1. Introduction
Global warming, caused by the increasing production of greenhouse gases from different sources including agriculture and livestock, is of great global concern [
1,
2]. Therefore, it is a prerequisite to increase the number of heat tolerant breeds globally. This is because sustainable animal production depends on environmental temperature, and thermoneutrality is needed for normal metabolism and physiological activities. Both seasonal stressors, either cold or heat, can negatively affect animal performance [
3,
4,
5,
6,
7,
8,
9]. During cold stress, increased maintenance energy is required to retain body temperature, and feed efficiency is greatly hampered [
3]. In contrast, it is not uncommon for cattle to reduce dry matter intake (DMI) and rumen motility during heat stress [
10]. Jersey cows had better adaptation capabilities to heat stress compared to Holstein cows [
11,
12]; however, Holstein cows are well adapted to lower temperatures (the lower and upper critical temperature varies from −15 °C to 22 °C) [
13]. It is well known that Holstein and Jersey are two important dairy breeds, and the contribution steers of these breeds make to beef production is of considerable value [
14,
15]. However, most previous studies have focused on dairy breeds and little is known about the steers of dairy breeds.
Members of the rumen microbiota, including bacteria, protozoa, fungi, and archaea, can ferment a wide variety of ingested feedstuffs to subsequently produce volatile fatty acids (VFAs), such as acetate, propionate, and butyrate, which are then absorbed by the cattle for energy metabolism and protein synthesis [
16,
17,
18,
19]. Simultaneously, carbon dioxide (CO
2), hydrogen (H
2) and formic acid are produced as end products. Enteric methane (CH
4) can also be produced by methanogens through methanogenesis [
20,
21,
22]. CH
4 is an indicator of dietary gross energy losses, and it has a negative environmental impact contributing to global warming [
23,
24]. Previous studies revealed that several factors, including diet, feed additives, host genetics, age, and physiological state affect the rumen microbiomes, rumen fermentation characteristics, and CH
4 production [
25,
26,
27,
28,
29,
30,
31,
32,
33,
34]. O’Hara et al. [
28] reported an association between the rumen microbiome and its fermentation products with feed efficiency and CH
4 emissions. They also reported that Firmicutes, Bacteroidetes, and Proteobacteria were the dominant bacterial phyla that can ferment a wide variety of dietary carbohydrates and peptides. However, few studies have focused on the seasonal influence on the rumen microbiome and CH
4 emissions. Li et al. [
35] conducted an experiment examining seasonal effects on microbial diversity in the feces of Holstein dairy cows and stated that fecal microbial diversity and composition varied at different temperature humidity index (THI) values. Noel et al. [
36] reported a shift in the digesta-adherent rumen microbiome of Holstein dairy cow grazing pastures over the seasons. However, the fecal microbiome in the former study and the pasture grazing cattle of the other study did not completely represent the rumen microbiome changes in feedlot cattle over the seasons. Moreover, the majority of previous studies have focused on dairy cattle, whereas steers are much less researched. To the best of our knowledge, no previous study has examined the influence of seasonal stress on the rumen microbiomes, its fermentation parameters, and enteric CH
4 emissions of Holstein and Jersey steers fed the same total mixed ration (TMR). We hypothesized that season and breed can influence the rumen microbiota, rumen fermentation, and enteric CH
4 emissions. In this context, the present study was conducted (i) to evaluate whether the rumen fermentation characteristics and enteric methane emissions of Holstein and Jersey steers fed the same TMR over different seasons are similar and (ii) to determine to what extent the diversity and composition of the rumen microbiome vary between breeds and among seasons.
4. Discussion
Both breed and season can affect the growth, rumen fermentation, methane emissions, feed utilization, and other animal productivity traits of ruminants in an age-dependent manner [
25,
26,
30,
31,
32]. Many studies have compared and evaluated the difference between breeds and seasons [
27,
36,
53]. The present study compared and examined how breed and season might affect the growth performance, rumen fermentation characteristics, methane emissions, and the rumen microbiota using both Holstein and Jersey steers as animal models. To eliminate age and diet as confounding factors, all the animals used in the present study had the same age and consumed the same TMR. The results of this study provided some basic information on the effects of host genetics and physiology on some of the important traits of ruminants.
As expected, Holstein steers had higher total DMI than Jersey steers, which agrees with the results of Flay et al. [
54] who reported that the heavier breed of Holstein heifers had higher DMI than their Jersey counterparts. However, Holstein steers had the lowest metabolic DMI in summer, while the lowest total DMI of Jersey steers was observed in winter. This might be due to the higher and lower THI recorded during summer and winter, leading to heat and cold sensitivity of Holstein and Jersey steers, respectively [
13]. It was reported earlier that seasonal changes influenced dietary composition and intake of grazing beef steers [
55,
56]; however, this study offered the same TMR throughout the feeding trial, which indicates the seasonal influence of intake of feedlot steers. Ruminal pH decreases with an increase in VFA production by microbial fermentation or decreases in VFA absorption via the ruminal epithelium or saliva secretion [
57]. In the present study, the significantly lower pH observed in summer may be due to decrease in saliva secretion and increase in saliva drooling, which is often observed in animals exposed to high THI [
58]. However, the gradual decrease in the concentrations of total VFA and propionate proportion concomitant with the increase in THI may be associated with disturbances in microbial activity in the rumen during summer, which was supported by decrease in OTU abundances in summer, as observed in this study. Earlier studies also reported a significant decrease in VFA production during heat stress conditions [
59,
60,
61]. In contrast, butyrate absorption through the rumen epithelium could greatly decrease under heat stress [
62], which corroborates the significantly higher butyrate concentration observed in the present study during high THI in summer. Both seasons and breeds also had a significant influence on the A:P ratio. Holstein steers had a lower A:P ratio, and the lowest value occurred in winter. This might be due to the significantly higher propionate production in winter and by Holstein steers in the current study. Rumen NH
3-N concentrations can be influenced by dietary protein breakdown, NH
3 utilization by rumen microbes, absorption by rumen wall, and urea hydrolysis in the rumen [
63,
64,
65]. The highest NH
3-N concentration observed in summer might be explained by the decrease in its absorption by the rumen wall and utilization by rumen microbes, which may have been greatly affected by high THI during summer. However, the differences in rumen NH
3-N concentration between the Jersey and Holstein steers could also be attributed to variation in rumen microbiota in the two breeds.
It was hypothesized that Holstein steers would produce more CH
4 owing to their higher DMI, while Jersey steers should yield less CH
4 due to their greater feed efficiency [
66]. However, we observed numerically higher CH
4 production and significantly higher CH
4 yield and intensity in Jersey steers than in Holstein steers. This finding is in agreement with the results of Olijhoek et al. [
67], who reported that CH
4 yield was significantly higher in Jersey cows than in Holstein cows. Propionate-producing rumen microbes compete with methanogens to metabolize H
2, thereby lowering methane production [
68,
69]. The lower CH
4 emissions observed in the Holstein steers could be due to the significantly higher propionate production compared to Jersey steers. In addition, the A:P ratio was higher in high CH
4 producing Jersey dairy cows [
67]. Likewise, our study showed a higher A:P ratio in high CH
4-producing Jersey steers than in Holstein steers regardless of the season. However, the significantly lower A:P ratio in the winter season might be due to the significantly higher rate of propionate production.
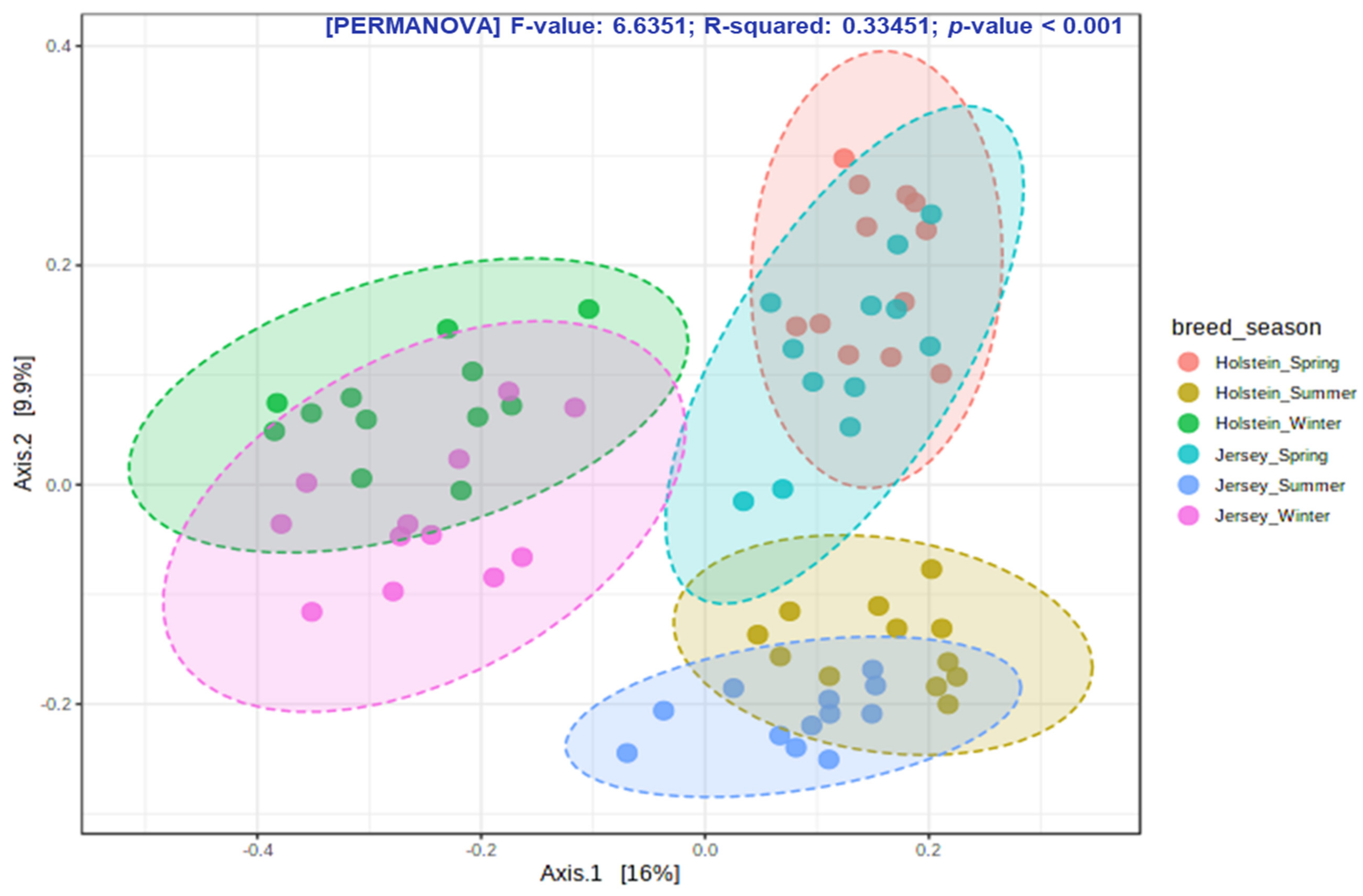
The rumen microbiota responds to variations in host genetics [
70], physiological status [
25], and diet, among other factors [
32]. Its seasonal variation independent of alteration of diet has not been well studied. In the present study, we comparatively examine how season affect the rumen microbiota in both Holstein and Jersey steers fed the identical TMR. Spring witnessed the highest number of observed OTUs and highest Chao 1 richness estimate compared to other seasons; however, summer had highly unstable values irrespective of breed. These results suggest that the rumen microbiota was rich in species when animals are free of stress during spring but that the higher THI in summer could lead to heat stress and significantly affect species richness of the rumen microbiota. The highest Shannon diversity index, which is determined by both species richness and evenness, recorded in the summer season suggests that the rumen environment of heat-stressed steers is more suitable for the proliferation of diverse group of microbes. However, the lowest diversity in the winter season might be attributed to the selective proliferation of microbes mostly associated with high metabolic heat production, which is necessary to maintain homeothermy. The PCoA showed that the composition of the rumen microbiota during winter was different from that of the other seasons. Moreover, seasonal shifting of overall rumen microbiome was observed between spring and summer. The above-mentioned findings confirm seasonal influence on the rumen microbiota in this study. Seasonal shifting of the rumen microbiota has been reported in grazing dairy cows by Noel et al. [
36], but the changes in pastures confounded any potential seasonal effect. Martinez-Fernandez et al. [
53] also observed that the bacterial community of grazing cattle had changed at mid-dry and wet season with or without a Nitrogen-based supplement. However, the differences in nutrient contents of grazing pastures at different seasons does not represent the seasonal influence of feedlot cattle with same TMR. Usually, cattle alter their energy requirement and body physiology along with the increase and decrease in ambient temperature through a variety of mechanisms. During heat stress, animal core body temperature increases about 1 °C and cattle start thermal homeostasis by increasing sweating, panting, and respiration rate phenotypically. Cattle stimulate the appetite center to reduce feed intake, which leads to reduced rumen motility. Moreover, decreased pH, ruminal absorption of fermented products, and increased rumen temperature change the rumen environment [
58,
71,
72,
73]. In contrast, cold stressed-cattle require more energy to maintain homeothermy, which is primarily achieved by increased feed intake and more metabolic heat production [
3]. Though the rumen microbiome is the key player in the rumen ecosystem, we hypothesized that the dominancy of rumen microbes might change along with the alteration of rumen environment in different season. Bacteroidetes and Firmicutes are the most abundant group of bacteria in ruminants [
74,
75,
76,
77,
78,
79]. Similarly, in the current study, Bacteroidetes and Firmicutes were the most abundant bacterial phyla in both steers regardless of season; however, the highest relative abundance of Bacteroidetes was recorded in spring and summer, while that of Firmicutes was recorded in winter. Likewise, seasonal variation of some other phyla was seen to exist in this study. These variations in the relative abundance suggest that the rumen ecosystem might be altered according to seasonal changes. Previous studies reported that
Prevotella was the most abundant bacterial genus in ruminants [
77,
78,
79]. Similarly, in the present study,
Prevotella was the most abundant bacterial genus in all seasons; however, the relative abundance of this genus was significantly higher than that of
P. brevis and
P. copri, which were observed in spring, suggesting their preferential growth in the rumen of steers during spring with normal THI. The
C. jeotgali can metabolize various carbohydrates as energy sources [
80]. In the present study, the genus
Carnobacterium and the species
C. jeotgali were only found in winter with a higher percentage of relative abundance, which might be associated with higher VFA production in winter. The species
R. bromii, belonging to the genus
Ruminococcus, family
Ruminococcaceae and phylum Firmicutes, is a starch degrading bacteria present in the rumen [
81,
82]. The higher abundance of
R. bromii in winter suggests higher amylolytic activity through their preferential growth in the rumen, which might be attributed to higher VFA production in winter. Baek et al. [
83] revealed that heat stress reduced the abundance of fibrolytic Ruminococcaceae while increasing the lactate-producing Lactobacillaceae and amylolytic
Prevotella and
Ruminobacter in Hanwoo steers. Likewise, Zhao et al. [
84] reported that heat-stressed dairy cows had a significantly higher relative abundance of
Streptococcus, unclassified
Enterobacteriaceae,
Ruminobacter,
Treponema, and unclassified Bacteroidaceae. Similarly, in the present study, both steers had higher relative abundance of
Treponema, Paludibacter,
Pal. propionicigenes during summer, suggesting their suitable growth environment in the rumen of steers during high THI. Moreover, both steers had some other distinct bacterial genera and species with higher relative abundance in different seasons, further confirming the seasonal influence. Therefore, rumen microbial richness, diversity, and community composition were greatly altered according to seasonal stress, either cold or heat, even when cattle were fed the same TMR.
Holstein and Jersey steers did not differ in numbers of observed OTUs, Chao 1 richness estimate, Shannon, and inverse Simpson diversity indexes, which is in contrast to the report of Paz et al. [
27], which reported significantly higher alpha diversity metrics, including Chao1 richness estimates and the number of observed OTUs, in Holstein cows than in lactating Jersey cows. This discrepancy may be due to the variation in the microbiota affecting the host genetics and other factors, especially sex [
70,
85] and physiological state [
86]. Similar to seasons, Bacteroidetes and Firmicutes were the most predominant bacterial phyla in the rumen of both the Holstein and Jersey steers but did not differ significantly between breeds. The relative abundances of the phyla Proteobacteria, the genera
Succinivibrio and
Gilliamella, and the species
S. dextrinosolvens,
G. bombicola,
B. clarus, and
P. enoeca were higher in the Holstein steers, while the genera
Flintibacter,
Barnesiella,
Capnocytophaga, UCG_
Ruminococcaceae,
Enterocloster, and
Oscillibacter, and the species
Cap. cynodegmi,
Fl. Butyricus,
O. ruminantium, and
Clostridium methylpentosum were more abundant in Jersey steers, suggesting their preferential growth in the rumen of particular breeds.
Fl. Butyricus and
O. ruminantium can produce butyrate from carbohydrates [
87,
88]. In our study, a significantly higher butyrate concentration was observed in Jersey steers, which may be associated with the higher relative abundance of these bacteria in this breed. Several species of the genus
Succinivibrio,
S. dextrinosolvens in particular, produce succinate, which can subsequently be converted to propionate by propionate-producing bacteria [
89,
90]. In the present study, Holstein steers had a significantly higher relative abundance of
S. dextrinosolvens than Jersey steers, which was consistent with the relative propionate concentrations and CH
4 emissions in the two breeds. The differences in the above-mentioned microbial abundances suggest that breed has a significant influence on rumen microbial community composition even when the same TMR diet is given.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}