Tick Infestation and Piroplasm Infection in Barbarine and Queue Fine de l’Ouest Autochthonous Sheep Breeds in Tunisia, North Africa



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
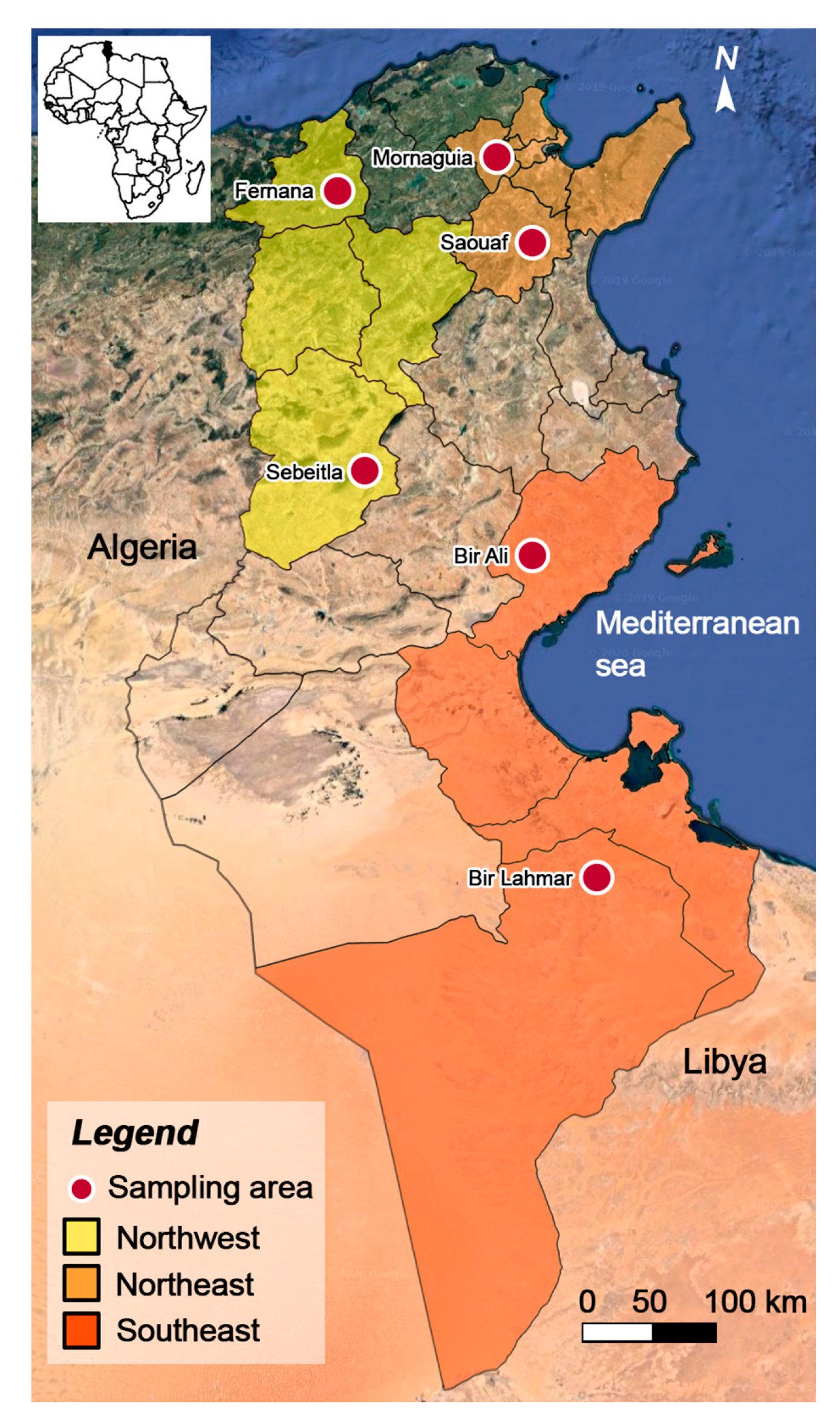
2.1. Study Region and Sampling Design
2.2. Animals
2.3. Sampling and Data Collection
2.4. Hematology
2.5. Molecular Analysis for Piroplasms Detection
2.5.1. DNA Extraction
2.5.2. Polymerase Chain Reactions
2.6. Statistical Analyses
2.7. Ethics Statement
3. Results
3.1. Overall, Collected Tick Population and Overall Parasitological Indicators
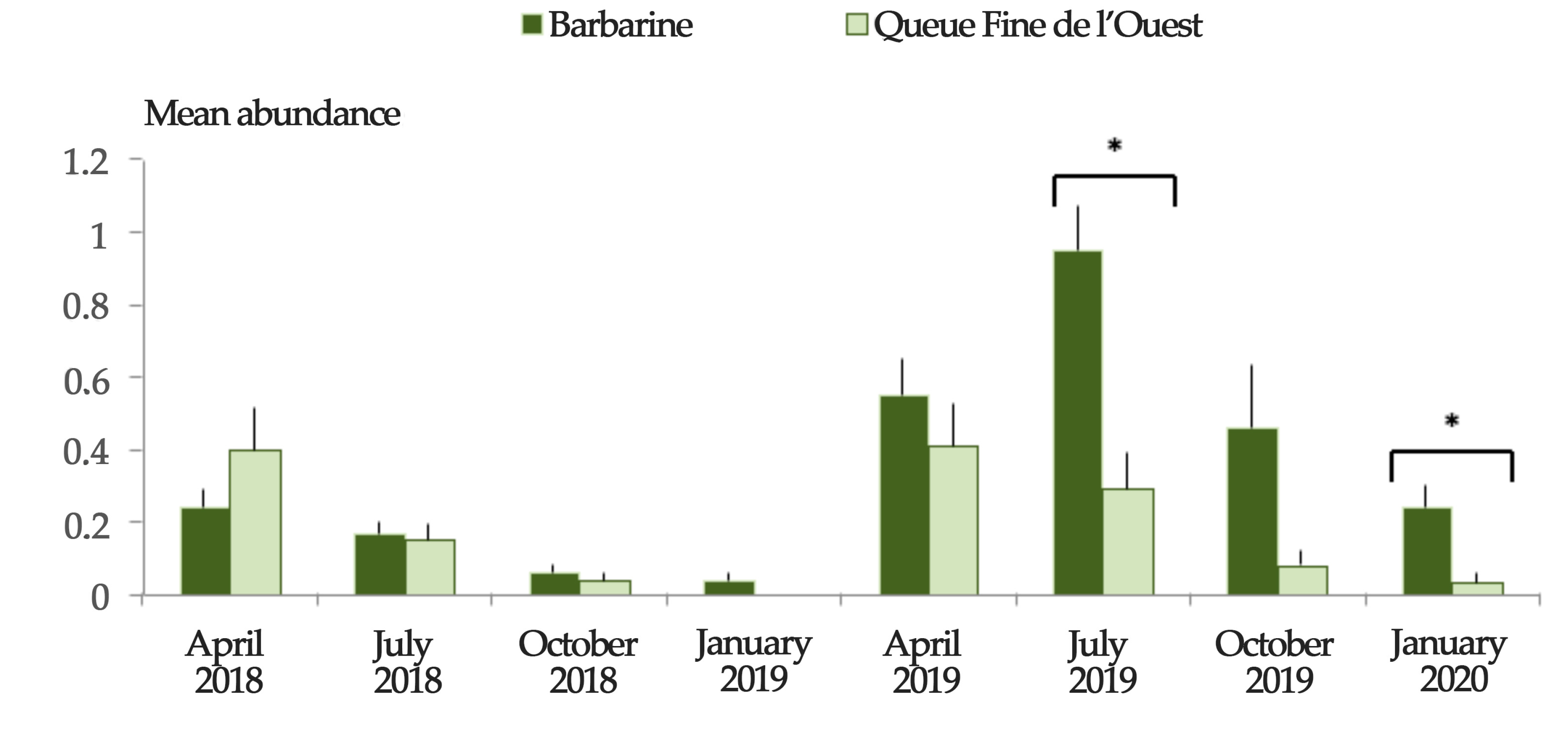
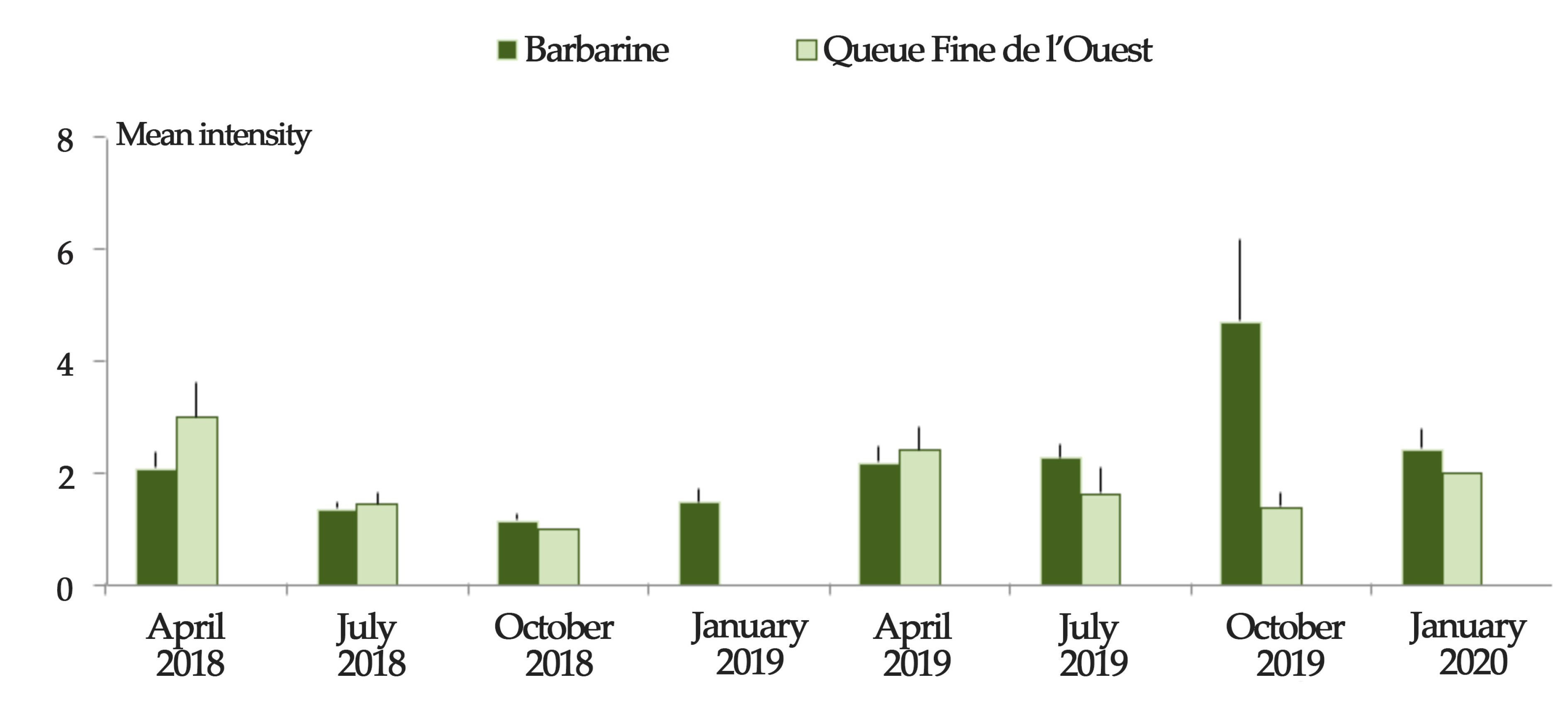
3.2. Breed Differences in Tick Infestation
3.2.1. Tick Species Distribution
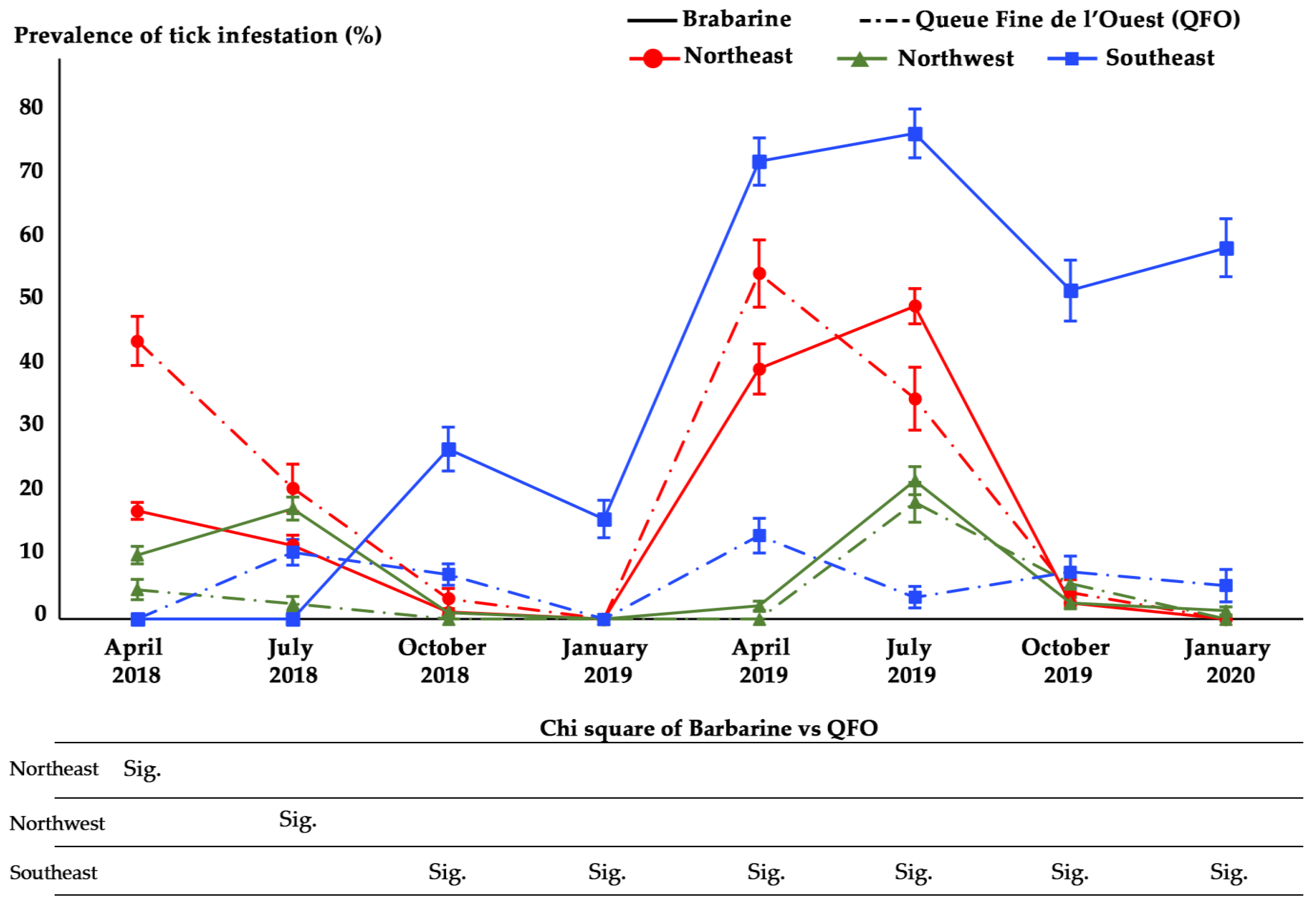
3.2.2. Tick’s Infestation Prevalence According to Sheep Breed and Region
3.3. Molecular Prevalence of Piroplasms
4. Discussion
4.1. Breed Differences
4.2. Sheep Ticks and Agroecological Variability
4.3. Dynamic of Sheep Ticks under Climate Change Context
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Rjeibi, M.R.; Darghouth, M.A.; Gharbi, M. Prevalence of Theileria and Babesia species in Tunisian sheep. Onderstepoort J. Vet. Res. 2016, 83, 6. [Google Scholar] [CrossRef] [PubMed]
- Rjeibi, M.R.; Darghouth, M.A.; Omri, H.; Souidi, K.; Rekik, M.; Gharbi, M. First molecular isolation of Mycoplasma ovis from small ruminants in North Africa. Onderstepoort J. Vet. Res. 2015, 82, e6. [Google Scholar] [CrossRef] [PubMed]
- Belkahia, H.; Ben Said, M.; El Mabrouk, N.; Saidani, M.; Cherni, C.; Ben Hassen, M.; Bouattour, A.; Messadi, L. Seasonal dynamics, spatial distribution and genetic analysis of Anaplasma species infecting small ruminants from Northern Tunisia. Infect. Genet. Evol. 2017, 54, 66–73. [Google Scholar] [CrossRef]
- Ben Said, M.; Belkahia, H.; Alberti, A.; Abdi, K.; Zhioua, M.; Daaloul-Jedidi, M.; Messadi, L. First molecular evidence of Borrelia burgdorferi sensu lato in goats, sheep, cattle and camels in Tunisia. Ann. Agric. Environ. Med. 2016, 23, 442–447. [Google Scholar] [CrossRef]
- De La Fuente, J. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef]
- Kunz, S.; Kemp, D. Insecticides and acaricides: Resistance and environmental impact. Rev. Sci. Tech. 1994, 13, 1249–1286. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: The state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef]
- Shyma, K.P.; Gupta, J.P.; Singh, V. Breeding strategies for tick resistance in tropical cattle: A sustainable approach for tick control. J. Parasit. Dis. 2013, 39, 1–6. [Google Scholar] [CrossRef]
- Råberg, L.; Sim, D.; Read, A.F. Disentangling Genetic Variation for Resistance and Tolerance to Infectious Diseases in Animals. Science 2007, 318, 812–814. [Google Scholar] [CrossRef]
- Marufu, M.C.; Qokweni, L.; Chimonyo, M.; Dzama, K. Relationships between tick counts and coat characteristics in Nguni and Bonsmara cattle reared on semiarid rangelands in South Africa. Ticks Tick Borne Dis. 2011, 2, 172–177. [Google Scholar] [CrossRef]
- De Castro, J.J.; Capstick, P.B.; Nokoe, S.; Kiara, H.; Rinkanya, F.; Slade, R.; Okello, O.; Bennun, L. Towards the selection of cattle for tick resistance in Africa. Exp. Appl. Acarol. 1991, 12, 219–227. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-borne encephalitis virus—A review of an emerging zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Rechav, Y.; Magano, S.R.; Fielden, L.J. The effects of tick numbers and intervals between infestations on the resistance acquired by guinea-pigs to adults of Rhipicephalus evertsi evertsi (Acari: Ixodidae). Exp. Appl. Acarol. 1994, 18, 735–745. [Google Scholar] [CrossRef]
- Rechav, Y.; Fielden, L.J. The effect of various host species on the feeding performance of immature stages of the tick Hyalomma truncatum (Acari: Ixodidae). Exp. Appl. Acarol. 1997, 21, 551–559. [Google Scholar] [CrossRef]
- Robbertse, L.; Richards, S.A.; Maritz-Olivier, C. Bovine Immune Factors Underlying Tick Resistance: Integration and Future Directions. Front. Cell. Infect. Microbiol. 2017, 7, 522. [Google Scholar] [CrossRef]
- Mapholi, N.; Maiwashe, A.; Matika, O.; Riggio, V.; Banga, C.; MacNeil, M.; Muchenje, V.; Nephawe, K.; Dzama, K. Genetic parameters for tick counts across months for different tick species and anatomical locations in South African Nguni cattle. Trop. Anim. Health Prod. 2017, 49, 1201–1210. [Google Scholar] [CrossRef]
- Scholtz, A.J.; Cloete, S.W.P.; Du Toit, E.; Van Wyk, J.B.; De K van der Linde, T.C. A survey of the prevalence of blowfly strike and the control measures used in the Rûens area of the Western Cape Province of South Africa. J. S. Afr. Vet. Assoc. 2011, 82, 107–115. [Google Scholar] [CrossRef][Green Version]
- Bishop, S.C. Possibilities to breed for resistance to nematode parasite infections in small ruminants in tropical production systems. Animal 2012, 6, 741–747. [Google Scholar] [CrossRef]
- Grova, L.; Sae-Lim, P.; Olesen, I. Genetic parameters of tick-infestation on lambs of the Norwegian White Sheep. In Proceedings of the 10th World Congress on Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–24 August 2014. [Google Scholar]
- Budeli, M.; Nephawe, K.; Norris, D.; Selapa, N.; Bergh, L.; Maiwashe, A. Genetic parameter estimates for tick resistance in Bonsmara cattle. S. Afr. J. Anim. Sci. 2010, 39, 321–327. [Google Scholar] [CrossRef][Green Version]
- Maharana, B.; Baithalu, R.; Allaie, I.; Samal, C. Mechanism of Immunity to Tick infestation in Livestock. Vet. World 2011, 4, 131–135. [Google Scholar] [CrossRef]
- Wada, T.; Ishiwata, K.; Koseki, H.; Ishikura, T.; Ugajin, T.; Ohnuma, N.; Obata, K.; Ishikawa, R.; Yoshikawa, S.; Mukai, K.; et al. Selective ablation of basophils in mice reveals their non redundant role in acquired immunity against ticks. J. Clin. Investig. 2010, 120, 2867–2875. [Google Scholar] [CrossRef]
- Barriga, O.O.; Andujar, F.; Andrzejewski, W.J. Manifestations of Immunity in Sheep Repeatedly Infested with Amblyomma americanum Ticks. J. Parasitol. 1991, 77, 703. [Google Scholar] [CrossRef]
- Abdul-Amir, I.; Gray, J. Resistance of sheep to laboratory infestations of the tick, Ixodes ricinus. Res. Vet. Sci. 1987, 43, 266–267. [Google Scholar] [CrossRef]
- Boppana, D.K.V.; Dhinakarraj, G.; John, L.; Wikel, S.K.; Latha, B.R.; Gomathinayagam, S. In vivo immunomodulatory effects of ixodid ticks on ovine circulating T- and B-lymphocytes. Parasite Immunol. 2004, 26, 83–93. [Google Scholar] [CrossRef]
- Piper, E.K.; Jackson, L.; Bielefeldt-Ohmann, H.; Gondro, C.; Lew-Tabor, A.E.; Jonsson, N.N. Tick-susceptible Bos taurus cattle display an increased cellular response at the site of larval Rhipicephalus (Boophilus) microplus attachment, compared with tick-resistant Bos indicus cattle. Int. J. Parasitol. 2010, 40, 431–441. [Google Scholar] [CrossRef]
- Marufu, M.C.; Chimonyo, M.; Mans, B.J.; Dzama, K. Cutaneous hypersensitivity responses to Rhipicephalus tick larval antigens in pre-sensitized cattle. Ticks Tick Borne Dis. 2013, 4, 311–316. [Google Scholar] [CrossRef]
- Elati, K.; Hamdi, D.; Jdidi, M.; Rekik, M.; Gharbi, M. Differences in tick infestation of Tunisian sheep breeds. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 50–54. [Google Scholar] [CrossRef]
- Rekik, M.; Aloulou, R.; Ben Hammouda, M. Small Ruminant Breeds of Tunisia. In Characterisation of Small Ruminant Breeds in West Asia and North Africa; Iniguez, L., Ed.; International Center for Agricultural Research in the Dry Areas (ICARDA): Beirut, Lebanon, 2005; pp. 91–140. [Google Scholar]
- Bedhiaf-Romdhani, S.; Djemali, M.; Zaklouta, M.; Iñiguez, L. Monitoring crossbreeding trends in native Tunisian sheep breeds. Small Rumin. Res. 2008, 74, 274–278. [Google Scholar] [CrossRef]
- Rouabeh, A.; Jemli, M.H. Agricultura in Tunisia; Fondazione de Sardegna: Sardaigne, Italy, 2016; Available online: https://slideplayer.fr/slide/12024178/ (accessed on 5 July 2019).
- Ben Sassi-Zaidy, Y.; Maretto, F.; Charfi-Cheikrouha, F.; Cassandro, M. Genetic diversity, structure, and breed relationships in Tunisian sheep. Small Rumin. Res. 2014, 119, 52–56. [Google Scholar] [CrossRef]
- Mason, I.L. Sheep Breeds of the Mediterranean; Bucks, F.R., Ed.; Commonwealth Agricultural Bureaux: Edinburgh, UK, 1967. [Google Scholar]
- Djemali, M.; Aloulou, R.; Sassi, M. Adjustment factors and genetic and phenotypic parameters for growth traits of Barbarine lambs in Tunisia. Small Rumin. Res. 1994, 13, 41–47. [Google Scholar] [CrossRef]
- Tolone, M.; Mastrangelo, S.; Rosa, A.J.; Portolano, B. Genetic diversity and population structure of Sicilian sheep breeds using microsatellite markers. Small Rumin. Res. 2012, 102, 18–25. [Google Scholar] [CrossRef]
- Ben Salem, H.; Lassoued, N.; Rekik, M. Merits of the fat-tailed Barbarine sheep raised in different production systems in Tunisia: Digestive, productive and reproductive characteristics. Trop. Anim. Health Prod. 2011, 43, 1357–1370. [Google Scholar] [CrossRef] [PubMed]
- Khaldi, Z.; Rekik, B.; Haddad, B.; Zourgui, L.; Souid, S. Genetic characterization of three ovine breeds in Tunisia using randomly amplified polymorphic DNA markers. Livest. Res. Rural Dev. 2010, 22, 3. [Google Scholar]
- Atti, N.; Khaldi, G. Performance of the fat-tailed Barbarine sheep in its environment: Adaptive capacity to alternation of underfeeding and re-feeding periods. A review. Anim. Res. 2004, 53, 165–176. [Google Scholar] [CrossRef]
- Bedhiaf-Romdhani, S.; Djemali, M.; Belló, A. Inventaire des différents écotypes de la race Barbarine en Tunisie. Anim. Genet. Resour. Inf. 2008, 43, 41–47. [Google Scholar] [CrossRef]
- Sassi-Zaidy, Y.B.; Maretto, F.; Zanetti, E.; Hajji, G.M.; Charfi-Cheikrouha, F.; Cassandro, M. Genetic structure and variability within and among populations of the fat-tailed Barbarine sheep breed using microsatellites markers. Afr. J. Biotechnol. 2014, 13, 44–54. [Google Scholar] [CrossRef][Green Version]
- M’naour, D. Genetic Improvement Objectives of Sheep and Goats in Tunisia. Lessons Learned. In Analysis and Definition of the Objectives in Genetic Improvement Programmes in Sheep and Goats. An Economic Approach to Increase Their Profitability; CIHEAM: Zaragoza, Spain, 2000; Volume 43, pp. 121–127. [Google Scholar]
- El Hentati, H. Genetic diversity of two Tunisian sheep breeds using random amplified polymorphic DNA (RAPD) analysis. Afr. J. Biotechnol. 2012, 11, 4109–4115. [Google Scholar] [CrossRef][Green Version]
- Laval, G.; Iannuccelli, N.; Legault, C.; Milan, D.; Groenen, M.A.; Giuffra, E.; Andersson, L.; Nissen, P.H.; Jørgensen, C.B.; Beeckmann, P.; et al. Genetic diversity of eleven European pig breeds. Genet. Sel. Evol. 2000, 32, 187–203. [Google Scholar] [CrossRef]
- Kenyon, P.R.; Maloney, S.K.; Blache, D. Review of sheep body condition score in relation to production characteristics. N. Zeal. J. Agric. Res. 2014, 57, 38–64. [Google Scholar] [CrossRef]
- Walker, A.R.; Bouattour, A.; Camicas, J.; Estrada-Pena, A.; Horac, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003; ISBN 0-9545173-0-X. [Google Scholar]
- Cebra, C.; Cebra, M. Diseases of the Hematologic, Immunologic, and Lymphatic Systems (Multisystem Diseases). In Sheep and Goat Medicine; Pugh, D.G., Braid, A.N., Eds.; Elsevier-Saunders: Philadelphia, PA, USA, 2012; pp. 466–514. ISBN 9781437723533. [Google Scholar]
- Wang, Y.; Tian, R.-M.; Gao, Z.M.; Bougouffa, S.; Qian, P.-Y. Optimal Eukaryotic 18S and Universal 16S/18S Ribosomal RNA Primers and Their Application in a Study of Symbiosis. PLoS ONE 2014, 9, e90053. [Google Scholar] [CrossRef]
- Gubbels, J.M.; De Vos, A.P.; Van Der Weide, M.; Viseras, J.; Schouls, L.M.; De Vries, E.; Jongejan, F. Simultaneous Detection of Bovine Theileria and Babesia Species by Reverse Line Blot Hybridization. J. Clin. Microbiol. 1999, 37, 1782–1789. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D. Méthodes Statistiques à L’usage des Médecins et des Biologistes, 4th ed.; Flammarion: Paris, France, 1993; ISBN 2257103262. [Google Scholar]
- Sae-Lim, P.; Grøva, L.; Olesen, I.; Varona, L. A comparison of nonlinear mixed models and response to selection of tick-infestation on lambs. PLoS ONE 2017, 12, e0172711. [Google Scholar] [CrossRef]
- Zaibet, L.; Abelhafid, H.; Hammami, S.; Jabbar, M. Efficience du Marché des Petits Ruminants: Rôle de la Santé Animale et Coûts de Transaction. In Sustainability of Small Ruminant Production Systems in Tunisia: A Health Marketing Approach; Zaibet, L., Hammami, S., Jabbar, M., Eds.; International Livestock Research Institute: Nairobi, Kenya, 2009; pp. 41–58. [Google Scholar]
- Rouatbi, M.; Amairia, S.; Amdouni, Y.; Boussaadoun, M.A.; Ayadi, O.; Al-Hosary, A.A.T.; Rekik, M.; Ben Abdallah, R.; Aoun, K.; Darghouth, M.A.; et al. Toxoplasma gondii infection and toxoplasmosis in North Africa: A review. Parasite 2019, 26, 6. [Google Scholar] [CrossRef]
- Amdouni, Y.; Rouatbi, M.; Lassoued, N.; Rekik, M.; Gharbi, M. Neospora caninum Natural Infection in Tunisian Rams: Serological Study and Molecular Identification of Infection in Semen. Acta Parasitol. 2019, 64, 821–828. [Google Scholar] [CrossRef]
- Rouatbi, M.; Gharbi, M.; Rjeibi, M.R.; Ben Salem, I.; Akkari, H.; Lassoued, N.; Rekik, M. Effect of the infection with the nematode Haemonchus contortus (Strongylida: Trichostrongylidae) on the haematological, biochemical, clinical and reproductive traits in rams. Onderstepoort J. Vet. Res. 2016, 83. [Google Scholar] [CrossRef] [PubMed]
- Elati, K.; Aloui, S.; Dhibi, M.; Rekik, M.; Gharbi, M. Seasonal variation of lungworm infestation of ewes at Sidi Bouzid slaughterhouse (Central Tunisia). Rev. Élev. Méd. Vét. Pays Trop. 2018, 70, 137–141. [Google Scholar] [CrossRef][Green Version]
- Rechav, Y. Naturally acquired resistance to ticks—A global view. Int. J. Trop. Insect Sci. 1992, 13, 495–504. [Google Scholar] [CrossRef]
- Rechav, Y.; Kostrzewski, M.W.; Els, D.A. Resistance of indigenous African cattle to the tick Amblyomma hebraeum. Exp. Appl. Acarol. 1991, 12, 229–241. [Google Scholar] [CrossRef]
- Rechav, Y.; Clarke, F.C.; Dauth, J. Acquisition of immunity in cattle against the blue tick, Boophilus decoloratus. Exp. Appl. Acarol. 1991, 11, 51–56. [Google Scholar] [CrossRef]
- Allen, J. Host resistance to ectoparasites. Rev. Sci. Tech. (OIE) 1994, 13, 1287–1303. [Google Scholar] [CrossRef]
- Piper, E.K.; Jackson, L.; Bagnall, N.H.; Kongsuwan, K.; Lew, A.E.; Jonsson, N.N.; Tabor, A.E. Gene expression in the skin of Bos taurus and Bos indicus cattle infested with the cattle tick, Rhipicephalus (Boophilus) microplus. Vet. Immunol. Immunopathol. 2008, 126, 110–119. [Google Scholar] [CrossRef]
- Piper, E.K.; Jonsson, N.N.; Gondro, C.; Lew-Tabor, A.E.; Moolhuijzen, P.; Vance, M.E.; Jackson, L. Immunological Profiles of Bos taurus and Bos indicus Cattle Infested with the Cattle Tick, Rhipicephalus (Boophilus) microplus. Clin. Vaccine Immunol. 2009, 16, 1074–1086. [Google Scholar] [CrossRef]
- Springell, P.H. The Cattle Tick in Relation to Animal Production in Australia; FAO: Rome, Italy, 1983. [Google Scholar]
- Mirkena, T.; Duguma, G.; Haile, A.; Tibbo, M.; Okeyo, A.; Wurzinger, M.; Sölkner, J. Genetics of adaptation in domestic farm animals: A review. Livest. Sci. 2010, 132, 1–12. [Google Scholar] [CrossRef]
- Cloete, J.J.E.; Cloete, S.W.P.; Scholtz, A.J.; Matthee, S. The effect of breed, ewe age and season on tick counts of indigenous and commercial sheep in South Africa. Proc. Assoc. Adv. Anim. Breed. Genet. 2013, 20, 187–190. [Google Scholar]
- Cloete, S.; Cloete, J.; Scholtz, A. Genetic parameters for tick count and udder health in commercial and indigenous ewes in South Africa. Vet. Parasitol. 2016, 230, 33–42. [Google Scholar] [CrossRef]
- Rjeibi, M.R.; Gharbi, M.; Mhadhbi, M.; Mabrouk, W.; Ayari, B.; Nasfi, I.; Jedidi, M.; Sassi, L.; Rekik, M.; Darghouth, M.A. Prevalence of piroplasms in small ruminants in North-West Tunisia and the first genetic characterisation of Babesia ovis in Africa. Parasite 2014, 21, 23. [Google Scholar] [CrossRef]
- Ben Said, M.; Belkahia, H.; Alberti, A.; Zobba, R.; Bousrih, M.; Yahiaoui, M.; Daaloul-Jedidi, M.; Mamlouk, A.; Gharbi, M.; Messadi, L. Molecular Survey of Anaplasma Species in Small Ruminants Reveals the Presence of Novel Strains Closely Related to A. phagocytophilumin Tunisia. Vector Borne Zoonotic Dis. 2015, 15, 580–590. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasites Vectors 2010, 3, 26. [Google Scholar] [CrossRef]
- Papa, A.; Tsergouli, K.; Tsioka, K.; Mirazimi, A. Crimean-Congo Haemorrhagic Fever: Tick-Host-Virus Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 213. [Google Scholar] [CrossRef]
- Rjeibi, M.R.; Darghouth, M.A.; Rekik, M.; Amor, B.; Sassi, L.; Gharbi, M. First Molecular Identification and Genetic Characterization of Theileria lestoquardi in Sheep of the Maghreb Region. Transbound. Emerg. Dis. 2016, 63, 278–284. [Google Scholar] [CrossRef]
- Elati, K.; Ayadi, A.A.; Khbou, M.K.; Jdidi, M.; Rekik, M.; Gharbi, M. Dynamique des populations de tiques infestant les ovins dans les steppes arides de Tunisie. Rev. Élev. Méd. Vét. Pays Trop. 2018, 71, 131. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.L.; Walker, A.R. Ticks of Domestic Animals in the Mediterranean Region. A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004; ISBN 9788496214187. [Google Scholar]
- Rubel, F.; Kottek, M. Observed and projected climate shifts 1901-2100 depicted by world maps of the Köppen-Geiger climate classification. Meteorol. Z. 2010, 19, 135–141. [Google Scholar] [CrossRef]
- Organization Climate Data. Climate Tunisia. Available online: https://fr.climate-data.org/afrique/tunisie/tunis-414/ (accessed on 10 April 2020).
- World Meteorological Organization. WMO Provisional Statement on the Status of the Global Climate in 2019; WMO: Geneva, Switzerland, 2019. [Google Scholar]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef]
- Bouchard, C.; Dibernardo, A.; Koffi, J.; Wood, H.; Leighton, P.; Lindsay, L.R. Increased risk of tick-borne diseases with climate and environmental changes. Can. Commun. Dis. Rep. 2019, 45, 83–89. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Locality (District) | Number of Surveyed Farms (Total Number of Animals) | Number of Barbarine Sheep in the Region/Total Number of Barbarine Sheep (%) | Number of QFO Sheep in the Region/Total Number of QFO Sheep (%) |
|---|---|---|---|---|
| Northeast | Mornaguia (Manouba) and Saouef (Zaghouan) | 6 (164) | 123/288 (42.7) | 41/151 (27.1) |
| Northwest | Fernana (Jendouba) and Sebeitla (Kasserine) | 6 (162) | 119/288 (41.3) | 43/151 (28.5) |
| Southeast | Bir Ali (Sfax) and Bir Lahmar (Tataouine) | 5 (113) | 46/288 (16) | 67/151 (44.4) |
| Overall | 17 (439) | 288 (100) | 151 (100) |
| Primers | Sequences (5′------3′) | Author |
|---|---|---|
| 1A | AACCTGGTTGATCCTGCCAGT | [47] |
| 564R | GGCACCAGACTTGCCCTC | |
| RLB-F | GAGGTAGTGACAAGAAATAACAATA | [48] |
| RLB-R | TCTTCGATCCCCTAACTTTC |
| Tick Species | Number of Ticks in the Breed (%) a | ||
|---|---|---|---|
| Barbarine | Queue Fine de l’Ouest | Overall (%) | |
| Rhipicephalus sanguineus s.l. | 487 (75.4) | 159 (24.6) | 646 (91.4) b |
| Hyalomma impeltatum | 31 | 0 | 31 (4.4) |
| Hyalomma excavatum | 15 | 0 | 15 (2.1) |
| Hyalomma marginatum | 5 | 0 | 5 (0.7) |
| Hyalomma dromedarii | 4 | 0 | 4 (0.6) |
| Rhipicephalus bursa | 2 | 1 | 3 (0.4) |
| Rhipicephalus annulatus | 2 | 0 | 2 (0.3) |
| Haemaphysalis sulcata | 1 | 0 | 1 (0.1) |
| Overall (%) | 547 (77.4) c | 160 (22.6) | 707 (100) |
| Variables | April 2018 | July 2018 | October 2018 | January 2019 | April 2019 | July 2019 | October 2019 | January 2020 | p |
|---|---|---|---|---|---|---|---|---|---|
| Northeast | |||||||||
| Mean temperature (°C) (Min–Max) a | 15.05 (9.7–207) | 27.34 (21.09–34.25) | 21.48 (17.2–26.73) | 10.11 (6.44–14.04) | 13.58 (8.93–18.86) | 27.96 (21.65–34.32) | 21.61 (16.68–27.03) | 9.87(6.12–13.99) | N.A. |
| Cumulative precipitation (mm) b | 16.44 | 0.5 | 8.02 | 17.12 | 44.32 | 1.58 | 19.39 | 14.43 | |
| Northwest | |||||||||
| Mean temperature (°C) (Min–Max) | 12.48 (7.31–18.23) | 22.66 (17.43–30.53) | 20.21 (15.93–25–45) | 8.02 (3.91–11.27) | 11.17 (6.81–15.99) | 27.48 (20.72–33.99) | 20.80 (15.04–27.07) | 9.71 (6.36–13.16) | N.A. |
| Cumulative precipitation (mm) | 40.44 | 4.22 | 43.99 | 33.06 | 56.06 | 1.73 | 15.37 | 17.5 | |
| Southeast | |||||||||
| Mean temperature (°C) (Min–Max) | 17.62 (12.12–23.35) | 28.7 (22.69–35.26) | 23.55 (19.11–28.71) | 10.97 (13.07–15.52) | 15.05 (11.06–19.57) | 30.9 (24.88–37.80) | 24.4 (19.9–29.63) | 10 (6.83–13.52) | N.A. |
| Cumulative precipitation (mm) | 6 | 1.6 | 19.8 | 5 | 27.17 | 0 | 14.08 | 16.93 | |
| Infestation prevalence c | 53/439 | 44/382 | 18/362 | 6/341 | 60/269 | 98/288 | 23/272 | 19/258 | <0.001 f |
| (% ± SE) | (12.1 ± 1.6) | (11.5 ± 1.6) | (5 ± 1.1) | (1.8 ± 0.7) | (22.3 ± 2.5) | (34 ± 2.8) | (8.5 ± 1.7) | (7.4 ± 1.6) | |
| Mean intensity d | 129/53 | 61/44 | 20/18 | 9/6 | 135/60 | 215/98 | 92/23 | 46/19 | <0.001 g |
| (mi ± SE) | (2.43 ± 0.2) | (1.39 ± 0.09) | (1.11 ± 0.07) | (1.5 ± 0.22) | (2.25 ± 0.22) | (2.19 ± 0.19) | (4 ± 1.16) | (2.42 ± 0.3) | |
| Mean abundance e | 129/439 | 61/382 | 20/362 | 9/341 | 135/269 | 215/288 | 92/272 | 46/258 | <0.001 h |
| (ma ± SE) | (0.29 ± 0.05) | (0.16 ± 0.02) | (0.06 ± 0.01) | (0.03 ± 0.01) | (0.5 ± 0.07) | (0.75 ± 0.09) | (0.34 ± 0.11) | (0.18 ± 0.04) |
| April 2018 | July 2018 | October 2018 | January 2019 | April 2019 | July 2019 | October 2019 | January 2020 | |
|---|---|---|---|---|---|---|---|---|
| Mean Tick Abundance | ||||||||
| April 2018 | 1 | 0.35 | 0.009 | 0.003 | 0.08 | 0.001 | 1 | 0.61 |
| July 2018 | - | 1 | 0.87 | 0.67 | 0.001 | 0.001 | 0.47 | 1 |
| October 2018 | - | - | 1 | 1 | 0.001 | 0.001 | 0.028 | 0.87 |
| January 2019 | - | - | - | 1 | 0.001 | 0.001 | 0.011 | 0.7 |
| April 2019 | - | - | - | - | 1 | 0.445 | 0.196 | 0.001 |
| July 2019 | - | - | - | - | - | 1 | 0.001 | 0.001 |
| October 2019 | - | - | - | - | - | - | 1 | 0.69 |
| January 2020 | - | - | - | - | - | - | - | 1 |
| Mean Tick Intensity | ||||||||
| April 2018 | 1 | 0.15 | 0.27 | 0.96 | 1 | 0.96 | 0.15 | 1 |
| July 2018 | - | 1 | 1 | 1 | 0.24 | 0.54 | 0.001 | 0.68 |
| October 2018 | - | - | 1 | 1 | 0.37 | 0.64 | 0.002 | 0.65 |
| January 2019 | - | - | - | 1 | 0.98 | 0.99 | 0.24 | 0.98 |
| April 2019 | - | - | - | - | 1 | 0.99 | 0.08 | 1 |
| July 2019 | - | - | - | - | - | 1 | 0.009 | 1 |
| October 2019 | - | - | - | - | - | - | 1 | 0.32 |
| January 2020 | - | - | - | - | - | - | - | 1 |
| April 2018 | July 2018 | October 2018 | January 2019 | April 2019 | July 2019 | October 2019 | January 2020 | |
|---|---|---|---|---|---|---|---|---|
| Region | ||||||||
| NE | N.S. | N.S. | N.S. | N.S. | 0.001 | 0.003 | N.S. | N.S. |
| NW | N.S. | N.S. | <0.001 | N.S. | <0.001 | N.S. | N.S. | <0.001 |
| SE R | ||||||||
| Breed | ||||||||
| Barbarine | N.S. | <0.001 | 0.02 | N.S. | <0.001 | <0.001 | <0.001 | 0.001 |
| QFO R | ||||||||
| Region × Breed | ||||||||
| NE × Barbarine | N.S. | <0.001 | N.S. | N.S. | <0.001 | 0.001 | 0.01 | N.S. |
| NE × QFO R | ||||||||
| NW × Barbarine | <0.001 | |||||||
| NW × QFO R | ||||||||
| SE × Barbarine | ||||||||
| SE × QFO R |
| Sampling Round | Number of Theileria-Babesia Positive Sheep/Number of Examined Sheep (% ± SE) | |
|---|---|---|
| Barbarine | Queue Fine de L’Ouest | |
| April 2018 | 2/286 (0.7 ± 0.5) a | 0/152 (0) b |
| July 2018 | 12/242 (4.96 ± 1.4) | 0/128 (0) |
| October 2018 | 13/219 (5.94 ± 1.6) | 0/129 (0) |
| January 2019 | 8/208 (3.85 ± 1.3) | 0/113 (0) |
| Total | 35/955 (3.66 ± 0.6) | 0/522 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khamassi Khbou, M.; Rouatbi, M.; Romdhane, R.; Sassi, L.; Jdidi, M.; Haile, A.; Rekik, M.; Gharbi, M. Tick Infestation and Piroplasm Infection in Barbarine and Queue Fine de l’Ouest Autochthonous Sheep Breeds in Tunisia, North Africa. Animals 2021, 11, 839. https://doi.org/10.3390/ani11030839
Khamassi Khbou M, Rouatbi M, Romdhane R, Sassi L, Jdidi M, Haile A, Rekik M, Gharbi M. Tick Infestation and Piroplasm Infection in Barbarine and Queue Fine de l’Ouest Autochthonous Sheep Breeds in Tunisia, North Africa. Animals. 2021; 11(3):839. https://doi.org/10.3390/ani11030839
Chicago/Turabian StyleKhamassi Khbou, Médiha, Mariem Rouatbi, Rihab Romdhane, Limam Sassi, Mohamed Jdidi, Aynalem Haile, Mourad Rekik, and Mohamed Gharbi. 2021. "Tick Infestation and Piroplasm Infection in Barbarine and Queue Fine de l’Ouest Autochthonous Sheep Breeds in Tunisia, North Africa" Animals 11, no. 3: 839. https://doi.org/10.3390/ani11030839
APA StyleKhamassi Khbou, M., Rouatbi, M., Romdhane, R., Sassi, L., Jdidi, M., Haile, A., Rekik, M., & Gharbi, M. (2021). Tick Infestation and Piroplasm Infection in Barbarine and Queue Fine de l’Ouest Autochthonous Sheep Breeds in Tunisia, North Africa. Animals, 11(3), 839. https://doi.org/10.3390/ani11030839