petrášová, J.; Kišidayová, S.; Váradyová, Z.; Modrý, D. A survey of entodiniomorphid ciliates in chimpanzees and bonobos. Am. J. Phys. Anthr. 2009, 142, 42–48. [Google Scholar] [CrossRef]
- UNE 16636:2015. Available online: https://www.aenor.com/normas-y-libros/buscador-de-normas/une/?Tipo=N&c=N0055762 (accessed on 6 February 2021).
- Mayer, J.; Mans, C. Rodents. In Exotic Animal Formulary, 5th ed.; Carpenter, J.W., Marion, C.J., Eds.; Elsevier: St. Louis, MO, USA, 2018; pp. 673–675. [Google Scholar]
- Verweij, J.J.; Schinkel, J.; Laeijendecker, D.; van Rooyen, M.A.; van Lieshout, L.; Polderman, A.M. Real-time PCR for the detection of Giardia lamblia. Mol. Cell. Probes 2003, 17, 223–225. [Google Scholar] [CrossRef]
- Read, C.M.; Monis, P.T.; Thompson, R.C. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydro-genase locus using PCR-RFLP. Infect. Genet. Evol. 2004, 4, 125–130. [Google Scholar] [CrossRef]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic heterogeneity at the beta-giardin locus among human and animal isolates of Giardia duodenalis and identification of potentially zoonotic subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Fayer, R.; Bern, C.; Gilman, R.H.; Trout, J.M.; Schantz, P.M.; Das, P.; Lal, A.A.; Xiao, L. Triosephosphate isomerase gene characterization and potential zoonotic transmission of Giardia duodenalis. Emerg. Infect. Dis. 2003, 9, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Tiangtip, R.; Jongwutiwes, S. Molecular analysis of Cryptosporidium species isolated from HIV-infected patients in Thai-land. Trop. Med. Int. Health 2002, 7, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Scicluna, S.M.; Tawari, B.; Clark, C.G. DNA Barcoding of Blastocystis. Protist 2006, 157, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.G.; Molokin, A.; Santin, M. Next generation amplicon sequencing improves detection of Blastocystis mixed subtype infections. Infect. Genet. Evol. 2019, 73, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Gómez-Muñoz, M.T.; Solano-Aguilar, G.; Fayer, R. Development of a new PCR protocol to detect and subtype Blastocystis spp. from humans and animals. Parasitol. Res. 2011, 109, 205–212. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap Download. 2014. Available online: SourceForge.net (accessed on 6 February 2021).
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Buckholt, M.A.; Lee, J.H.; Tzipori, S. Prevalence of Enterocytozoon bieneusi in swine: An 18-month survey at a slaughter-house in Massachusetts. Appl. Environ. Microbiol. 2002, 68, 2595–2599. [Google Scholar] [CrossRef] [PubMed]
- Verweij, J.J.; Oostvogel, F.; Brienen, E.A.; Nang-Beifubah, A.; Ziem, J.; Polderman, A.M. Prevalence of Entamoeba histolytica and Entamoeba dispar in northern Ghana. Trop. Med. Int. Health 2003, 8, 1153–1156. [Google Scholar] [CrossRef]
- Gutiérrez-Cisneros, M.J.; Cogollos, R.; López-Vélez, R.; Martín-Rabadán, P.; Martínez-Ruiz, R.; Subirats, M.; Merino, F.J.; Fuentes, I. Application of real-time PCR for the differentiation of Entamoeba histolytica and E. dispar in cyst-positive faecal samples from 130 immigrants living in Spain. Ann. Trop. Med. Parasitol. 2010, 104, 145–149. [Google Scholar] [CrossRef]
- Ponce-Gordo, F.; Fonseca-Salamanca, F.; Martínez-Díaz, R.A. Genetic heterogeneity in internal transcribed spacer genes of Balantidium coli (Litostomatea, Ciliophora). Protist 2011, 162, 774–794. [Google Scholar] [CrossRef]
- Vallo, P.; Petrželková, K.J.; Profousová, I.; Petrášová, J.; Pomajbíková, K.; Leendertz, F.; Hashimoto, C.; Simmons, N.; Babweteera, F.; Machanda, Z.; et al. Molecular diversity of entodiniomorphid ciliate Troglodytella abrassarti and its coevolution with chimpanzees. Am. J. Phys. Anthropol. 2012, 148, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Cruz, I.; Chicharro, C.; Nieto, J.; Bailo, B.; Canavate, C.; Figueras, M.A.-C.; Alvar, J. Comparison of new diagnostic tools for management of pediatric Mediterranean visceral leishmaniasis. J. Clin. Microbiol. 2006, 44, 2343–2347. [Google Scholar] [CrossRef][Green Version]
- Ježková, J.; Prediger, J.; Holubová, N.; Sak, B.; Konečný, R.; Feng, Y.; Xiao, L.; Rost, M.; McEvoy, J.; Kváč, M. Cryptosporidium ratti n. sp. (Apicomplexa: Cryptosporidiidae) and genetic diversity of Cryptosporidium spp. in brown rats (Rattus norvegicus) in the Czech Republic. Parasitology 2021, 148, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L. The burden and etiology of diarrheal illness in developing countries. Pediatr. Clin. North Am. 2017, 64, 799–814. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Gracenea, M.; Gómez, M.S.; Arrizabalaga, A.; González-Moreno, O. The occurrence of Cryptosporidium parvum and C. muris in wild rodents and insectivores in Spain. Vet. Parasitol. 2000, 92, 253–260. [Google Scholar] [CrossRef]
- García-Livia, K.; Martín-Alonso, A.; Foronda, P. Diversity of Cryptosporidium spp. in wild rodents from the Canary Islands, Spain. Parasites Vectors 2020, 13, 445. [Google Scholar] [CrossRef] [PubMed]
- Guy, R.A.; Yanta, C.A.; Muchaal, P.K.; Rankin, M.A.; Thivierge, K.; Lau, R.; Boggild, A.K. Molecular characterization of Cryptosporidium isolates from humans in Ontario, Canada. Parasites Vectors 2021, 14, 1–14. [Google Scholar] [CrossRef]
- Ayinmode, A.B.; Obebe, O.O.; Falohun, O.O. Molecular detection of Cryptosporidium species in street-sampled dog faeces in Ibadan, Nigeria. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Ying, J.L.J.; Monis, P.; Ryan, U. Molecular characterisation of Cryptosporidium and Giardia in cats (Felis catus) in Western Australia. Exp. Parasitol. 2015, 155, 13–18. [Google Scholar] [CrossRef]
- Pavlásek, I.; Ryan, U. The first finding of a natural infection of Cryptosporidium muris in a cat. Vet. Parasitol. 2007, 144, 349–352. [Google Scholar] [CrossRef] [PubMed]
- De Lucio, A.; Martínez-Ruiz, R.; Merino, F.J.; Bailo, B.; Aguilera, M.; Fuentes, I.; Carmena, D. Molecular genotyping of Giardia duodenalis isolates from symptomatic individuals attending two major public hospitals in Madrid, Spain. PLoS ONE 2015, 10, e0143981. [Google Scholar] [CrossRef] [PubMed]
- Azcona-Gutiérrez, J.M.; De Lucio, A.; Hernández-De-Mingo, M.; García-García, C.; Soria-Blanco, L.M.; Morales, L.; Aguilera, M.; Fuentes, I.; Carmena, D. Molecular diversity and frequency of the diarrheagenic enteric protozoan Giardia duodenalis and Cryptosporidium spp. in a hospital setting in Northern Spain. PLoS ONE 2017, 12, e0178575. [Google Scholar] [CrossRef] [PubMed]
- Cardona, G.A.; Carabin, H.; Goñi, P.; Arriola, L.; Robinson, G.; Fernández-Crespo, J.C.; Clavel, A.; Chalmers, R.M.; Carmena, D. Identification and molecular characterization of Cryptosporidium and Giardia in children and cattle populations from the province of Álava, North of Spain. Sci. Total Environ. 2011, 412/413, 101–108. [Google Scholar] [CrossRef]
- Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernández-de-Mingo, M.; Reh, L.; Balasegaram, S.; Verlander, N.Q.; Ruiz Chércoles, E.; Carmena, D. Molecular diversity of Giardia duodenalis, Cryptosporidium spp. and Blastocystis sp. in asymptomatic school children in Leganés, Madrid (Spain). Microorganisms 2020, 8, 466. [Google Scholar] [CrossRef]
- Renelies-Hamilton, J.; Noguera-Julian, M.; Parera, M.; Paredes, R.; Pacheco, L.; Dacal, E.; Saugar, J.M.; Rubio, J.M.; Poulsen, M.; Köster, P.C.; et al. Exploring interactions between Blastocystis sp., Strongyloides spp. and the gut microbiomes of wild chimpanzees in Senegal. Infect. Genet. Evol. 2019, 74, 104010. [Google Scholar] [CrossRef]
- Paulos, S.; Köster, P.C.; De Lucio, A.; Hernández-De-Mingo, M.; Cardona, G.A.; Fernández-Crespo, J.C.; Stensvold, C.R.; Carmena, D. Occurrence and subtype distribution of Blastocystis sp. in humans, dogs and cats sharing household in northern Spain and assessment of zoonotic transmission risk. Zoonoses Public Health 2018, 65, 993–1002. [Google Scholar] [CrossRef]
- Köster, P.C.; Molina, A.; García, M.; Cifre, S.; Trelis, M.; Pérez de Ayala, A.; Azcona, J.M.; García, C.; Paulos, S.; Hernández-de-Mingo, M.; et al. Molecular diversity and frequency of Blastocystis sp. subtypes in Spanish clinical patients: A pilot multicentre study. In Proceedings of the 2nd International Blastocystis Conference, Bogotá, Colombia, 9–12 October 2018. [Google Scholar]
- Oliveira-Arbex, A.P.; David, É.B.; Tenório, M.D.S.; Cicchi, P.J.P.; Patti, M.; Coradi, S.T.; Lucheis, S.B.; Jim, J.; Guimarães, S. Diversity of Blastocystis subtypes in wild mammals from a zoo and two conservation units in southeastern Brazil. Infect. Genet. Evol. 2020, 78, 104053. [Google Scholar] [CrossRef]
- Helenbrook, W.D.; Shields, W.M.; Whipps, C.M. Characterization of Blastocystis species infection in humans and mantled howler monkeys, Alouatta palliata aequatorialis, living in close proximity to one another. Parasitol. Res. 2015, 114, 2517–2525. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Alfellani, M.A.; Nørskov-Lauritsen, S.; Prip, K.; Victory, E.L.; Maddox-Hyttel, C.; Nielsen, H.V.; Clark, C.G. Subtype distribution of Blastocystis isolates from synanthropic and zoo animals and identification of a new subtype. Int. J. Parasitol. 2009, 39, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Mohammadpour, I.; Bozorg-Ghalati, F.; Gazzonis, A.L.; Manfredi, M.T.; Motazedian, M.H.; Mohammadpour, N. First molecular subtyping and phylogeny of Blastocystis sp. isolated from domestic and synanthropic animals (dogs, cats and brown rats) in southern Iran. Parasites Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Alfellani, M.; Clark, C.G. Levels of genetic diversity vary dramatically between Blastocystis subtypes. Infect. Genet. Evol. 2012, 12, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhong, Z.; Song, Y.; Gong, C.; Deng, L.; Cao, Y.; Zhou, Z.; Cao, X.; Tian, Y.; Li, H.; et al. Human-pathogenic Enterocytozoon bieneusi in captive giant pandas (Ailuropoda melanoleuca) in China. Sci. Rep. 2018, 8, 6590. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, J.; Li, N.; Guo, Y.; Feng, Y.; Feng, Y.; Xiao, L. Genotypes and public health potential of Enterocytozoon bie-neusi and Giardia duodenalis in crab-eating macaques. Parasites Vectors 2019, 12, 254. [Google Scholar] [CrossRef]
- Galván, A.L.; Sánchez, A.M.; Valentín, M.A.; Henriques-Gil, N.; Izquierdo, F.; Fenoy, S.; del Aguila, C. First cases of mi-crosporidiosis in transplant recipients in Spain and review of the literature. J. Clin. Microbiol. 2011, 49, 1301–1306. [Google Scholar] [CrossRef]
- Galván-Díaz, A.L.; Magnet, A.; Fenoy, S.; Henriques-Gil, N.; Haro, M.; Gordo, F.P.; Millán, J.; Miró, G.; Del Águila, C.; Izquierdo, F. Microsporidia detection and genotyping study of human pathogenic E. bieneusi in animals from Spain. PLoS ONE 2014, 9, e92289. [Google Scholar] [CrossRef]
- Dashti, A.; Santín, M.; Cano, L.; de Lucio, A.; Bailo, B.; de Mingo, M.H.; Köster, P.C.; Fernández-Basterra, J.A.; Aramburu-Aguirre, J.; López-Molina, N.; et al. Occurrence and genetic diversity of Enterocytozoon bieneusi (Microsporidia) in owned and sheltered dogs and cats in Northern Spain. Parasitol. Res. 2019, 118, 2979–2987. [Google Scholar] [CrossRef]
- Galván, A.L.; Magnet, A.; Izquierdo, F.; Fenoy, S.; Rueda, C.; Fernández Vadillo, C.; Henriques-Gil, N.; Del Aguila, C. Molecular characterization of human-pathogenic Microsporidia and Cyclospora cayetanensis isolated from various water sources in Spain: A year-long longitudinal study. Appl. Environ. Microbiol. 2013, 79, 449–459. [Google Scholar] [CrossRef]
- Kooriyama, T.; Hasegawa, H.; Shimozuru, M.; Tsubota, T.; Nishida, T.; Iwaki, T. Parasitology of five primates in Mahale Mountains National Park, Tanzania. Primates 2012, 53, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Petrželková, K.J.; Schovancová, K.; Profousová, I.; Kišidayová, S.; Váradyová, Z.; Pekár, S.; Kamler, J.; Modrý, D. The effect of low- and high-fiber diets on the population of entodiniomorphid ciliates Troglodytella abrassarti in captive chimpanzees (Pan troglodytes). Am. J. Primatol. 2012, 74, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Ortega-García, M.V.; Salguero, F.J.; Rodríguez-Bertos, A.; Moreno, I.; García, N.; García-Seco, T.; Luz Torre, G.; Domínguez, L.; Domínguez, M. A pathological study of Leishmania infantum natural infection in European rabbits (Oryctolagus cuniculus) and Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 66, 2474–2481. [Google Scholar] [CrossRef] [PubMed]
- Millán, J. Molecular investigation of vector-borne parasites in wild micromammals, Barcelona (Spain). Parasitol. Res. 2018, 117, 3015–3018. [Google Scholar] [CrossRef]
- Galán-Puchades, M.T.; Gómez-Samblás, M.; Suárez-Morán, J.M.; Osuna, A.; Sanxis-Furió, J.; Pascual, J.; Bueno-Marí, R.; Franco, S.; Peracho, V.; Montalvo, T.; et al. Leishmaniasis in Norway Rats in Sewers, Barcelona, Spain. Emerg. Infect. Dis. 2019, 25, 1222–1224. [Google Scholar] [CrossRef]



{kind=link}
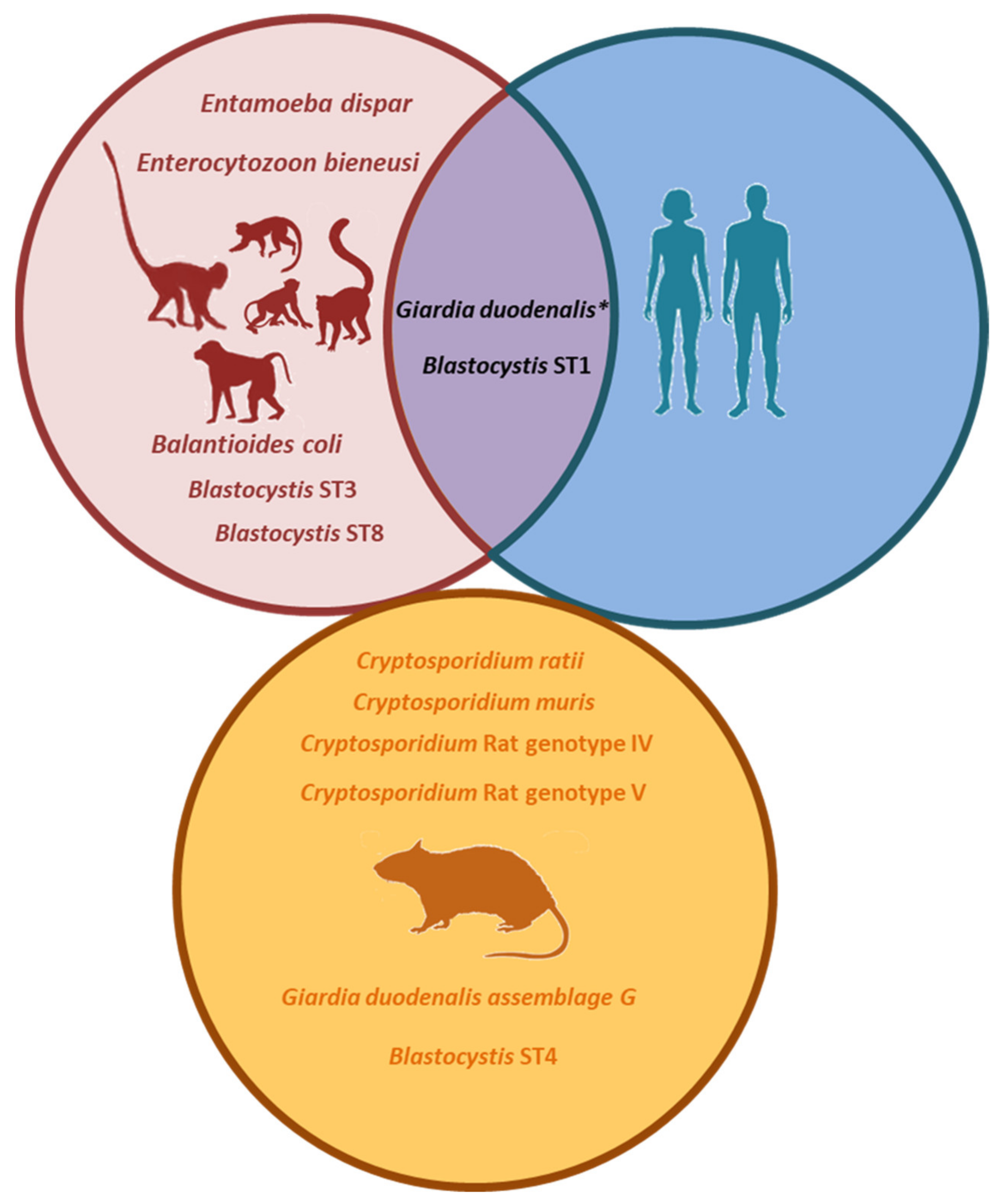{kind=link}
| First Sampling Campaign | Second Sampling Campaign | All | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frequency Positive Results (%) | Frequency Positive Results (%) | Frequency Positive Results (%) | ||||||||||||||||
| Species | No. | Bl | Ed | Gd | Bc | Eb | No. | Bl | Ed | Gd | Bc | Eb | No. | Bl | Ed | Gd | Bc | Eb |
| Cebuella pygmaea | 1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.0 | 0.0 | 50.0 | 0.0 | 0.0 | 3 | 0.0 | 0.0 | 33.3 | 0.0 | 0.0 |
| Cercocebus lunulatus | 3 | 100 | 0.0 | 100 | 0.0 | 0.0 | 3 | 33.3 | 100 | 66.7 | 0.0 | 0.0 | 6 | 66.7 | 50.0 | 83.3 | 0.0 | 0.0 |
| Cercopithecus neglectus | 2 | 50.0 | 0.0 | 50.0 | 0.0 | 0.0 | 3 | 66.7 | 66.7 | 0.0 | 0.0 | 0.0 | 5 | 60.0 | 40.0 | 20.0 | 0.0 | 0.0 |
| Eulemur fulvus | 2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Hylobates leucogenys Ogilby | 4 | 75.0 | 100 | 25.0 | 0.0 | 0.0 | 3 | 33.3 | 33.3 | 0.0 | 0.0 | 33.3 | 7 | 57.1 | 71.4 | 14.3 | 0.0 | 14.3 |
| Lemur catta | 2 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2 | 100 | 0.0 | 0.0 | 0.0 | 0.0 |
| Macaca sylvanus | 5 | 40.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3 | 33.3 | 100 | 100 | 0.0 | 0.0 | 8 | 37.5 | 37.5 | 37.5 | 0.0 | 0.0 |
| Mandrillus leucophaeus | 5 | 40.0 | 0.0 | 0.0 | 20.0 | 0.0 | 3 | 33.3 | 33.3 | 0.0 | 33.3 | 0.0 | 8 | 37.5 | 12.5 | 0.0 | 25.0 | 0.0 |
| Saimiri sciureus | 2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Varecia variegata variegata | 2 | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 5 | 80.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Total | 28 | 50.0 | 14.3 | 17.9 | 3.6 | 0.0 | 23 | 39.1 | 43.5 | 26.1 | 4.3 | 4.3 | 51 | 45.1 | 27.5 | 21.6 | 3.9 | 2.0 |
| Species Combination | No. of Faecal Samples |
|---|---|
| Blastocystis sp. Only | 10 |
| E. dispar only | 4 |
| G. duodenalis only | 1 |
| Blastocystis sp. + E. dispar | 5 |
| Blastocystis sp. + G. duodenalis | 4 |
| G. duodenalis + E. dispar | 3 |
| Blastocystis sp. + B. coli | 1 |
| E. dispar + E. bieneusi | 1 |
| G. duodenalis + Blastocystis sp. + E. dispar | 3 |
| Total | 32 |
| Species | Genotype | Sub-Genotype | Host Species | No. of Isolates | Locus | Reference Sequence | Stretch | Single Nucleotide Polymorphisms | GenBank ID |
|---|---|---|---|---|---|---|---|---|---|
| Giardia duodenalis | A | AII | C.t. | 1 | bg | AY072723 | 205–539 | None | MW417420 |
| Blastocystis sp. | ST1 | Allele 1 | M.l., H.l., C.t. | 4 | ssu rRNA | MK357786 | 4–602 | None | MW414634 |
| Allele 2 | C.t. | 1 | ssu rRNA | MT094302 | 36–539 | None | MW414635 | ||
| Allele 2 | C.n. | 1 | ssu rRNA | MT094302 | 32–539 | C57A, 65InsG, A112G, C128A, C237T, C272T, A458C | MW414636 | ||
| Alleles 1 + 2 | C.t., M.l. | 3 | ssu rRNA | MK357786 | 1–603 | G128R, A474W | MW414637 | ||
| ST3 | Allele 34 | H.l., M.c., C.n. | 7 | ssu rRNA | MK801359 | 1–581 | G114A, A115T, A116G, A159G, T160A, A161T | MW414638 | |
| Alleles 32 + 34 | C.n. | 1 | ssu rRNA | MK801359 | 1–586 | G114A, A115T, A116G, A159K, T160R, A161K, A162R | MW414639 | ||
| ST8 | Allele 21 | V.v.v., L. c. | 6 | ssu rRNA | MT509451 | 1–525 | None | MW414640 | |
| Balantioides coli | Unknown | - | M.l. | 2 | ITS | - | - | - | - |
| Enterocytozoon bieneusi | D | - | H.l. | 1 | ITS | AF101200 | 31–419 | None | MW414645 |
| Variable | Subject 38 | Subject 79 | Subject 86 |
|---|---|---|---|
| Sociodemographic factors | |||
| Sex | Male | Male | Female |
| Age (years) | 56 | 58 | 49 |
| Protist infection/carriage | |||
| Giardia duodenalis | Negative | Positive | Positive |
| Blastocystis sp. | Positive | Negative | Positive |
| Clinical factors | |||
| Diarrhoea in the last 7 days | No | No | No |
| Contact with children <5-years | No | No | No |
| Diarrhoea in family members/relatives | Yes | No | No |
| Work-related factors | |||
| Activity | Veterinarian | Zookeeper | Zookeeper |
| Exposure to faeces from NHP | Yes | Yes | Yes |
| Exposure to faeces from animals other than NHP | Yes | Yes | Yes |
| Any of these animal species with diarrhoea | Yes | Yes | Yes |
| Food handler | Yes | Yes | Yes |
| Behavioural factors | |||
| Recent travel | Yes | No | No |
| Contact with pet dogs | Yes | Yes | Yes |
| Contact with pet cats | Yes | No | Yes |
| Main drinking source—tap | Yes | Yes | Yes |
| Main drinking source—bottled | No | No | No |
| Swimming | No | No | No |
| Handwashing | Frequently | Always | Always |
| Vegetable washing | Always | Always | Always |
| Species/Genotype | No. of Isolates | Reference Sequence | Stretch | Single Nucleotide Polymorphisms | GenBank ID |
|---|---|---|---|---|---|
| C. muris | 3 | AB089284 | 504–1012 | None | MW406908 |
| C. rattia | 6 | MT504541 | 293–751 | None | MW406909 |
| Rat genotype IV | 3 | JN172970 | 377–775 | None | MW406910 |
| 1 | JN172970 | 332–798 | C342T, C410T, C423T, 448DelT, G493A | MW406911 | |
| 1 | JN172970 | 332–752 | C342T, C410T, C423T, 490_491DelTT, G493A | MW406912 | |
| 1 | JN172970 | 328–814 | A428G, 448DelT, G493A | MW406913 | |
| 1 | JN172970 | 328–767 | A445G, 448DelT, G493A, T541A, T542A | MW406914 | |
| 1 | JN172970 | 348–776 | A445G, 448DelT, G493A, T541W, T542W | MW406915 | |
| 5 | JN172970 | 375–814 | 448DelT, G493A | MW406916 | |
| 1 | JN172970 | 454–678 | A459T, A475T, G493A | MW406917 | |
| 1 | JN172970 | 332–788 | A472G, 492InsT, G493A | MW406918 | |
| 3 | JN172970 | 341–814 | 490_491DelTT, G493A | MW406919 | |
| 1 | JN172970 | 481–814 | 490_491DelTT, G493A, G635A | MW406920 | |
| Rat genotype V | 1 | MT504543 | 306–699 | A667G | MW406921 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köster, P.C.; Dashti, A.; Bailo, B.; Muadica, A.S.; Maloney, J.G.; Santín, M.; Chicharro, C.; Migueláñez, S.; Nieto, F.J.; Cano-Terriza, D.; et al. Occurrence and Genetic Diversity of Protist Parasites in Captive Non-Human Primates, Zookeepers, and Free-Living Sympatric Rats in the Córdoba Zoo Conservation Centre, Southern Spain. Animals 2021, 11, 700. https://doi.org/10.3390/ani11030700
Köster PC, Dashti A, Bailo B, Muadica AS, Maloney JG, Santín M, Chicharro C, Migueláñez S, Nieto FJ, Cano-Terriza D, et al. Occurrence and Genetic Diversity of Protist Parasites in Captive Non-Human Primates, Zookeepers, and Free-Living Sympatric Rats in the Córdoba Zoo Conservation Centre, Southern Spain. Animals. 2021; 11(3):700. https://doi.org/10.3390/ani11030700
Chicago/Turabian StyleKöster, Pamela C., Alejandro Dashti, Begoña Bailo, Aly S. Muadica, Jenny G. Maloney, Mónica Santín, Carmen Chicharro, Silvia Migueláñez, Francisco J. Nieto, David Cano-Terriza, and et al. 2021. "Occurrence and Genetic Diversity of Protist Parasites in Captive Non-Human Primates, Zookeepers, and Free-Living Sympatric Rats in the Córdoba Zoo Conservation Centre, Southern Spain" Animals 11, no. 3: 700. https://doi.org/10.3390/ani11030700
APA StyleKöster, P. C., Dashti, A., Bailo, B., Muadica, A. S., Maloney, J. G., Santín, M., Chicharro, C., Migueláñez, S., Nieto, F. J., Cano-Terriza, D., García-Bocanegra, I., Guerra, R., Ponce-Gordo, F., Calero-Bernal, R., González-Barrio, D., & Carmena, D. (2021). Occurrence and Genetic Diversity of Protist Parasites in Captive Non-Human Primates, Zookeepers, and Free-Living Sympatric Rats in the Córdoba Zoo Conservation Centre, Southern Spain. Animals, 11(3), 700. https://doi.org/10.3390/ani11030700