
New Echinoderm-Crab Epibiotic Associations from the Coastal Barents Sea



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
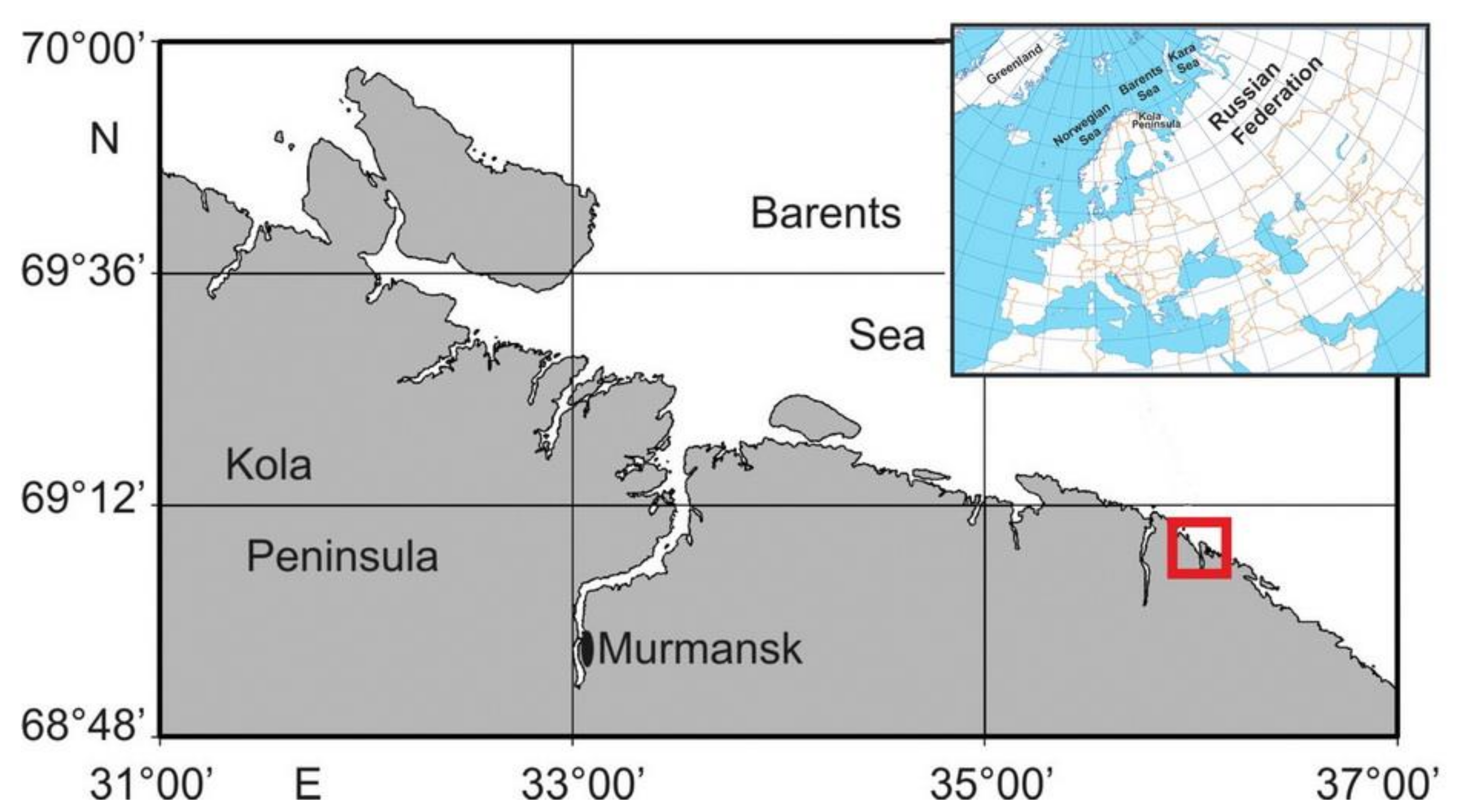
2.1. Study Area
2.2. Sampling and Processing
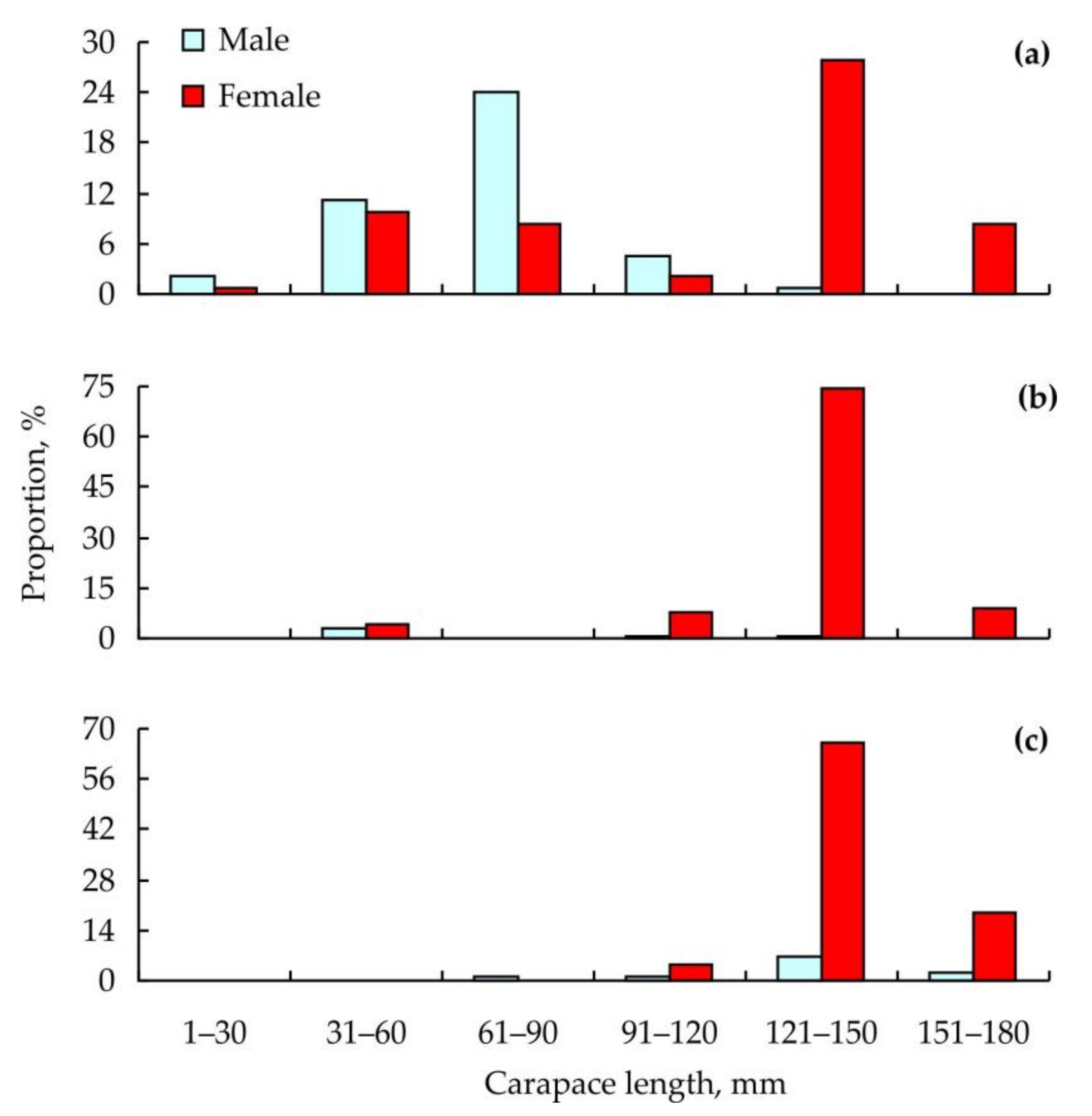
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fisheries 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (red king) Crab in the Barents Sea. Pecularities of Biology, Perspectives of Fishery; KSC RAS Press: Apatity, Russia, 2002. (in Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fisheries 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Fuhrmann, M.M.; Pedersen, T.; Nilssen, E.M. Trophic niche of the invasive red king crab Paralithodes camtschaticus in a benthic food web. Mar. Ecol. Prog. Ser. 2017, 565, 113–129. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Bakanev, S.V.; Stesko, A.V. Red king crab. In Materials of Total Allowable Catches of Water Biological Resources in Fishing Areas in Inland Seas of the Russian Federation, on the Continental Shelf of the Russian Federation, in the Exclusive Economical Zone of the Russian Federation, in the Azov and Caspian Seas in 2022; Sologub, D.O., Ed.; FGBUN VNIRO: Murmansk, Russia, 2021; pp. 2–16. (in Russian) [Google Scholar]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Fernandez-Leborans, G. Epibiosis in Crustacea: An overview. Crustaceana 2010, 83, 549–640. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Some aspects of the biology of the amphipods Ischyrocerus anguipes associated with the red king crab, Paralithodes camtschaticus, in the Barents Sea. Polar Biol. 2009, 32, 463–469. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution of amphipods Ischyrocerus on the red king crab, Paralithodes camtschaticus: Possible interactions with the host in the Barents Sea. Estuar. Coast. Shelf Sci. 2009, 82, 390–396. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epifauna associated with an introduced crab in the Barents Sea: A 5-year study. ICES J. Mar. Sci. 2010, 67, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Copepods associated with the red king crab Paralithodes camtschaticus (Tilesius, 1815) in the Barents Sea. Zool. Stud. 2013, 52, 17. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, L.V. Red king crab trophic relations and its influence on bottom biocenoses. In Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; Matishov, G.G., Ed.; KSC RAS Press: Apatity, Russia, 2008; pp. 77–104. (in Russian) [Google Scholar]
- Stevens, B.G.; Jewett, S.C. Growth, molting, and feeding of king crabs. In King Crabs of the World: Biology and Fisheries Management; Stevens, B.G., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 315–361. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Fouling community of the red king crab, Paralithodes camtschaticus (Tilesius 1815), in a subarctic fjord of the Barents Sea. Polar Biol. 2009, 32, 1047–1054. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab Paralithodes camtschaticus (Tilesius, 1815) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Miller, A.; Inglis, G.J.; Poulin, R. Comparison of the ectosymbionts and parasites of an introduced crab, Charybdis japonica, with sympatric and allopatric populations of a native New Zealand crab, Ovalipes catharus (Brachyura: Portunidae). N. Z. J. Mar. Freshw. Res. 2006, 40, 369–378. [Google Scholar] [CrossRef]
- Goedknegt, M.A.; Feis, M.E.; Wegner, K.M.; Luttikhuizen, P.C.; Buschbaum, C.; Camphuysen, K.C.J.; van der Meer, J.; Thieltges, D.W. Parasites and marine invasions: Ecological and evolutionary perspectives. J. Sea Res. 2016, 113, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Purivirojkul, W.; Songsuk, A. New records of fish parasitic isopods (Crustacea: Isopoda) from the Gulf of Thailand. Animals 2020, 10, 2298. [Google Scholar] [CrossRef]
- Torchin, M.E.; Lafferty, K.D.; Kuris, A.M. Parasites and marine invasions. Parasitology 2002, 124, 137–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquaculture 2020b, 12, 1280–1290. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs; Fairbanks, Alaska Sea Grant College Program; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- World Register of Marine Species. 2020. Available online: http://www.marinespecies.org/ (accessed on 22 December 2020).
- Stachowitsch, M. The epibiotic and endolithic species associated with the gastropod shells inhabited by the hermit crabs Paguristes oculatus and Pagurus cuanensis. P. S. Z. N. Mar. Ecol. 1980, 1, 73–101. [Google Scholar] [CrossRef]
- Cuadras, J.; Pereira, F. Invertebrates associated with Dardanus arrosor (Anomura, Diogenidae). Vie Milieu 1977, 27, 301–310. [Google Scholar]
- Parapar, J.; Fernandez, L.; Gonzalez-Gurriaran, E.; Muino, R. Epibiosis and masking material in the spider crab Maja squinado (Decapoda: Majidae) in the Ria de Arousa (Galicia, NW Spain). Cah. Biol. Mar. 1997, 38, 221–234. [Google Scholar]
- Budd, G. Asterias rubens. Common starfish. Marine Life information network: Biology and sensitivity key information sub-programme [on-line]. Plymouth: Marine Biological Association of the United Kingdom. 2008; Available online: http://www.marlin.ac.uk/speciesfullreview.php?speciesID=2657 (accessed on 21 February 2021).
- Schäfer, W. Ecology and palaeoecology of marine environments; University of Chicago Press: Chicago, IL, USA, 1972. [Google Scholar]
- Anger, K.; Rogal, U.; Schriever, G.; Valentin, C. In situ investigations on the echinoderm Asterias rubens as a predator of soft-bottom communities in the western Baltic sea. Helgol. wiss Meeresunt. 1977, 29, 439–459. [Google Scholar] [CrossRef] [Green Version]
- Sloan, N.A. Aspects of the feeding biology of asteroids. Oceanogr. Mar. Biol. Ann. Rev. 1980, 18, 57–124. [Google Scholar]
- Mortensen, T. Handbook of the Echinoderms of the British Isles; Oxford University Press: London, UK, 1927. [Google Scholar]
- Scheibling, R.E.; Hatcher, B.G. Strongylocentrotus droebachiensi. In Sea Urchins: Biology and Ecology, 3rd ed.; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 381–412. [Google Scholar]
- Shin, H.C.; Kho, C.-H. Distribution and abundance of ophiuroids on the continental shelf and slope of the East Sea (southwestern Sea of Japan), Korea. Mar. Biol. 1993, 115, 393–399. [Google Scholar] [CrossRef]
- Packer, D.B.; Watling, L.; Langton, R.W. The population structure of the brittle star Ophiura sarsii Lütken in the Gulf of Maine and its trophic relationship to American plaice (Hippoglossoides platessoides Fabricius). J. Exp. Mar. Biol. Ecol. 1994, 179, 207–222. [Google Scholar] [CrossRef]
- Harris, J.L.; MacIsaac, K.; Gilkinson, K.D.; Kenchington, E.L. Feeding biology of Ophiura sarsii Lütken, 1855 on Banquereau bank and the effects of fishing. Mar. Biol. 2009, 156, 1891–1902. [Google Scholar] [CrossRef]
- Aronson, R.B. Brittlestar beds: Low-predation anachronisms in the British Isles. Ecology 1989, 70, 856–865. [Google Scholar] [CrossRef]
- Hamel, J.-F.; Mercier, A. Population status, fisheries and trade of sea cucumbers in temperate areas of the Northern Hemisphere. In Sea Cucumbers. A Global Review of Fisheries and Trade. FAO Fisheries and Aquaculture Technical Paper. No. 516; Toral-Granda, V., Lovatelli, A., Vasconcellos, M., Eds.; FAO: Rome, Italy, 2008; pp. 257–291. [Google Scholar]
- Gudimova, E.N. Sea Cucumber Cucumaria frondosa (Gunner, 1767). In Harvesting and Perspective for uses Algae and Invertebrates of the Barents and White Seas; Matishov, G.G., Ed.; KSC RAS Press: Apatity, Russia, 1998; pp. 453–528. (in Russian) [Google Scholar]
- Gudimova, E.N.; Gudimov, A.; Collin, P. A study of the biology for fishery in two populations of Cucumaria frondosa: In the Barents Sea (Russia) and in the Gulf of Maine (USA). In Echinoderms Munchen, Proceedings of the 11th Internation Echinoderm Conference, Munich, Germany, 6–10 Octobre 2003; Heinzeller, T., Nebelsick, J.H., Eds.; A. A. Balkema Publishers: Leiden, The Netherlands, 2004; pp. 269–275. [Google Scholar]
- Poulter, R.; Oliver, P.G.; Hauton, C.; Sanders, T.; Ciotti, B.G. Infestation of shore crab gills by a free-living mussel species. Mar. Biodiv. 2018, 48, 1241–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas, M.J.; Stotz, W.; Laudien, J. First record of an epibiosis between the sand crab Emerita analoga (Stimpson, 1857) (Decapoda: Hippidae) and the mussel Semimytilus algosus (Gould, 1850) (Bivalvia, Mytilidae) in southern Peru. Helgol. Mar. Res. 2006, 60, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Pereladov, M.V. Some aspects of distribution and behavior of red king crab (Paralithodes camtschaticus), on the Barents Sea shallow coastal water. Tr. VNIRO 2003, 142, 103–119, (in Russian with English abstract). [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population biology of Ischyrocerus commensalis, a crab-associated amphipod, in the southern Barents Sea: A multi-annual summer study. Mar. Ecol. 2011, 32, 498–508. [Google Scholar] [CrossRef]
- Talberg, N.B. Comparative characteristics in the red king crab migration pattern in the shallows of the Barents and Okhotsk Sea. Tr. VNIRO 2005, 142, 91–101, (in Russian with English abstract). [Google Scholar]
- Kuris, A.M.; Blau, S.F.; Paul, A.J.; Shields, J.D.; Wickham, D.E. Infestation by brood symbionts and their impact on egg mortality in the red king crab, Paralithodes camtschatica, in Alaska: Geographic and temporal variation. Can. J. Fish. Aquat. Sci. 1991, 48, 559–568. [Google Scholar] [CrossRef]
- Gannon, A.T.; Wheatly, M.G. Physiological effects of an ectocommensal gill barnacle, Octolasmis muelleri, on gas exchange in the blue crab Callinectes sapidus. J. Crustac. Biol. 1992, 12, 11–18. [Google Scholar] [CrossRef]
- Williams, J.D.; McDermott, J.J. Hermit crab biocoenoses: A worldwide review of the diversity and natural history of hermit crab associates. J. Exp. Mar. Biol. Ecol. 2004, 305, 1–128. [Google Scholar] [CrossRef]
- Conover, M.R. The influence of some symbionts on the shell-selection behaviour of the hermit crabs, Pagurus pollicarus [sic] and Pagurus longicarpus. Anim. Behav. 1976, 24, 191–194. [Google Scholar] [CrossRef]
- Taylor, P.D. Observations on symbiotic associations of bryozoans and hermit crabs from the Otago Shelf of New Zealand. Bull. Soc. Sci. Nat. Ouest Fr. Mem. 1991, 1, 487–495. [Google Scholar]
- Hemmingsen, W.; Jansen, P.A.; MacKenzie, K. Crabs, leeches and trypanosomes: An unholy trinity? Mar. Pollut. Bull. 2005, 50, 336–339. [Google Scholar] [CrossRef]

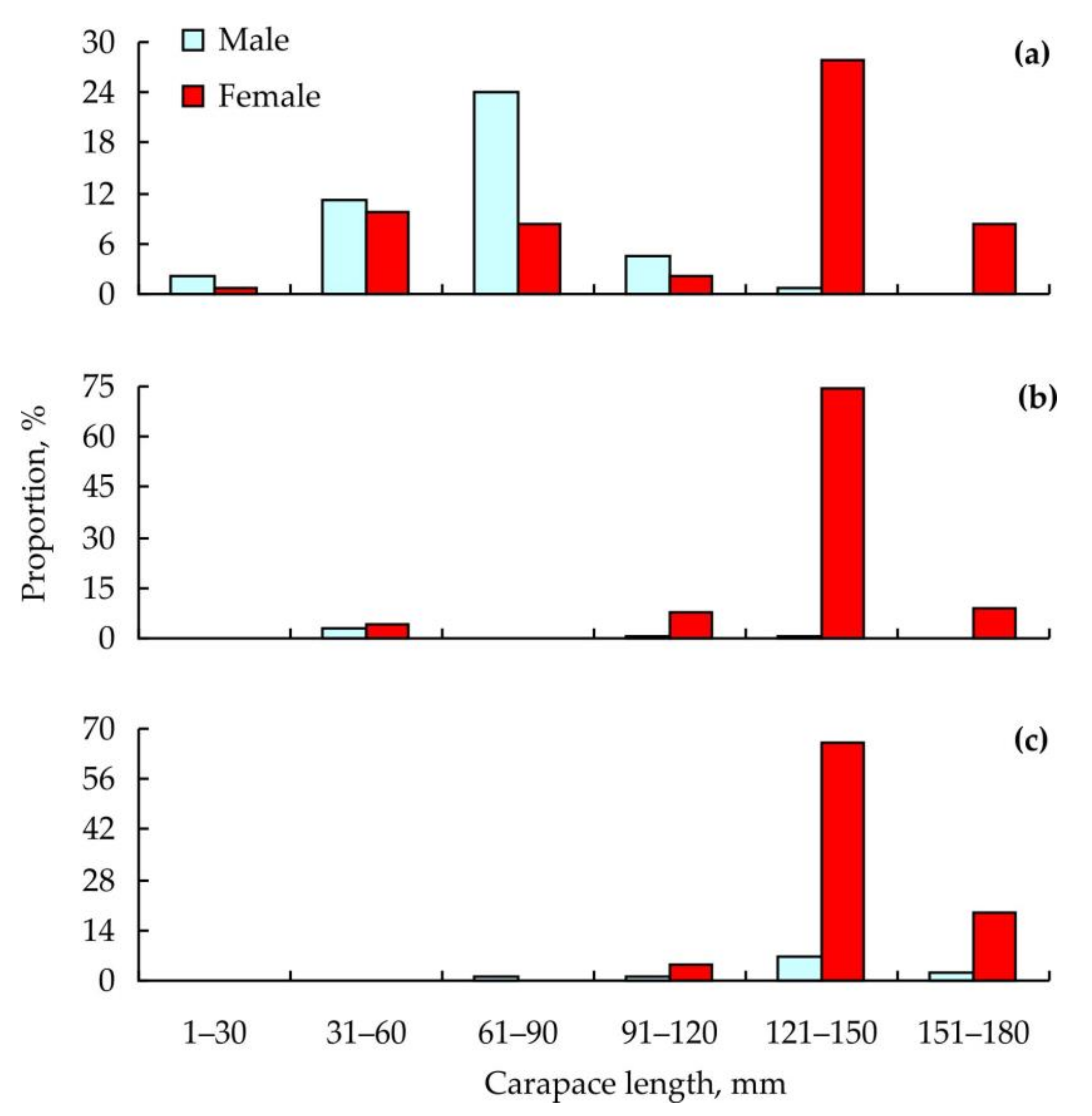


{kind=link}
{kind=link}
{kind=link}
| Other Epibionts | Echinoderm Epibiont | |||||
|---|---|---|---|---|---|---|
| Sea Star | Sea Urchin | Sea Star | Sea Star | Brittle Star | Sea Cucumber | |
| 2010 | 2010 | 2018 | 2019 | 2019 | 2019 | |
| Ampelisca sp. | – | – | – | – | 3 | – |
| Balanus crenatus Bruguière, 1789 | – | 2 | ||||
| Callopora lineata (L., 1767) | – | – | + | – | + | – |
| Caprella sp. | – | – | – | – | 1 | – |
| Chone sp. | – | – | – | 1 | – | |
| Craniella cranium (Müller, 1776) | – | – | – | – | 1 | – |
| Coryne hincksi Bonnevie, 1898 | – | 1 | – | – | – | – |
| Crisia denticulata (Lamarck, 1816) | – | – | + | – | + | – |
| Disporella hispida (Fleming, 1828) | – | – | + | – | + | – |
| Ectinosoma normani Scott T. & A., 1896 | – | – | – | 4 | – | – |
| Eumida sanguinea (Örsted, 1843) | – | – | – | – | 1 | – |
| Harmothoe imbricata (L., 1767) | – | – | – | – | 2 | – |
| Harpacticus chelifer (O.F. Müller, 1776) | – | – | – | – | – | 1 |
| Ischyrocerus anguipes Krǿyer, 1838 | – | – | – | – | 71 | 1 |
| Ischyrocerus commensalis Chevreux, 1900 | 6 | 5 | 18 | 51 | 113 | 68 |
| Johanssonia arctica (Johansson, 1898) | 1 | – | – | – | 2 | – |
| Lacuna vincta (Montagu, 1803) | – | – | – | – | 1 | – |
| Mesochra pygmaea (Claus, 1863) | – | – | – | 4 | – | – |
| Obelia longissima (Pallas, 1766) | – | + | – | – | + | – |
| Patinella verrucaria (Linnaeus, 1758) | – | – | – | – | + | – |
| Semibalanus balanoides (Linnaeus, 1767) | – | – | – | – | 40 | – |
| Tisbe furcata (Baird, 1837) | – | 1 | 45 | 20 | 251 | 17 |
| Tisbe minor (Scott T. & A., 1896) | – | – | – | – | – | 3 |
| Tisbe tenera (Sars G.O., 1905) | – | – | 20 | – | 4 | |
| Tricellaria arctica (Busk, 1855) | – | + | + | – | + | – |
| Zaus spinatus (Goodsir, 1845) | – | – | – | 1 | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Dvoretsky, V.G. New Echinoderm-Crab Epibiotic Associations from the Coastal Barents Sea. Animals 2021, 11, 917. https://doi.org/10.3390/ani11030917
Dvoretsky AG, Dvoretsky VG. New Echinoderm-Crab Epibiotic Associations from the Coastal Barents Sea. Animals. 2021; 11(3):917. https://doi.org/10.3390/ani11030917
Chicago/Turabian StyleDvoretsky, Alexander G., and Vladimir G. Dvoretsky. 2021. "New Echinoderm-Crab Epibiotic Associations from the Coastal Barents Sea" Animals 11, no. 3: 917. https://doi.org/10.3390/ani11030917
APA StyleDvoretsky, A. G., & Dvoretsky, V. G. (2021). New Echinoderm-Crab Epibiotic Associations from the Coastal Barents Sea. Animals, 11(3), 917. https://doi.org/10.3390/ani11030917