1. Introduction
Intestinal homeostasis in piglets is impacted by environmental factors (including stress and dietary nutrients) that regulate the flora colonisation. Specifically, weaned stress, an inevitable event for infants and piglets, reconstructs the microbial ecological community in the gastrointestinal tract [
1,
2]. Tryptophan (Trp) is a vital nutrient for the development of weaned piglets. Trp and its metabolites play important roles in growth performance, intestinal mucosal barrier function, immune regulation, appetite, and general health [
3]. Intestinal antimicrobial peptides are important components of the intestinal mucosal chemical barrier and play a key role in defense against intestinal pathogenic microbial infection. In recent years, studies have shown that Trp promotes the expression of β-defensin 2 (BD-2) in the intestinal mucosa [
4,
5,
6,
7,
8], thereby limiting the negative effects of pathogenic
E. coli on piglet growth [
9]. Our previous study showed that Trp promotes BD-2 expression via the mammalian target of rapamycin (mTOR) pathway and that its metabolite (kynurenine) binds to the aryl hydrocarbon receptor in the rat intestine [
10]. Dietary Trp supplementation improves the growth of weanling piglets and regulates the composition of the microbiota in the hindgut [
11]. However, it remains unclear how adequate and inadequate dietary Trp impact intestinal microbial ecology. At present, the Trp requirement for 28-day-old weaned piglets recommended by the National Research Council (NRC) is 0.21%. Therefore, a corn- and soybean-based diet containing 0.14% Trp was supplemented with 0.0% (deficient), 0.07% (adequate), 0.14% (excess), or 0.21% Trp (excess) according to the nutritional requirements for weaned piglets in the present study. The purpose of our study was to reveal the mechanism of the effects of insufficient, adequate, and excess Trp on the intestinal microflora and growth performance of weaned piglets.
2. Materials and Methods
2.1. Animals, Experimental Design, and Diets
Forty 28-d Duroc × Landrace × Yorkshire weaned piglets (5.42 ± 1.52 kg) obtained from the Chongqing Academy of Animal Science (Chongqing, China) and randomly divided into four groups with 10 barrows per group and one pig per replicate. They were fed a corn- and soybean-based diet (containing 0.14% Trp) supplemented with 0.0% (deficient), 0.07% (adequate), 0.14% (excess), or 0.21% Trp (excess). The final Trp levels of the four maize-soybean meal diets were 0.14%, 0.21%, 0.28%, and 0.35%. The experiment lasted for 4 weeks. The ingredients and composition of the basal diet were formulated according to the NRC requirements (2012) and are listed in
Table 1. The piglets were kept individually in pens (1.5 m length × 0.5 m width × 0.8 m depth) in a mechanically ventilated and temperature-controlled room (22 ± 1.2 °C). Feed and water were provided on an ad libitum basis. All experimental procedures involving piglets were approved by the License of Experimental Animals (SYXK 2014-0002) of the Animal Experimentation Ethics Committee of Southwest University, Chongqing, China.
2.2. Measurements and Sampling
The piglets were weighed on day 28 before the morning feeding. Feed intake was recorded daily throughout the experimental period. Before the morning feeding on day 28, a 10-mL blood sample was collected from the orbital sinus of five piglets selected from each treatment group. The blood samples were left undisturbed for 60 min and then centrifuged at 3500 g for 10 min at 4 °C to harvest the serum. The serum was stored at –20 °C for biochemical analysis and ELISA. After blood sampling, the five piglets selected from each treatment were anaesthetized with an intravenous injection of sodium pentobarbital (50 mg/kg BW) and exsanguinated. The jejunal and ileal mucosa were rinsed with cold saline, then scraped gently with a scalpel blade and collected. The harvested jejunal and ileal mucosa were immediately frozen in liquid N2 and stored at −80 °C for quantitative PCR analysis. Samples of caecum contents collected for analysis of 16S rDNA genes sequencing.
2.3. Growth Performance and Diarrhoea Incidence
Feed intake in pigs was recorded daily. The pigs were weighed on day 28 before the morning feeding. The average daily weight gain (ADG), average daily feed intake (ADFI), and feed conversion rate (FCR) were calculated according to the following formulas:
The diarrhoea incidence, stool shape, and diarrhoea index in pigs were recorded daily according to the scoring criteria reported by Kelly (1990): 0 points for normal faces, 1 point for soft faeces, 2 points for unshaped loose faeces, and 3 points for watery faeces, mucous stools, or purulent bloody stools. The diarrhoea rate and index were calculated according to the following formulas:
2.4. Biochemical and ELISA Analysis
The jejunum mucosa sample (1 g) was homogenized in 9 mL 0.9% NaCl with a polytron (Brinkmann Instruments Inc., Westbury, NY, USA) and centrifuged at 6500 g for 20 min at 4 °C. The supernatant was collected and stored at –80 °C for later analysis. The concentration of growth hormone (GH), cortisol, insulin, melatonin, TPH (Trp hydroxylase), 5-HT (5-hydroxytryptamine), indoleamine 2,3-dioxygenase (IDO), interleukin (IL)-17 and IL-22 in the jejunum and ileum mucosa were determined by porcine-specific ELISA kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
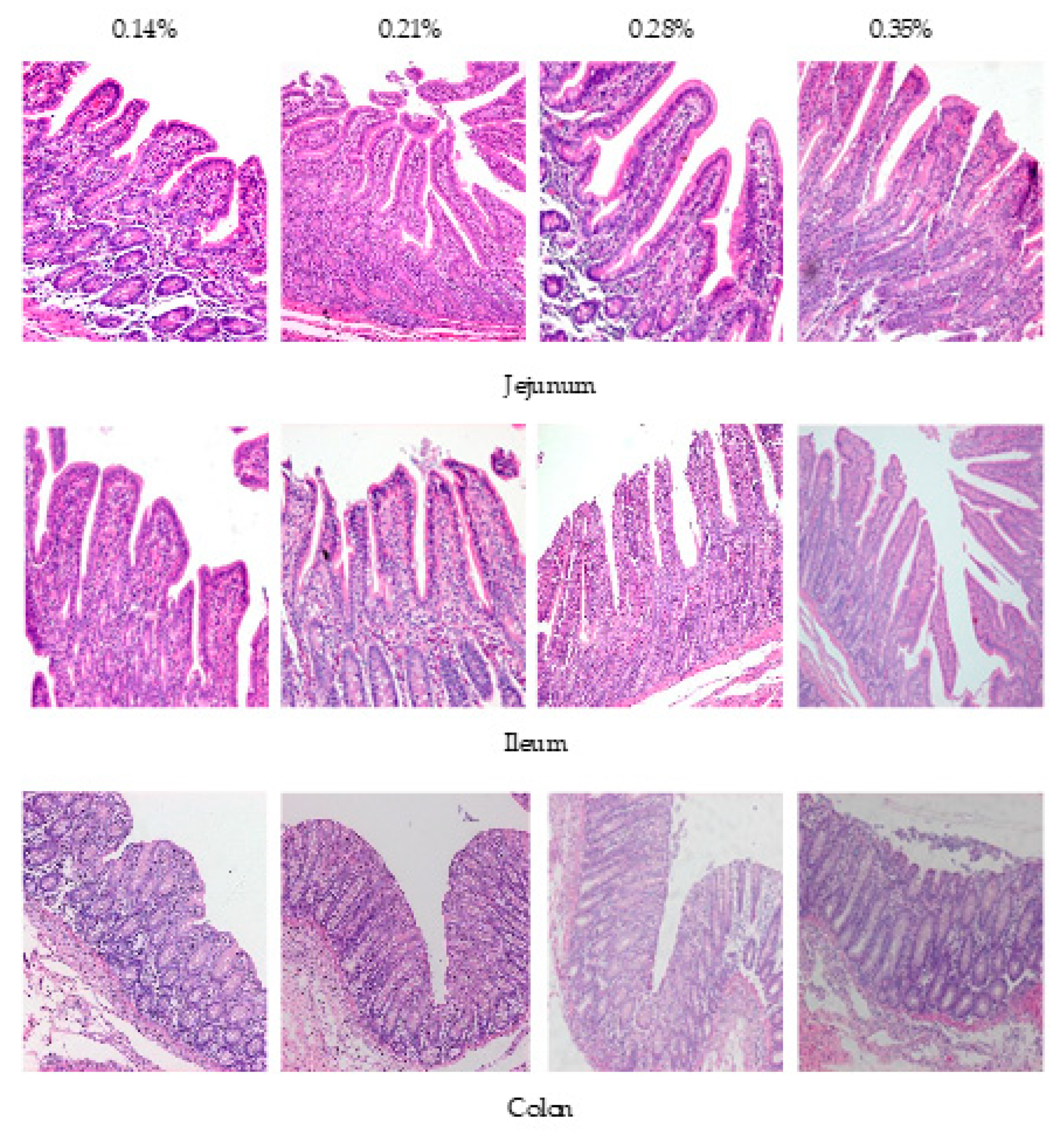
2.5. H&E Staining
The morphology of the jejunum and the ileum was analyzed according to the H&E staining method described by Wang et al. [
12]. The sliced sample was viewed under an optical microscope (Carl Zeiss Inc., Oberkochen, Bayern, Germany). Digital images were taken using a color video camera (Sony 3CCD-VX3 camcorder) for goblet cell measurements and lymphocyte counts. The villus height and crypt depth were measured using image analysis software (Intronic GmbH & Co., Rothenstein, Berlin, Germany).
2.6. Determination of Serum Trp and Kynurenine Concentration
To obtain the supernatant, the collected blood samples were diluted twice with 4% sulfosalicylic acid and centrifuged at 16,000 rpm for 2 min at 4 °C. After the supernatants were filtered with a 0.22 nm filter, the supernatant samples were measured on a Hitachi L-8800 automatic amino acid analyzer with a C18 column (250 mm × 4.6 mm, i.d. 5 μm) and a mobile phase including 0.2 M zinc acetate, 8.3 mM acetic acid and 2.8% acetonitrile at a 1.5 mL/min isocratic flow rate. The Trp excitation wavelength was 254 nm and the emission wavelength was 404 nm. The kynurenine excitation wavelength was 365 nm and the kynurenine emission wavelength was 480 nm. The column temperature was 25 °C and the analysis period for each sample was 60 min. A standard curve was created according to the corresponding peak areas of standard samples (Trp 0, 0.625, 1.25, 2.5, 5, 10, and 20 μM; kynurenine 0, 0.05, 0.5, 2, 10, 20, and 50 μM). The contents of the corresponding amino acids in the samples were calculated using this standard curve.
2.7. Real-Time RT-PCR
The frozen jejunal and ileal mucosal samples (0.05 g) were homogenized in 5 mL of TRIzol reagent containing RNAlater, and total RNA was isolated according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). Total RNA was reverse-transcribed to cDNA by an AMV First Strand cDNA Synthesis Kit (Bio Basic Inc., Markham, ON, Canada). Real-time PCR was performed to quantify the relative amounts of glyceraldehyde-3-phosphate (GAPDH) and BD-2 in the jejunal and ileal mucosa. The primers used to amplify genes in the jejunal mucosa are shown in
Table 2. PCR analysis was performed using the SYBR Green method on the Applied Biosystems 7900HT Fast Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). The thermal cycling parameters were set as follows: 94 °C for 30 s, followed by 40 cycles of 94 °C for 5 s, annealing (GAPDH, 58 °C; BD-2, 57 °C) for 20 s, followed by 72 °C for 20 s. The PCR products were identified by melting curve analysis and sequencing (Sangon Biotech, Shanghai, China). Standard calibration curves for the target genes were created according to the cDNA concentration and Ct value. The expression levels of the target genes were estimated by the calibration curve and normalized against GAPDH expression using REST 2009 Software (Qiagen, Düsseldorf, Germany).
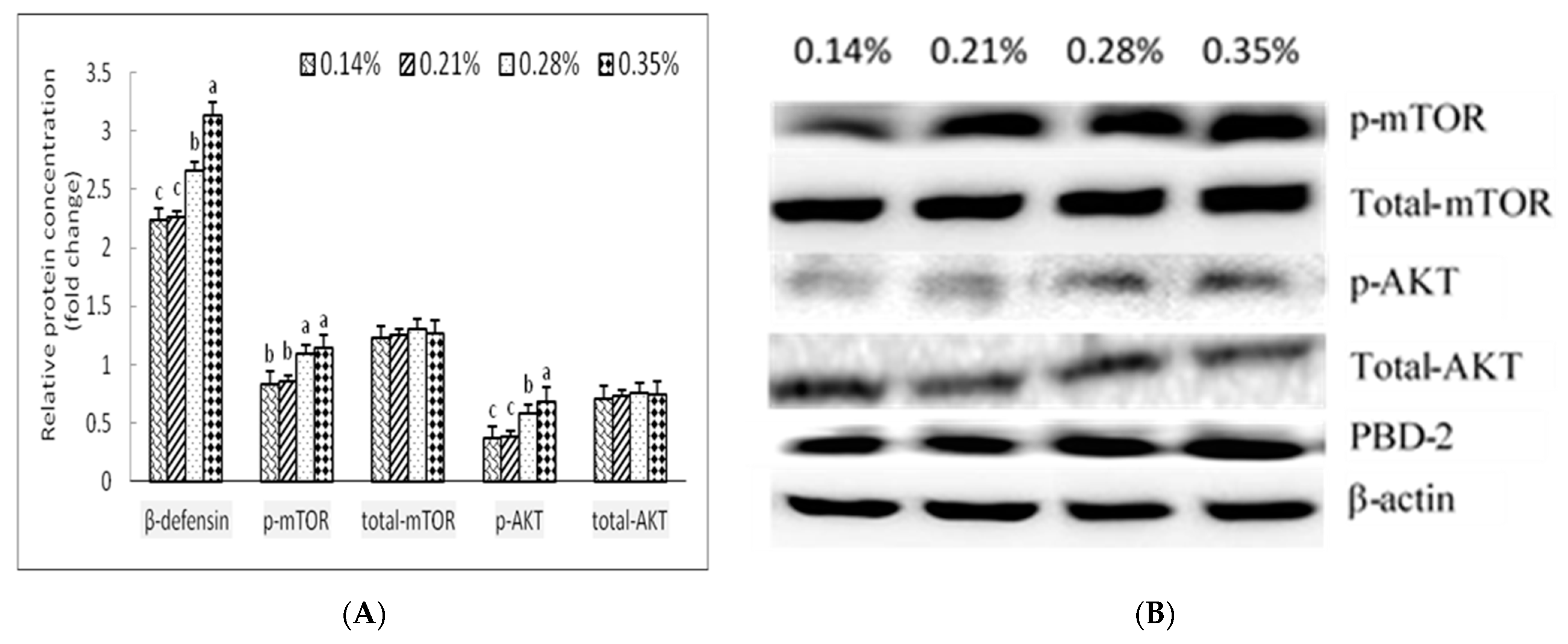
2.8. Western Blot
Samples of jejunal and ileal mucosa (100 mg) were homogenised in 1 mL of RIPA buffer (1.0 mM ethylenediamine tetraacetic acid (EDTA)), 50 mM Tris-base, 0.1% sodium dodecyl sulfate (SDS), 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, and 1 mM phenylmethanesulfonyl fluoride (PMSF) and separated by SDS-polyacrylamide gel electrophoresis. The proteins were transferred to a polyvinylidene fluoride (PVDF) membrane by the semi-dry transfer method. The PVDF membranes were blocked in a blocking buffer overnight at 4 °C, then incubated in blocking buffer with rabbits antibodies for β-actin (13E5) (5125S, 1:1000, Cell Signaling Technology, Boston, MA, USA), BD-2 (ab178728, 1:1000, Abcam, Cambridge, MA, USA), The mechanistic target of rapamycin complex 1 (mTORC1) (ab32028; 1:1000, Abcam), Ser2448-phosphorylated mTOR (ab109268, 1:1000, Abcam), protein kinase B (AKT) (9272, 1:1000, Cell Signaling Technology, Boston, MA, USA), Ser473-phosphorylated AKT (FAB127472; 1:1000; Fantibody). They were then incubated in blocking buffer with F(ab)2 of goat anti-rabbit Ig (1:2500) labelled with horseradish peroxidase diluted in PBSS. The PVDF membrane was soaked in a chemiluminescent liquid (Millipore, Boston, MA, USA). Pictures were taken using a Chemiluminescence Imaging System (Bio-Rad, Bio rad Hercules, CA, USA).
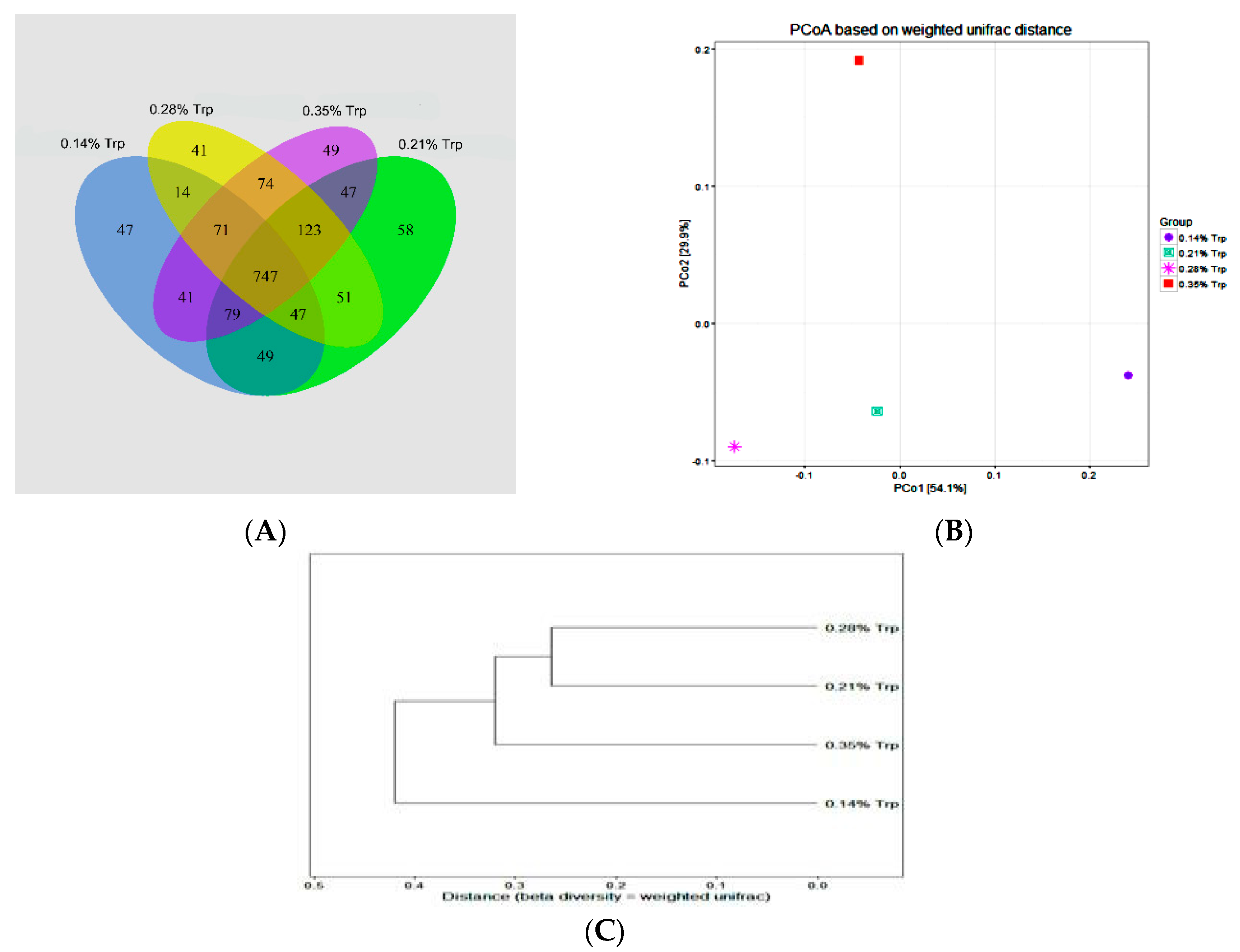
2.9. Analysis of 16S rDNA Genes Sequencing
The DNA samples were extracted from Samples of caecum contents using the Power Fecal DNA Isolation Kit (Mobio, Carlsbad, CA, USA), stained using the Quant-iT Pico Green dsDNA Kit (Invitrogen Ltd., Paisley, UK) and quantified using a Nanodrop spectrophotometer (Nyxor Biotech, Paris, France).
For DNA MiSeq sequencing, PCR amplification of the V4 region of bacterial 16S rDNA was performed using the universal primers 515F (5ʹ-GTGCCAGCM GCCGCGGTAA-3ʹ) and 806R (5ʹ-GGACTACHVGGGTWTCTAAT-3ʹ), incorporating the FLX Titanium adapters and a sample barcode sequence. The cycling parameters were as follows: 5 min of initial denaturation at 95 °C; 25 cycles of denaturation at 95 °C (30 s), annealing at 55 °C (30 s), and elongation at 72 °C (30 s); and final extension at 72 °C for 5 min. Three separate PCR reactions for each sample were pooled for MiSeq sequencing. The PCR products were separated by 1.5% agarose gel electrophoresis and purified using the QIAquick Gel extraction kit (QIAGEN, Düsseldorf, Germany). Amplicons were quantified using the Quant-iT Pico Green dsDNA Assay Kit (Invitrogen). Equal concentrations of amplicons were pooled from each sample. Libraries were constructed using the TruSeq DNA PCR-Free Sample Prep Kit and MiSeq sequencing was performed with the MiSeq Reagent Kit v2 (Illumina, San Diego, CA, USA).
Raw reads were obtained from MiSeq sequencing for analysis. All reads were aligned according to barcode and primer sequences. The resulting sequences were further screened and filtered for quality and length. Sequences that were less than 200 nucleotides or contained ambiguous characters, over two primer mismatches, or mononucleotide repeats of over six nucleotides were removed. The high-quality sequences were assigned to samples according to the barcodes. Sequences with a similarity level of at least 97% were clustered into OTUs (operational taxonomic units) using UPARSE algorithm 7. The representative sequences at an OTU distance of 0.03 were obtained and classified using Ribosomal Database Project (RDP) classifier. Any sequences with annotations for chloroplast, mitochondrial, or archaeal OTUs not identified as being of bacterial origin were excluded from all further analyses. We calculated the abundance-based coverage estimator (ACE), bias-corrected Chao1 richness estimator, and Shannon and Simpson diversity indices. All analyses were performed using mothur (v1.24) (
http://www.mothur.org, accessed on 19 May 2020). Based on the above results for community composition (compositional) and development level and system level (phylogenetic), the analysis of species-level differences, correlation, and alpha, beta, and community structure diversity was performed using QIIME [
13], mothur [
14], and R 4.0.3 software (R Foundation for Statistical Computing, Vienna, Austria).
2.10. Statistical Analysis
All data are presented as means ± Standard Error of Mean (SEM). The data were subjected to one-way analysis of variance using the general linear model procedure in the SAS 8.2statistical software (SAS Institute, Inc., Cary, NC, USA), according to a completely randomised one-factorial design. The LSD test was performed to identify differences between groups. Significance was set at p < 0.05.
5. Conclusions
In conclusion, the results of the present study indicate that adequate provision of Trp decreases the diarrhoea rate and enhances growth performance, intestinal mucosal growth, and microbial ecology in the hindgut as shown by the enhanced abundances of ADG and FCR, as well as the up-regulation of intestinal antimicrobial peptides (porcine BD-2). These beneficial effects of Trp are associated with Trp metabolites, the activation of mTOR signalling, and the enrichment of probiotics in the small intestine of weaned pigs. A corn and soybean meal-based diet with 0.35% Trp may be a nutritional strategy to improve growth performance, intestinal mucosal barrier integrity, and intestinal microbial ecology in weaned piglets.






{kind=link}
{kind=link}
{kind=link}