
Clinical Cytogenetics of the Dog: A Review


Department of Genetics and Animal Breeding, Poznan University of Life Sciences, 60-637 Poznan, Poland
*
Author to whom correspondence should be addressed.
Animals 2021, 11(4), 947; https://doi.org/10.3390/ani11040947
Submission received: 27 February 2021
/
Revised: 22 March 2021
/
Accepted: 22 March 2021
/
Published: 27 March 2021
(This article belongs to the Special Issue Cytogenetics of Domestic Animals: Clinical, Molecular and Evolutionary Aspects)
Abstract
:Simple Summary
The cytogenetic analysis of dogs is mainly focused on the diagnosis of disorders of sex development (DSD) and cancers. Unfortunately, the study of canine chromosomes is a challenging task due a high chromosome number (2n = 78) and the one-arm morphology of all autosomes. For years, the application of conventional cytogenetic techniques, Giemsa staining and G and DAPI (4′,6-diamidino-2-phenylindole) bandings, allowed the identification of sex chromosome aneuploidies and centric fusions. An advanced clinical cytogenetic diagnosis is also needed due to the fact that the dog is a valuable animal model in biomedical research. The application of hybridization methods, such as fluorescence in situ hybridization (FISH) and array comparative genome hybridization (aCGH), facilitated the detection of other chromosomal rearrangements. It can be foreseen that a wide use of modern molecular techniques (e.g., SNP microarray and next generation sequencing) will substantially extend the knowledge on canine chromosome mutations.
Abstract
The dog is an important companion animal and has been recognized as a model in biomedical research. Its karyotype is characterized by a high chromosome number (2n = 78) and by the presence of one-arm autosomes, which are mostly small in size. This makes the dog a difficult subject for cytogenetic studies. However, there are some chromosome abnormalities that can be easily identified, such as sex chromosome aneuploidies, XX/XY leukocyte chimerism, and centric fusions (Robertsonian translocations). Fluorescence in situ hybridization (FISH) with the use of whole-chromosome painting or locus-specific probes has improved our ability to identify and characterize chromosomal abnormalities, including reciprocal translocations. The evaluation of sex chromosome complement is an important diagnostic step in dogs with disorders of sex development (DSD). In such cases, FISH can detect the copy number variants (CNVs) associated with the DSD phenotype. Since cancers are frequently diagnosed in dogs, cytogenetic evaluation of tumors has also been undertaken and specific chromosome mutations for some cancers have been reported. However, the study of meiotic, gamete, and embryo chromosomes is not very advanced. Knowledge of canine genome organization and new molecular tools, such as aCGH (array comparative genome hybridization), SNP (single nucleotide polymorphism) microarray, and ddPCR (droplet digital PCR) allow the identification of chromosomal rearrangements. It is anticipated that the comprehensive use of chromosome banding, FISH, and molecular techniques will substantially improve the diagnosis of chromosome abnormalities in dogs.
1. Introduction
The dog is the most important companion animal species, and one for which extreme interbreed phenotypic variation has arisen over the last 200 years [1]. One side effect of this intensive breeding, caused by genetic drift, is the preservation of undesired mutations in the gene pool. About 400 DNA variants that cause hereditary diseases in dogs have been described (Online Mendelian Inheritance in Animals—OMIA, https://omia.org/home/, (accessed on 20 December 2020)).
Our knowledge of canine chromosomal mutations is less advanced, as the chromosome set of this species is a very difficult analytic subject. The diploid number of chromosomes is high (2n = 78), but the genome size of this species (2.4 Gb) is similar to other domestic mammals. The majority of chromosomes are thus small, and their banding patterns do not allow for unambiguous recognition of all homologs. Sex chromosomes are biarmed and easily recognizable, but all autosomes are acrocentric, and only the largest pair of chromosome 1 (CFA1, Canis Familiaris chromosome 1) is distinctly different from the other autosomes.
There have been several attempts to arrange a reference banded karyotype of the dog (review by [2]). In 1993, the DogMap consortium, which was focused on the development of the canine marker genome map, suggested that a commonly accepted chromosome nomenclature for this species be established by a group of cytogeneticists experienced in canine chromosomes research. The international committee agreed that, due to the similarity of G-banding patterns of the small autosomes, only the largest 21 autosome pairs and the sex chromosome pair can be recognized with certainty. As a result of this work, a partial standard karyotype was developed [3]. An important step in characterizing the canine chromosomes was the use of a set of chromosome-specific painting probes for all autosomes and sex chromosomes [4]. Unfortunately, these probes are not available for diagnostic purposes. BAC (Bacterial Artificial Chromosome) probes with known chromosomal localization are very useful tools in clinical cytogenetics. Canine BAC clones can be purchased from the BAC libraries: CHORI-82 Canine boxer (Canis familiaris) BAC library and RPCI-81 Canine BAC Library (https://bacpacresources.org/clones.htm, accessed on 2 March 2021). Information on the localization of the CHORI-82 BAC clones is available in the CanFam3.1 assembly in NCBI - National Center for Biotechnology Information (Genome Data Viewer, accessed on 2 March 2021). Moreover, chromosome specific BAC clones from the RPCI-81 library were cytogenetically assigned [5,6]. Such probes can prove very helpful in recognizing all chromosomes, including the small autosomes (pairs 22–38) that are not included in the standard partial karyotype. However, it should be pointed out that conventional Giemsa staining is sufficient to identify sex chromosome aneuploidies and XX/XY leukocyte chimerism. Detecting centric fusions is also easy on Giemsa stained metaphase spreads, but identifying the autosomes involved requires the use of chromosome banding or fluorescence in situ hybridization (FISH).
2. Sex Chromosome Aneuploidies
Although sex chromosome aneuploidies are an important cause of infertility and sterility, they have rarely been observed in dogs (Table 1). X monosomy has been reported in five female dogs to date, usually presenting abnormal estrus cycle and small ovaries, with no evidence of ovarian follicle development. Other abnormalities, such as small stature, juvenile appearance, or excessive skin in the ventrum of the neck (typical of Turner syndrome in women), were observed only in dogs with a pure monosomy, 77,X [8,9]. The vertical septum in the vagina, observed in a single case, could be coincidental [10]. The frequency of the monosomic cell line in individuals with mosaic karyotypes (77,X/78,XX) varied over a wide range, from 5% [10] to 95% [11]. This shows that a large number of metaphase spreads need to be evaluated in infertile female dogs suspected of X monosomy.
The low incidence of X monosomy in female dogs is probably associated with the large size of the pseudoautosomal region (PAR), estimated at 6.4 Mb, which is more rich in genes than human or equine PAR [12,13]. The loss of one X chromosome is associated with the lack of a long PAR, leading to haploinsufficiency in a long genomic segment responsible for embryonic mortality. No correlation between PAR size and X trisomy was observed, which indicates that the overdose of PAR-located genes has no effect on the phenotype of X trisomy carriers [12].
Only a few females with X trisomy have been described in dogs, and they usually had abnormal estrous cycles and hypoplastic ovaries (Table 2). Interestingly, among the six reported cases with X trisomy, three females showed behavioral problems, such as fearfulness, lack of barking, or coprophagy. The majority of these cases had only a single cell line, 79,XXX. A mosaic 79,XXX/78,XX karyotype was incidentally diagnosed in a female dog with normal estrus [14]. It is worth mentioning that trisomic cell lines occurred in a low frequency (5%) of cells, which suggests that the frequency of the mosaic karyotype (79,XXX/78,XX)—associated with normal fertility or subfertility—may be underestimated.
The XXY complement has been found in six dogs, including two cases of XX/XXY mosaicism (Table 3). This abnormality is known as a cause of hyperplastic testes and sterility. However, other abnormalities, such as congenital heart disease and bilateral cryptorchidism, have also been described [19,20,21]. In some XXY dogs, testicular tumors were diagnosed [20,22] and the feminization of such dogs has also been reported [23].
3. Leukocyte Chimerism XX/XY
The XX/XY leukocyte chimerism, which is caused by the formation of anastomoses between the placentas of heterosexual fetuses, is associated with freemartinism, a form of disorder of sex development (DSD). The anastomoses enable the exchange of hematopoietic cells and molecules involved in sex differentiation between the fetuses [25]. The masculinizing factors (Sex Determining Region Y - SRY, which is a transcription factor; anti-Mullerian hormone and testosterone) produced by the fetal testes alter the sexual differentiation of the female fetus. This syndrome is well known in ruminants [26], but is also observed in other species, including dogs (Table 4). Between-species differences in the frequency of the chimerism are associated with the type of placenta: the high incidence of anastomoses in ruminants is associated with a cotyledonary organization of the placenta, while in carnivores the incidence is much lower due to the zonary organization of the placenta. It also seems that diffused placentas may be associated with an elevated risk of anastomoses in litters with a large number of fetuses, as has been observed in highly prolific lines of pigs [27]. This may suggest that in dogs, too, a greater number of puppies in a litter may be associated with a higher incidence of freemartinism.
The appearance of external genitalia is a major criterion for identifying DSD in dogs, but is less useful in freemartin dogs. Some freemartins present almost normal female genitalia [28], while others have a normal male appearance [29,30]; in sporadic cases there are ambiguous external genitalia [31].
It is important to point out that virilization can be caused by freemartinism or by testicular or ovotesticular XX DSD; a correct diagnosis should thus be made using cytogenetic analysis. A comprehensive study of six French bulldogs with ambiguous external genitalia revealed that five were testicular or ovotesticular XX DSD, while one was a freemartin [32]. Using the nomenclature of DSD dogs, XX/XY leukocyte chimerism can be considered testicular, ovarian, or ovotesticular DSD [33]. These forms are also observed in dogs and, as in ruminants, there is no correlation between the percentage of XY cells and the extent of the virilization. In the reported cases, the proportion of the XY cell line ranged from 15% to 80% (Table 4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 4.
Leukocyte chimerism XX/XY reported in dogs.
| Proportion of XX and XY Cell Lines [%] | Breed | Phenotypic Sex Considered by Owners | Characteristic Feature of Phenotype | Reference |
|---|---|---|---|---|
| Lack of information | Schipperke | female | Enlarged clitoris, testis and ovotestis, uterus, | [34] |
| 43/57 | Pug | female | Enlarged clitoris, hypospadias, no signs of estrus, testis and ovotestis | [23] |
| 45/55 | Dachshund | male | Abnormal urogenital tract, hematuria, ovaries | [35] |
| Lack of information | Spaniel × Papillon | unknown | Ovaries | [36] |
| Lack of information | American Eskimo | female | Normal vulva and clitoris, ovotestis | [37] |
| Lack of information | Spaniel | unknown | Small penis, empty rudimentary scrotum, uterus, ovaries with reduced number of follicles | [38] |
| 85/15 | Belgium Shepherd | male | Aggressive behavior, intersexuality, abdominal testes, underdeveloped penis, urethra ended under the anus, vas deferens connected to an oviduct, blind uterus | [29] |
| Lack of information | Fila Brasileiro | male | Prepuce-like structure located closer to the anus, testicles with an immature epididymides | [39] |
| 43/57 | Border Terrier | male | Undeveloped penis, ovarian-like structure | [30] |
| 70/30 | Wirehaired Pointing Griffon | female | Primary anestrus, juvenile vulva, enlarged clitoris, testis | [28] |
| 78/22 | Shih Tzu | ambiguous | Residual penis with a prepuce located in a position typical of a male, prostate, gonads remained undetectable | [31] |
| 20/80 | French Bulldog | female | Enlarge clitoris, ovotestes | [32] |
| 30/70 | Great Dane | female | Underdeveloped internal reproductive organs, rudimentary testicles | [40] |
| 54/46 | Great Dane | female | Undeveloped foreskin | [14] |
The diagnosis of XX/XY leukocyte chimerism also requires the analysis of another tissue, such as hair follicles or buccal epithelial cells, in order to distinguish between leukocyte chimerism and whole-body chimerism. Moreover, it facilitates the establishment of concordance between phenotypic and chromosomal sex. Since external genitalia of freemartins are often ambiguous, it cannot be excluded that some cases are incorrectly considered by owners as males. In Table 4, two cases were described as males, but this was not confirmed by cytogenetic or molecular studies of other tissue.
A common diagnostic strategy involves the cytogenetic analysis of leukocytes (Figure 1), the molecular detection of Y-linked genes (e.g., SRY and ZFY), or microsatellite markers in DNA samples derived from the blood and the second tissue [31]. Droplet digital PCR (ddPCR) has recently been demonstrated to be a fast and reliable method for detecting XX/XY leukocyte chimerism in cattle and pigs [27,41]. This method can also be recommended for DSD diagnosis in dogs.
4. Structural Chromosome Rearrangements
Robertsonian translocations (centric fusions) have been reported quite often in dogs, probably due to their ease of identification. Such mutations lead to a reduction in the diploid chromosome number to 77 and the formation of a biarmed derivative (der) chromosome (Figure 2). The first Robertsonian translocations in dogs were described in the 1960s, and over a dozen different translocations have now been reported in total (Table 5). Different chromosomes are involved in these mutations, but in several cases no attempt was undertaken to indicate the autosomes involved. Two autosomes, named CFA13 and CFA23 (Canis Familiaris chromosome 13 and 23), were identified more often, but this finding should be taken with caution due to the difficulties in identifying small autosomes when only chromosome banding was used. There is only a single report on the use of FISH with locus-specific probes to describe a centric fusion, which found rob(5;23), as described by Switonski et al. [42].
Dogs with Robertsonian translocations present normal phenotype, but there is often a small decrease in fertility. However, this abnormality has also been diagnosed in infertile bitches [43,44,45], which either showed a lack of estrus or were unsuccessfully mated many times. Moreover, two dogs with persistent Müllerian duct syndrome (PMDS) were found to have centric fusions, but coincidentally [46,47]. An interesting case of centric fusion in a testicular/ovotesticular XX DSD (SRY-negative) female dog with an enlarged clitoris and uterus was described by Switonski et al. [42]. CFA5 and CFA23 were involved in this rearrangement and the fusion led to a pericentromeric fragment of CFA23 being deleted. It was hypothesized that this could cause the deletion of regulatory sequences for genes that are important in ovarian development located in CFA23, such as PISRT1, FOXL2, and CTNNB1.
Table 5.
Robertsonian translocations reported in dogs.
| Chromosome Involved in the Fusion | Breed (Number of Cases) | Characteristic Feature of Phenotype | Reference |
|---|---|---|---|
| Not identified | Mixed terrier (1) | Cardiac defect | [48] |
| Not identified | Miniature Poodle (1) | Bone chondrodysplasia | [49] |
| Not identified | Setter–Retriever cross (1) | Phenotypically and clinically normal female | [50] |
| 13 and 17 | Golden Retriever cross (1) | Normal, fertile female | [51] |
| 13 and 23 | Golden Retriever-type (1 + 11 offspring) | Normal phenotype with the exception of congenital inguinal hernia in two female homozygotes in progeny | [52] |
| 1 and 31 | Poodle (6, including 1 homozygote male) | Normal phenotype | [53] |
| 21 and 33 | Walker Hound (1 + sister and 4 offspring) | Narrow vulva, absence of estrus | [43] |
| Not identified | Mixed breed (1) | Infertile female | [44] |
| 8 and 14 | West Highland White Terrier (1) | Infertile female, normal reproductive organs | [45] |
| 5 and 23 | Bernese Mountain Dog (1) | XX DSD, SRY-negative enlarged clitoris, testicle, ovotestis, uterus | [42] |
| Not identified | Miniature Schnauzer (1) | XY DSD, PMDS (Persistent Mullerian Duct Syndrome) | [46] |
| 1 and unidentified | Miniature Schnauzer (1) | XY DSD, PMDS | [47] |
| Not identified | American Staffordshire Terrier (1) | XX DSD, SRY-negative (Sex Determining Region Y) enlarged clitoris, ovotestis, | [14] |
Reciprocal translocations have rarely been diagnosed in dogs, probably due to difficulties in the recognition of one-armed autosomes. Until now, only three X/autosome mutations have been found, and this was possible because the translocation chromosome derived from the X had the abnormal morphology. The first mutation was identified in a male-to-female sex-reversed Yorkshire terrier [54]. The dog had two cell lines—a normal 78,XY and a line with X-autosome translocation. The mutation was identified using a whole X-chromosome painting probe which showed the hybridization signals on the X chromosome and unidentified autosome. Recently, two new cases of such rearrangement were observed in two female dogs with abnormalities of the genitourinary system [14]. In one of the female dogs, a pure X/autosome translocation, 78,X,t(X;2), was found, while in the second case a mosaicism of 78,X,t(X;A)/78,XX was observed. The cell line with the translocation occurred with a low incidence and it was not possible to identify the autosome involved.
It can be foreseen that the detection of canine chromosome translocations could be more efficient if a more sophisticated tool, such as multihybridization slides with a set of canine subtelomeric probes, were available, as has been recently developed for the chromosomes of pigs [55,56] and cattle [57].
5. Cytogenetic Characterization of Other Forms of DSD Cases
Cytogenetic analysis is a crucial step in classifying DSD [33,58]. Some DSD dogs may have chromosome abnormalities (sex chromosome DSD), as described above, but the majority of cases have a normal chromosome set described as XX DSD or XY DSD. Sex chromosomes are usually identified in such cases by Giemsa staining, karyotyping of R-banded chromosomes, or FISH with chromosome-specific probes (Figure 3) [32,59,60,61].
Cytogenetic analysis is also helpful for visualizing copy number variation (CNV). It has been shown that, in some dogs, CNVs in the SOX9 gene region (CFA9) are associated with XX DSD phenotype. FISH with BAC probes specific to this region was used to identify duplication or multiplication (Figure 4) [62,63,64].
Since the resolution of the hybridization signals on metaphase chromosomes is not sufficient to detect SOX9 gene triplication, interphase nuclei were examined (Figure 5) [64]. It should be mentioned that molecular techniques such as MLPA (Multiplex Ligation-dependent Probe Amplification) and a-CGH (array Comparative Genome Hybridization) have also been employed for the identification of this CNV [60,61,62].
6. Sperm Cytogenetics
The dog is a valuable large animal model in studies of human reproduction and development [65]. The segregation of sperm into two fractions, rich in, respectively, X-chromosome or Y-chromosome bearing cells, is an assisted reproductive biotechnology that has been developed for dogs. The effectiveness of this technology can be validated using the FISH technique with molecular probes specific to sex chromosomes on the segregated sperm samples. Dual color FISH has shown that the sorting of dog sperm by flow cytometry is very efficient, and the purity of the sorted samples was high at about 90% [66]. Komaki et al. [67] also used FISH to evaluate sex chromosome aneuploidy in sperm. The authors performed three color FISH with probes for chromosomes CFAX, CFAY and CFA1. Altogether, the sperms from eight dogs were analyzed and the mean frequencies of aneuploidy were: 0.016% (XX), 0.024% (YY), 0.08% (XY), and 0.176% (lack of sex chromosomes but with CFA1).
There has to date been no information on chromosome abnormalities in oocytes or embryos, despite the fact that studies of in vitro embryo production in dogs are quite advanced [68]. It can be foreseen that more such studies will be undertaken due to the increase in interest in biomedical research using induced pluripotent somatic cells [69].
7. Cancer Cytogenetics
Cancer is a genetic disease caused by gene or chromosomal mutations, classified as germline (inherited) or somatic. The somatic mutations can cause the disease or can have an effect on its development. Knowledge of the germline mutations responsible for cancer development in domestic animals, including dogs, is scarce [70]. Studies to identify mutations in cancer cells are also poorly advanced in domestic animals, though canine cancers have been considered more frequently than others [71].
Cytogenetic studies of canine cancer have a long history, with the first papers being published almost sixty years ago [72]. Early reports of chromosome abnormalities in canine tumors should be taken with caution due to the difficulties in recognizing autosomes. Since the partial international standard karyotype of the dog was agreed on in 1996, and since chromosome-specific molecular probes for FISH became available in the late 1990s, we focused this review on reports published after 1996.
Sex chromosome and autosomal aneuploidies, as well as centric fusions, are easy to identify as it was already mentioned earlier; however, the identification of small autosomes involved in such abnormalities is challenging. There are several reports showing a clonal predominance of specific aneuploidies in canine cancer cells. Analysis of G-banded chromosomes of cells derived from thyroid adenomas showed that the trisomy of chromosome 18 (CFA18) was predominant in the studied metaphase spreads [73]. Another study of in vitro cultured lymphocytes derived from the bone marrow of two dogs suffering from acute myeloid leukemia revealed two clonal aberrations: a trisomy of chromosome 1 (CFA1) and a chromosome translocation t(X;8) [74]. These aberrations were identified using G-banding. Polysomy of chromosome 13 (CFA13), caused by centric fusion between these chromosomes, was observed with an elevated frequency in cells derived from the prostate carcinomas of two dogs [75,76]. Interestingly, aberrations of this chromosome have also been observed in other dog cancers [7].
The introduction of molecular techniques into chromosome analysis was a very important step for canine cancer cytogenetics. Establishing the canine BAC library allowed the identification of BAC clones harboring 25 candidate genes for different cancers, which could be used in FISH analysis of cancer cells [77]. Researchers have searched for BAC clones in the canine genome library to use as FISH probes. Using this approach, it was shown by Breen and Modiano [78] that the well-known somatic chromosome rearrangements associated with some human cancers are also present in canine counterparts. These researchers examined the canine counterparts of three human cancers: chronic myelogenous leukemia (CML) associated with BCR and ABL fusion, sporadic Burkitt lymphoma (BL) associated with MYC-IgH fusion, and chronic lymphocytic leukemia/small lymphocytic lymphoma (CLL) associated with a hemizygous deletion harboring the RB1 gene. They found that approximately 25% of the metaphase spreads or interphase nuclei of cancer cells they studied carried similar chromosome rearrangements. This study confirmed that the dog is a valuable animal model for studies of human cancerogenesis. The colocalization of BCR-ABL was also detected by FISH in dogs suffering from chronic monocytic leukemia (CMoL) [79] and acute myeloblastic leukemia without maturation (AML-M1) [80]. Canine BAC clones and whole chromosome painting probes were used by Vozdova et al. [81], who studied canine cutaneous mast cell tumors. Among different clonal aneuploidies and structural rearrangements, the most common was trisomy of CFA11.
Complex chromosome rearrangements causing genomic imbalances (loss or gain of genetic material) can be efficiently analyzed by comparative genomic hybridization (CGH). However, the classic CGH approach requires the reliable identification of banded chromosomes. The first attempt to use CGH to analyze canine cancer cells was by Dunn et al. [82], who studied a glial tumor cell line. Unfortunately, difficulties with chromosome recognition meant it was not possible to present a detailed characterization of the imbalances. The study showed that the only abnormality observed in all metaphase spreads was CFA1 trisomy. To overcome problems with identifying chromosomes using banding techniques, a molecular version of the CGH, called array CGH (aCGH), was developed. The first canine aCGH for 87 canine BAC clones was presented by Thomas et al. [83]. Soon after, two advanced aCGH microarrays were developed. One included 1158 canine BAC clones harboring canine genome fragments distributed along all chromosomes, with an average interval of 2 Mb [84]. In the second, the distribution of the BAC clones was approximately 10 Mb [85]. These molecular tools have replaced classical cytogenetic techniques in studies of complex chromosome rearrangements in cancer cells.
8. Conclusions and Perspectives
Although great progress has been achieved in studies of the organization of the canine genome, analysis of its chromosomes remains challenging. It is not surprising that the majority of abnormalities identified so far are sex chromosome abnormalities and centric fusions, as these can be identified by conventional Giemsa staining. The identification of sex chromosomes in DSD dogs plays a very important role in elucidating the DSD background, so classical analysis of chromosome preparations should be a common diagnostic approach. It facilitates the identification of sex chromosome abnormalities (e.g., X monosomy and XXY trisomy) or of abnormal sets of sex chromosomes in leukocytes (XX/XY leukocyte chimerism). The unequivocal identification of structural chromosome rearrangements in which small autosomes are involved usually requires the use of the FISH technique with probes derived from the canine BAC library. A promising perspective is related with the application of synthetic oligonucleotide probes (oligos) designed with the use of computational tools. The oligonucleotide libraries can be a valuable source of probes specific for a given chromosome, its region or a single gene [86,87,88].
It can also be expected that, in the near future, molecular techniques will play an important role in animal clinical cytogenetics. One of such techniques is digital droplet PCR (ddPCR), which allows the determination of the number of X and Y chromosome copies and the detection of sex chromosome aneuploidies and XX/XY leukocyte chimerism in a rapid, reliable manner. Other molecular techniques such as arrayCGH, SNP-microarray, MLPA, and NGS (next generation sequencing) are already very useful in human clinical cytogenetics in detecting structural rearrangements [89,90]. Moreover, the application of BioNano technologies offers the detection of chromosomal abnormalities, CNVs and structural variants [89] with a high resolution [91]. A very recent update of the canine genome reference sequence [92] should facilitate the successful use of the sequencing technologies. Taken together, it is expected that the spectrum of traditional cytogenetic techniques used in clinical diagnosis will be replaced by advanced DNA-based technologies, which are named “cytogenomics” [93].
The development of canine cytogenetics/cytogenomics also depends on the interest of veterinarians and dog breeders, who should be aware of the importance of such testing. Since the dog is an important biomedical animal, it may be expected that new diagnostic tools will be developed to overcome the difficulties of chromosome identification.
Author Contributions
The authors contributed equally in the preparation of the manuscript. Conceptualization, I.S. and M.S.; writing, I.S. and M.S.; and visualization, I.S. and M.S. Both authors have read and agreed to the published version of the manuscript.
Funding
This study was financed by the National Science Centre, Poland, project no. 2016/23/B/NZ9/03424, and by the statutory fund of the Department of Genetics and Animal Breeding (no. 506.534.04.00), Poznan University of Life Sciences, Poland.
Institutional Review Board Statement
The Institutional Review Board statement was not required.
Data Availability Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jung, C.; Pörtl, D. How Old Are (Pet) Dog Breeds? Pet Behav. Sci. 2019, 29–37. [Google Scholar] [CrossRef]
- Breen, M. Canine Cytogenetics — from Band to Basepair. Cytogenet. Genome Res. 2008, 120, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Świtoński, M.; Reimann, N.; Bosma, A.A.; Long, S.; Bartnitzke, S.; Pieńkowska, A.; Moreno-Milan, M.M.; Fischer, P. Report on the Progress of Standardization of the G-Banded Canine (Canis Familiaris) Karyotype. Chromosome Res. 1996, 4, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Breen, M.; Bullerdiek, J.; Langford, C.F. The DAPI banded karyotype of the domestic dog (Canis familiaris) generated using chromosome-specific paint probes. Chromosome Res. 1999, 7, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Breen, M.; Hitte, C.; Lorentzen, T.D.; Thomas, R.; Cadieu, E.; Sabacan, L.; Scott, A.; Evanno, G.; Parker, H.G.; Kirkness, E.F.; et al. An Integrated 4249 Marker FISH/RH Map of the Canine Genome. BMC Genom. 2004, 5, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, R.; Smith, K.C.; Ostrander, E.A.; Galibert, F.; Breen, M. Chromosome Aberrations in Canine Multicentric Lymphomas Detected with Comparative Genomic Hybridisation and a Panel of Single Locus Probes. Br. J. Cancer 2003, 89, 1530–1537. [Google Scholar] [CrossRef] [Green Version]
- Reimann-Berg, N.; Bullerdiek, J.; Escobar, H.; Nolte, I. Chromosome analyses in dogs. Tierärztl. Prax. Ausg. K Kleintiere Heimtiere 2012, 40, 191–196. [Google Scholar] [CrossRef]
- Smith, F.W.K.; Buoen, L.C.; Weber, A.F.; Johnston, S.D.; Randolph, J.F.; Waters, D.J. X-Chromosomal Monosomy (77, XO) in a Doberman Pinscher With Gonadal Dysgenesis. J. Vet. Intern. Med. 1989, 3, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Löfstedt, R.M.; Buoen, L.C.; Weber, A.F.; Johnston, S.D.; Huntington, A.; Concannon, P.W. Prolonged Proestrus in a Bitch with X Chromosomal Monosomy (77,XO). J. Am. Vet. Med. Assoc. 1992, 200, 1104–1106. [Google Scholar]
- Switonski, M. Two Cases of Infertile Bitches with 78,XX/77,X Mosaic Karyotype: A Need for Cytogenetic Evaluation of Dogs With Reproductive Disorders. J. Hered. 2003, 94, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Mayenco-Aguirre, A.M.; Padilla, J.A.; Flores, J.M.; Daza, M.A. Canine Gonadal Dysgenesis Syndrome: A Case of Mosaicism (77,XO-78,XX). Vet. Rec. 1999, 145, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, T.; Das, P.J.; Avila, F.; Chowdhary, B.P. The Pseudoautosomal Region and Sex Chromosome Aneuploidies in Domestic Species. Sex. Dev. 2012, 6, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, T.; Chowdhary, B.P. The Eutherian Pseudoautosomal Region. Cytogenet. Genome Res. 2015, 147, 81–94. [Google Scholar] [CrossRef]
- Szczerbal, I.; Nizanski, W.; Dzimira, S.; Nowacka-Woszuk, J.; Stachecka, J.; Biezynski, J.; Ligocka, Z.; Jagodka, D.; Fabian-Kurzok, H.; Switonski, M. Chromosome Abnormalities in Dogs with Disorders of Sex Development (DSD). Anim. Reprod. Sci. 2021. submitted. [Google Scholar]
- Johnston, S.D.; Buoen, L.C.; Weber, A.F.; Madl, J.E. X Trisomy in an Airedale Bitch with Ovarian Dysplasia and Primary Anestrus. Theriogenology 1985, 24, 597–607. [Google Scholar] [CrossRef]
- Switonski, M.; Godynicki, S.; Jackowiak, H.; Piekowska, A.; Turczuk-Bierla, I.; Szymas, J.; Golinski, P.; Bereszynski, A. Brief Communication. X Trisomy in an Infertile Bitch: Cytogenetic, Anatomic, and Histologic Studies. J. Hered. 2000, 91, 149–150. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt, B.; Paulino, F.O.; Sauza, L.M.; Gomes, H.F. Infertility Related to X-Trisomy in a Labrador Retriever Bitch. J. Israeli Vet. Med. Assoc. 2003, 58, 123–124. [Google Scholar]
- O’Connor, C.L.; Schweizer, C.; Gradil, C.; Schlafer, D.; Lopate, C.; Prociuk, U.; Meyers-Wallen, V.N.; Casal, M.L. Trisomy-X with Estrous Cycle Anomalies in Two Female Dogs. Theriogenology 2011, 76, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Clough, E.; Pyle, R.L.; Hare, W.C.D.; Kelly, D.F.; Patterson, D.F. An XXY Sex-Chromosome Constitution in a Dog with Testicular Hypoplasia and Congenital Heart Disease. Cytogenet. Genome Res. 1970, 9, 71–77. [Google Scholar] [CrossRef]
- Marshall, L.S.; Oehlert, M.L.; Haskins, M.E.; Selden, J.R.; Patterson, D.F. Persistent Müllerian Duct Syndrome in Miniature Schnauzers. J. Am. Vet. Med. Assoc. 1982, 181, 798–801. [Google Scholar]
- Goldschmidt, B.; El-Jaick, K.B.; Souza, L.M.; Carvalho, E.C.Q.; Moura, V.L.S.; Benevides Filho, I.M. Cryptorchidism Associated with 78,XY/79,XXY Mosaicism in Dog. Israel J. Vet. Med. 2001, 56, 56e8. [Google Scholar]
- Reimann-Berg, N.; Escobar, H.M.; Nolte, I.; Bullerdiek, J. Testicular Tumor in an XXY Dog. Cancer Genet. Cytogenet. 2008, 183, 114–116. [Google Scholar] [CrossRef]
- Bosu, W.T.; Chick, B.F.; Basrur, P.K. Clinical, Pathologic and Cytogenetic Observations on Two Intersex Dogs. Cornell Vet. 1978, 68, 375–390. [Google Scholar]
- Nie, G.J.; Johnston, S.D.; Hayden, D.W.; Buoen, L.C.; Stephens, M. Theriogenology Question of the Month. Azoospermia Associated with 79,XXY Chromosome Complement (Canine Klinefelter’s Syndrome). J. Am. Vet. Med. Assoc. 1998, 212, 1545–1547. [Google Scholar]
- Biason-Lauber, A. Control of Sex Development. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 163–186. [Google Scholar] [CrossRef] [Green Version]
- Esteves, A.; Bage, R.; Payan-Carreira, R. Freemartinism in cattle. In Ruminants: Anatomy, Behavior and Diseases; Mendes, R.E., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2012; pp. 99–120. [Google Scholar]
- Szczerbal, I.; Nowacka-Woszuk, J.; Dzimira, S.; Matuszczyk, A.; Iskrzak, P.; Switonski, M. Elevated Incidence of Freemartinism in Pigs Detected by Droplet Digital PCR and Cytogenetic Techniques. Livest. Sci. 2019, 219, 52–56. [Google Scholar] [CrossRef]
- Beccaglia, M.; Ronchese, M.; Grieco, V.; Parma, P.; Luvoni, G.C. XX/XY Chimaerism/Mosaicism in a Phenotypically Female Wirehaired Pointing Griffon Dog. In Proceedings of the 7th International Symposium on Canine and Feline Reproduction, Whistler, BC, Canada, 26–29 July 2012. [Google Scholar]
- Genero, E.R.; Moreno-Millán, M.; Ocaña-Quero, J.M. XX/XY Chromosome Chimaerism in an Intersex Dog. Vet. Rec. 1998, 142, 340. [Google Scholar] [CrossRef]
- Kuiper, H.; Distl, O. Intersexuality in dogs: Causes and genetics. DTW Dtsch. Tierarztl. Wochenschr. 2004, 111, 251–258. [Google Scholar]
- Szczerbal, I.; Nowacka-Woszuk, J.; Nizanski, W.; Salamon, S.; Ochota, M.; Dzimira, S.; Atamaniuk, W.; Switonski, M. A Case of Leucocyte Chimerism (78,XX/78,XY) in a Dog with a Disorder of Sexual Development. Reprod. Domest. Anim. 2014, 49, e31–e34. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Nowacka-Woszuk, J.; Nizanski, W.; Dzimira, S.; Ligocka, Z.; Jastrzebska, A.; Kabala, B.; Biernacik, M.; Przadka, P.; Switonski, M. Disorders of Sex Development Are an Emerging Problem in French Bulldogs: A Description of Six New Cases and a Review of the Literature. Sex. Dev. 2019, 13, 205–211. [Google Scholar] [CrossRef]
- Poth, T.; Breuer, W.; Walter, B.; Hecht, W.; Hermanns, W. Disorders of Sex Development in the Dog—Adoption of a New Nomenclature and Reclassification of Reported Cases. Anim. Reprod. Sci. 2010, 121, 197–207. [Google Scholar] [CrossRef]
- Hare, W.C. Intersexuality in the Dog. Can. Vet. J. Rev. Veterinaire Can. 1976, 17, 7–15. [Google Scholar]
- Weaver, A.D.; Harvey, M.J.; Munro, C.D.; Rogerson, P.; McDonald, M. Phenotypic Intersex (Female Pseudohermaphroditism) in a Dachshund Dog. Vet. Rec. 1979, 105, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.E.; Long, S.E.; Gibbs, C. Disorders of Urination Associated with Canine Intersexuality. J. Small Anim. Pract. 1983, 24, 475–487. [Google Scholar] [CrossRef]
- Johnston, S.D. Premature Gonadal Failure in Female Dogs and Cats. J. Reprod. Fertil. Suppl. 1989, 39, 65–72. [Google Scholar] [PubMed]
- Chaffaux, S.; Cribiu, E. Clinical, Histological and Cytogenetic Observations on Nine Intersex Dogs. Genet. Sel. Evol. 1991, 23, S81. [Google Scholar] [CrossRef]
- Meyers-Wallen, V.N. Inherited abnormalities of sexual development in dogs and cats. In Recent Advances in Small Animal Reproduction; Concannon, P.W., England, G., Ver-stegen, J., Eds.; International Veterinary Information Service, USA, 2001; Available online: https://www.ivis.org/library/recent-advances-small-animal-reproduction/inherited-abnormalities-of-sexual-development (accessed on 10 January 2021).
- Sumner, S.M.; Case, J.B.; Regier, P.J.; Oliveira, L.; Abbott, J.R. Laparoscopic Gonadectomy in a Dog with 78,XX/78,XY Chimerism and Underdeveloped Reproductive Organs. J. Am. Vet. Med. Assoc. 2021, 258, 80–84. [Google Scholar] [CrossRef]
- Szczerbal, I.; Nowacka-Woszuk, J.; Albarella, S.; Switonski, M. Technical Note: Droplet Digital PCR as a New Molecular Method for a Simple and Reliable Diagnosis of Freemartinism in Cattle. J. Dairy Sci. 2019, 102, 10100–10104. [Google Scholar] [CrossRef] [PubMed]
- Switonski, M.; Szczerbal, I.; Nizanski, W.; Kociucka, B.; Bartz, M.; Dzimira, S.; Mikolajewska, N. Robertsonian Translocation in a Sex Reversal Dog (XX, SRY Negative) May Indicate That the Causative Mutation for This Intersexuality Syndrome Resides on Canine Chromosome 23 (CFA23). Sex. Dev. 2011, 5, 141–146. [Google Scholar] [CrossRef]
- Stone, D.M.; Mickelsen, W.D.; Jacky, P.B.; Prieur, D.J. A Novel Robertsonian Translocation in a Family of Walker Hounds. Genome 1991, 34, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Switonski, M.; Slota, E.; Pietrzak, A.; Klukowska, J. Chimerism 78,XX/77,XX, Rb in a bitch revealed by chromosome and microsatellite studies. Vet. Med. Czech. 2000, 45, 296–298. [Google Scholar]
- Switonski, M.; Szczerbal, I.; Skorczyk, A.; Yang, F.; Antosik, P. Robertsonian Translocation (8;14) in an Infertile Bitch (Canis Familiaris). J. Appl. Genet. 2003, 44, 525–527. [Google Scholar] [PubMed]
- Dzimira, S.; Wydooghe, E.; Van Soom, A.; Van Brantegem, L.; Nowacka-Woszuk, J.; Szczerbal, I.; Switonski, M. Sertoli Cell Tumour and Uterine Leiomyoma in Miniature Schnauzer Dogs with Persistent Müllerian Duct Syndrome Caused by Mutation in the AMHR2 Gene. J. Comp. Pathol. 2018, 161, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, D.M.; Armada, J.L.A.; Penedo, D.M.; Tannouz, V.G.S.; Meyers-Wallen, V.N. Persistent Mullerian Duct Syndrome in a Brazilian Miniature Schnauzer Dog. An. Acad. Bras. Ciênc. 2019, 91, e20180752. [Google Scholar] [CrossRef]
- Shive, R.J.; Hare, W.C.D.; Patterson, D.F. Chromosome Studies in Dogs with Congenital Cardiac Defects. Cytogenet. Genome Res. 1965, 4, 340–348. [Google Scholar] [CrossRef]
- Hare, W.C.; Wilkinson, J.S.; McFeely, R.A.; Riser, W.H. Bone Chondroplasia and a Chromosome Abnormality in the Same Dog. Am. J. Vet. Res. 1967, 28, 583–587. [Google Scholar]
- Ma, N.S.F.; Gilmore, C.E. Chromosomal Abnormality in a Phenotypically and Clinically Normal Dog. Cytogenet. Genome Res. 1971, 10, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Larsen, R.E.; Dias, E.; Cervenka, J. Centric Fusion of Autosomal Chromosomes in a Bitch and Offspring. Am. J. Vet. Res. 1978, 39, 861–864. [Google Scholar]
- Larsen, R.E.; Dias, E.; Flores, G.; Selden, J.R. Breeding Studies Reveal Segregation of a Canine Robertsonian Translocation along Mendelian Proportions. Cytogenet. Cell Genet. 1979, 24, 95–101. [Google Scholar] [CrossRef]
- Mayr, B.; Krutzler, J.; Schleger, W.; Auer, H. A New Type of Robertsonian Translocation in the Domestic Dog. J. Hered. 1986, 77, 127. [Google Scholar] [CrossRef]
- Schelling, C.; Pieńkowska, A.; Arnold, S.; Hauser, B.; Switoński, M. A Male to Female Sex-Reversed Dog with a Reciprocal Translocation. J. Reprod. Fertil. Suppl. 2001, 57, 435–438. [Google Scholar] [PubMed]
- O’Connor, R.E.; Fonseka, G.; Frodsham, R.; Archibald, A.L.; Lawrie, M.; Walling, G.A.; Griffin, D.K. Isolation of Subtelomeric Sequences of Porcine Chromosomes for Translocation Screening Reveals Errors in the Pig Genome Assembly. Anim. Genet. 2017, 48, 395–403. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.E.; Kiazim, L.G.; Rathje, C.C.; Jennings, R.L.; Griffin, D.K. Rapid Multi-Hybridisation FISH Screening for Balanced Porcine Reciprocal Translocations Suggests a Much Higher Abnormality Rate Than Previously Appreciated. Cells 2021, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.L.; Griffin, D.K.; O’Connor, R.E. A New Approach for Accurate Detection of Chromosome Rearrangements That Affect Fertility in Cattle. Animals 2020, 10, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers-Wallen, V.N. Gonadal and Sex Differentiation Abnormalities of Dogs and Cats. Sex. Dev. 2012, 6, 46–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Switonski, M.; Payan-Carreira, R.; Bartz, M.; Nowacka-Woszuk, J.; Szczerbal, I.; Colaço, B.; Pires, M.A.; Ochota, M.; Nizanski, W. Hypospadias in a Male (78,XY.; SRY-Positive) Dog and Sex Reversal Female (78,XX.; SRY-Negative) Dogs: Clinical, Histological and Genetic Studies. Sex. Dev. 2012, 6, 128–134. [Google Scholar] [CrossRef]
- Rossi, E.; Radi, O.; De Lorenzi, L.; Vetro, A.; Groppetti, D.; Bigliardi, E.; Luvoni, G.C.; Rota, A.; Camerino, G.; Zuffardi, O.; et al. Sox9 Duplications Are a Relevant Cause of Sry-Negative XX Sex Reversal Dogs. PLoS ONE 2014, 9, e101244. [Google Scholar] [CrossRef] [Green Version]
- Albarella, S.; Lorenzi, L.D.; Rossi, E.; Prisco, F.; Riccardi, M.G.; Restucci, B.; Ciotola, F.; Parma, P. Analysis of XX SRY-Negative Sex Reversal Dogs. Animals 2020, 10, 1667. [Google Scholar] [CrossRef]
- Marcinkowska-Swojak, M.; Szczerbal, I.; Pausch, H.; Nowacka-Woszuk, J.; Flisikowski, K.; Dzimira, S.; Nizanski, W.; Payan-Carreira, R.; Fries, R.; Kozlowski, P.; et al. Copy Number Variation in the Region Harboring SOX9 Gene in Dogs with Testicular/Ovotesticular Disorder of Sex Development (78,XX.; SRY-Negative). Sci. Rep. 2015, 5, 14696. [Google Scholar] [CrossRef] [Green Version]
- Szczerbal, I.; Nowacka-Woszuk, J.; Dzimira, S.; Atamaniuk, W.; Nizanski, W.; Switonski, M. A Rare Case of Testicular Disorder of Sex Development in a Dog (78,XX.; SRY-Negative) with Male External Genitalia and Detection of Copy Number Variation in the Region Upstream of the SOX9 Gene. Sex. Dev. 2016, 10, 74–78. [Google Scholar] [CrossRef]
- Nowacka-Woszuk, J.; Szczerbal, I.; Stachowiak, M.; Szydlowski, M.; Nizanski, W.; Dzimira, S.; Maslak, A.; Payan-Carreira, R.; Wydooghe, E.; Nowak, T.; et al. Association between Polymorphisms in the SOX9 Region and Canine Disorder of Sex Development (78,XX.; SRY-Negative) Revisited in a Multibreed Case-Control Study. PLoS ONE 2019, 14, e0218565. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J. Spotlight on reproduction in domestic dogs as a model for human reproduction. In Animals Model and Human Reproduction; Schatten, H., Constantinescu, G.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 247–358. [Google Scholar]
- Oi, M.; Yamada, K.; Hayakawa, H.; Suzuki, H. Sexing of Dog Sperm by Fluorescence In Situ Hybridization. J. Reprod. Dev. 2012. [Google Scholar] [CrossRef] [Green Version]
- Komaki, H.; Oi, M.; Suzuki, H. Detection of Sex Chromosome Aneuploidy in Dog Spermatozoa by Triple Color Fluorescence in Situ Hybridization. Theriogenology 2014, 82, 652–656. [Google Scholar] [CrossRef]
- Nagashima, J.B.; Travis, A.J.; Songsasen, N. The Domestic Dog Embryo: In Vitro Fertilization, Culture, and Transfer. In Comparative Embryo Culture; Herrick, J.R., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 2006, pp. 247–267. ISBN 978-1-4939-9565-3. [Google Scholar]
- Hyttel, P.; Pessôa, L.V.d.F.; Secher, J.B.-M.; Dittlau, K.S.; Freude, K.; Hall, V.J.; Fair, T.; Assey, R.J.; Laurincik, J.; Callesen, H.; et al. Oocytes, Embryos and Pluripotent Stem Cells from a Biomedical Perspective. Anim. Reprod. 2019, 16, 508–523. [Google Scholar] [CrossRef]
- Flisikowski, K.; Flisikowska, T.; Sikorska, A.; Perkowska, A.; Kind, A.; Schnieke, A.; Switonski, M. Germline Gene Polymorphisms Predisposing Domestic Mammals to Carcinogenesis: Gene Polymorphisms Predisposing to Carcinogenesis. Vet. Comp. Oncol. 2017, 15, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Ostrander, E.A.; Dreger, D.L.; Evans, J.M. Canine Cancer Genomics: Lessons for Canine and Human Health. Annu. Rev. Anim. Biosci. 2019, 7, 449–472. [Google Scholar] [CrossRef] [PubMed]
- Makino, S. Some epidemiologic aspects of venereal tumors of dogs as revealed by chromosome and DNA studies. Ann. N. Y. Acad. Sci. 2006, 108, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- Reimann, N.; Nolte, I.; Bonk, U.; Werner, M.; Bullerdiek, J.; Bartnitzke, S. Trisomy 18 in a Canine Thyroid Adenoma. Cancer Genet. Cytogenet. 1996, 90, 154–156. [Google Scholar] [CrossRef]
- Reimann, N.; Bartnitzke, S.; Bullerdiek, J.; Mischke, R.; Nolte, I. Trisomy 1 in a Canine Acute Leukemia Indicating the Pathogenetic Importance of Polysomy 1 in Leukemias of the Dog. Cancer Genet. Cytogenet. 1998, 101, 49–52. [Google Scholar] [CrossRef]
- Winkler, S.; Reimann-Berg, N.; Escobar, H.M.; Loeschke, S.; Eberle, N.; Höinghaus, R.; Nolte, I.; Bullerdiek, J. Polysomy 13 in a Canine Prostate Carcinoma Underlining Its Significance in the Development of Prostate Cancer. Cancer Genet. Cytogenet. 2006, 169, 154–158. [Google Scholar] [CrossRef]
- Reimann-Berg, N.; Willenbrock, S.; Murua Escobar, H.; Eberle, N.; Gerhauser, I.; Mischke, R.; Bullerdiek, J.; Nolte, I. Two New Cases of Polysomy 13 in Canine Prostate Cancer. Cytogenet. Genome Res. 2011, 132, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Bridge, W.; Benke, K.; Breen, M. Isolation and Chromosomal Assignment of Canine Genomic BAC Clones Representing 25 Cancer-Related Genes. Cytogenet. Genome Res. 2003, 102, 249–253. [Google Scholar] [CrossRef]
- Breen, M.; Modiano, J.F. Evolutionarily Conserved Cytogenetic Changes in Hematological Malignancies of Dogs and Humans—Man and His Best Friend Share More than Companionship. Chromosome Res. 2008, 16, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Cruz Cardona, J.A.; Milner, R.; Alleman, A.R.; Williams, C.; Vernau, W.; Breen, M.; Tompkins, M. BCR-ABL Translocation in a Dog with Chronic Monocytic Leukemia: BCR-ABL Translocation in Canine CMoL. Vet. Clin. Pathol. 2011, 40, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.F.; Culver, S.; Behling-Kelly, E.; Breen, M.; Friedrichs, K.R. Acute Myeloblastic Leukemia with Associated BCR-ABL Translocation in a Dog. Vet. Clin. Pathol. 2012, 41, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Vozdova, M.; Kubickova, S.; Cernohorska, H.; Fröhlich, J.; Fictum, P.; Rubes, J. Structural and Copy Number Chromosome Abnormalities in Canine Cutaneous Mast Cell Tumours. J. Appl. Genet. 2019, 60, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.A.; Thomas, R.; Binns, M.M.; Breen, M. Comparative Genomic Hybridization (CGH) in Dogs—Application to the Study of a Canine Glial Tumour Cell Line. Vet. J. 2000, 160, 77–82. [Google Scholar] [CrossRef]
- Thomas, R.; Fiegler, H.; Ostrander, E.A.; Galibert, F.; Carter, N.P.; Breen, M. A Canine Cancer-Gene Microarray for CGH Analysis of Tumors. Cytogenet. Genome Res. 2003, 102, 254–260. [Google Scholar] [CrossRef]
- Thomas, R. Construction of a 2-Mb Resolution BAC Microarray for CGH Analysis of Canine Tumors. Genome Res. 2005, 15, 1831–1837. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Duke, S.E.; Bloom, S.K.; Breen, T.E.; Young, A.C.; Feiste, E.; Seiser, E.L.; Tsai, P.-C.; Langford, C.F.; Ellis, P.; et al. A Cytogenetically Characterized, Genome-Anchored 10-Mb BAC Set and CGH Array for the Domestic Dog. J. Hered. 2007, 98, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, D.; Voith von Voithenberg, L.; Kaigala, G.V. Fluorescence in Situ Hybridization (FISH): History, Limitations and What to Expect from Micro-Scale FISH? Micro Nano Eng. 2018, 1, 15–24. [Google Scholar] [CrossRef]
- Beliveau, B.J.; Kishi, J.Y.; Nir, G.; Sasaki, H.M.; Saka, S.K.; Nguyen, S.C.; Wu, C.; Yin, P. OligoMiner Provides a Rapid, Flexible Environment for the Design of Genome-Scale Oligonucleotide in Situ Hybridization Probes. Proc. Natl. Acad. Sci. USA 2018, 115, E2183–E2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beliveau, B.J.; Joyce, E.F.; Apostolopoulos, N.; Yilmaz, F.; Fonseka, C.Y.; McCole, R.B.; Chang, Y.; Li, J.B.; Senaratne, T.N.; Williams, B.R.; et al. Versatile Design and Synthesis Platform for Visualizing Genomes with Oligopaint FISH Probes. Proc. Natl. Acad. Sci. USA 2012, 109, 21301–21306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Wang, L.; Wang, H.; Ma, M.; Wang, S.; Liu, Z.; Xu, G.; Zhang, J.; Cram, D.S.; Yao, Y. The Clinical Utility of Next-Generation Sequencing for Identifying Chromosome Disease Syndromes in Human Embryos. Reprod. Biomed. Online 2015, 31, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Liang, F.; Cheng, D.; Zhang, Z.; Yu, G.; Zha, J.; Wang, Y.; Xia, Q.; Yuan, D.; Tan, Y.; et al. Location of Balanced Chromosome-Translocation Breakpoints by Long-Read Sequencing on the Oxford Nanopore Platform. Front. Genet. 2020, 10, 1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahajpal, N.S.; Barseghyan, H.; Kolhe, R.; Hastie, A.; Chaubey, A. Optical Genome Mapping as a Next-Generation Cytogenomic Tool for Detection of Structural and Copy Number Variations for Prenatal Genomic Analyses. Genes 2021, 12, 398. [Google Scholar] [CrossRef]
- Wang, C.; Wallerman, O.; Arendt, M.-L.; Sundström, E.; Karlsson, Å.; Nordin, J.; Mäkeläinen, S.; Pielberg, G.R.; Hanson, J.; Ohlsson, Å.; et al. A Novel Canine Reference Genome Resolves Genomic Architecture and Uncovers Transcript Complexity. Commun. Biol. 2021, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- Hochstenbach, R.; Liehr, T.; Hastings, R.J. Chromosomes in the Genomic Age. Preserving Cytogenomic Competence of Diagnostic Genome Laboratories. Eur. J. Hum. Genet. 2020. [Google Scholar] [CrossRef]
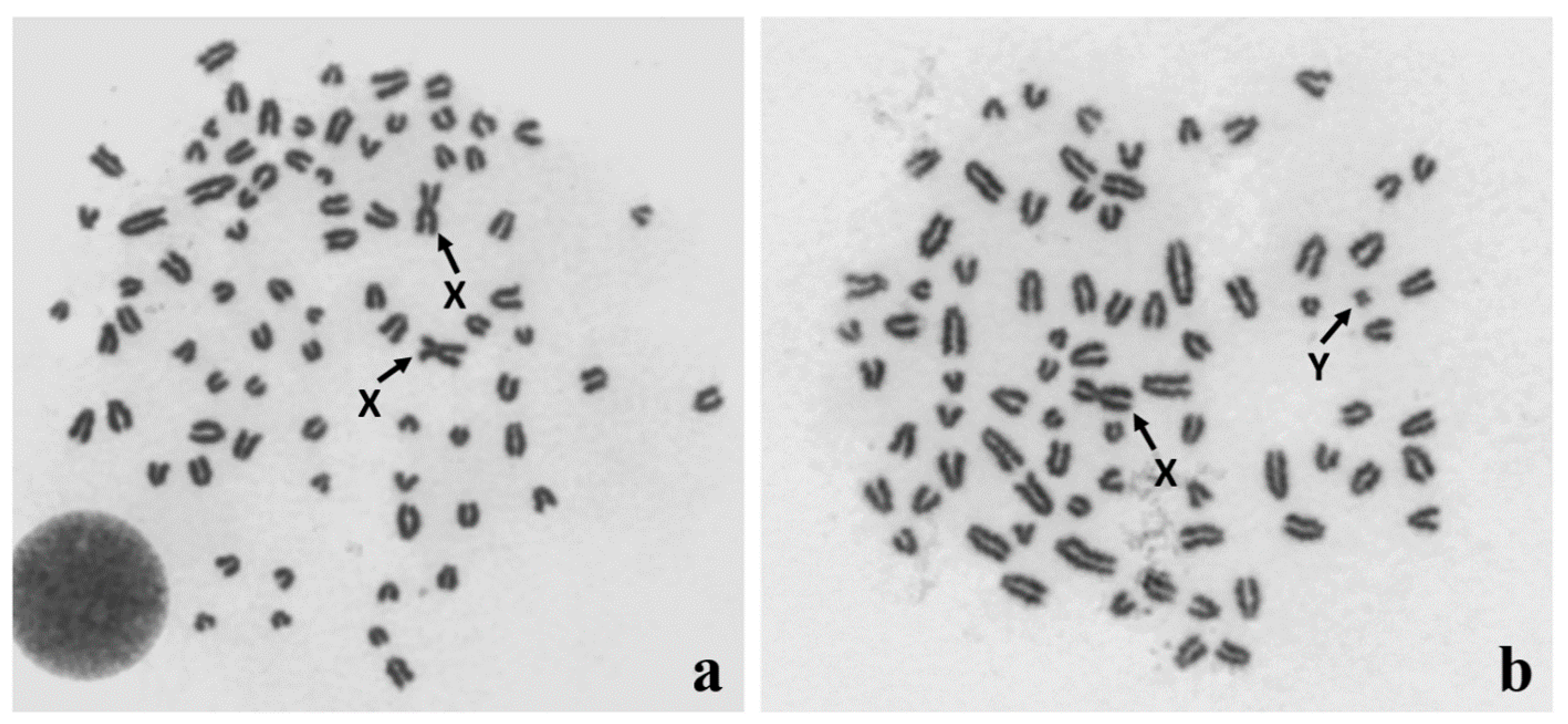
Figure 1.
Identification of (a) 78,XX and (b) 78,XY Giemsa-stained metaphase spreads from in vitro cultured leukocytes obtained from a DSD (disorder of sex development) dog. Sex chromosomes are indicated with arrows.
Figure 1.
Identification of (a) 78,XX and (b) 78,XY Giemsa-stained metaphase spreads from in vitro cultured leukocytes obtained from a DSD (disorder of sex development) dog. Sex chromosomes are indicated with arrows.

Figure 2.
Robertsonian translocation, 77,XX, rob(5;23) in an infertile DSD female dog: (a) Q-banded metaphase spread with three biarmed chromosomes; (b) the same metaphase spread after fluorescence in situ hybridization (FISH) using whole X chromosome painting probe to facilitate recognition of X chromosomes from the fused chromosome (rob). The autosomes involved in the translocation were identified by FISH with locus-specific probes (for details, see [42]).
Figure 2.
Robertsonian translocation, 77,XX, rob(5;23) in an infertile DSD female dog: (a) Q-banded metaphase spread with three biarmed chromosomes; (b) the same metaphase spread after fluorescence in situ hybridization (FISH) using whole X chromosome painting probe to facilitate recognition of X chromosomes from the fused chromosome (rob). The autosomes involved in the translocation were identified by FISH with locus-specific probes (for details, see [42]).
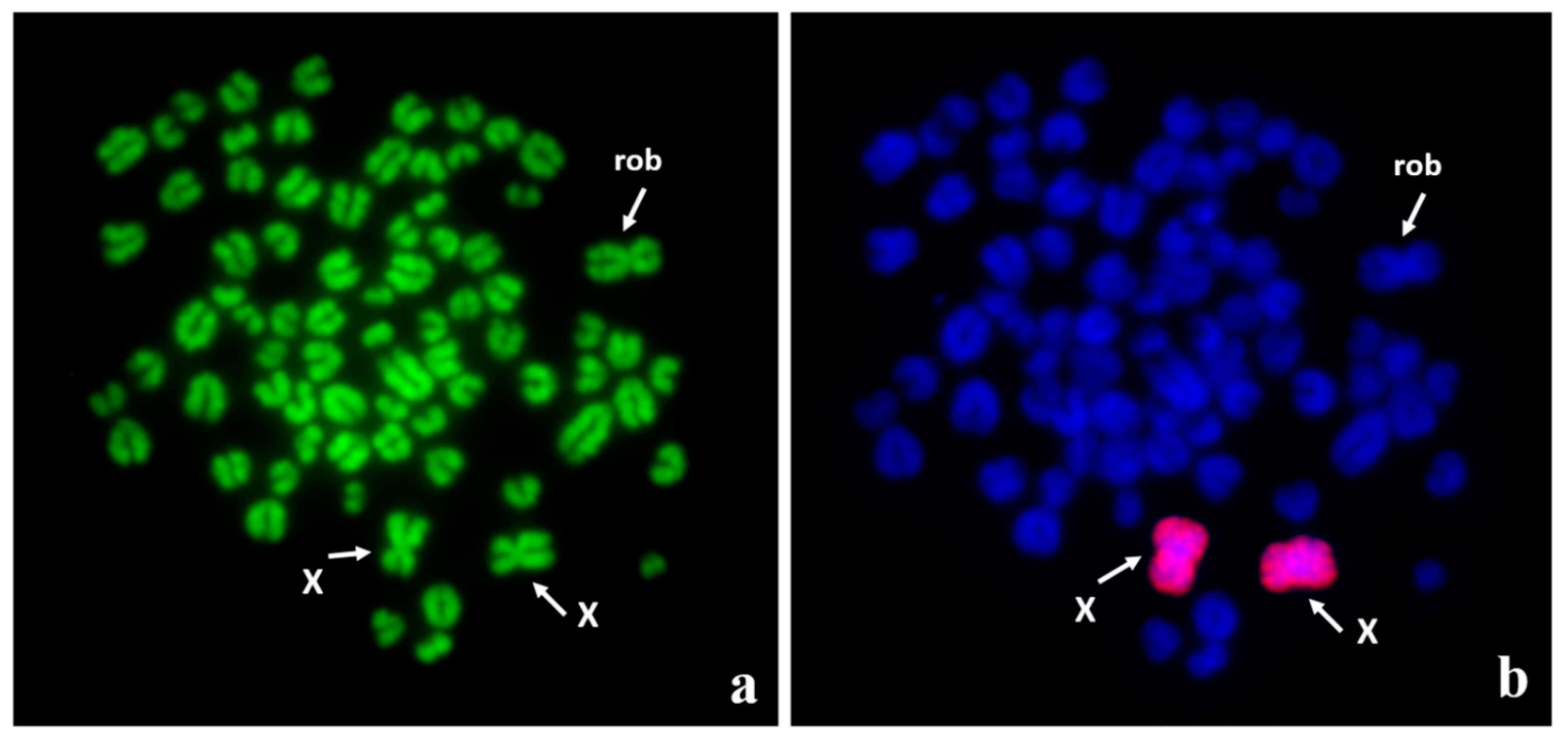
Figure 3.
Identification of sex chromosomes by FISH with the use of painting probes (X: red) and (Y: green): (a) 78,XX, (b) 78,XY, with visible signals in pseudoautosomal region (PAR).
Figure 3.
Identification of sex chromosomes by FISH with the use of painting probes (X: red) and (Y: green): (a) 78,XX, (b) 78,XY, with visible signals in pseudoautosomal region (PAR).

Figure 4.
Identification of the copy number variation (CNV) in the region of SOX9, located on CFA9. Two BAC (Bacterial Artificial Chromosome) clones were used: the green signals are specific to the SOX9 gene and the red signals are specific to the upstream CNV. The red probe also presents homology to CFA18. (a) The two large red signals on CFA9 indicate multiplication of the CNV region. (b) Another example of the variation - the single large, red signal is visible on one CFA9 chromosome, only.
Figure 4.
Identification of the copy number variation (CNV) in the region of SOX9, located on CFA9. Two BAC (Bacterial Artificial Chromosome) clones were used: the green signals are specific to the SOX9 gene and the red signals are specific to the upstream CNV. The red probe also presents homology to CFA18. (a) The two large red signals on CFA9 indicate multiplication of the CNV region. (b) Another example of the variation - the single large, red signal is visible on one CFA9 chromosome, only.
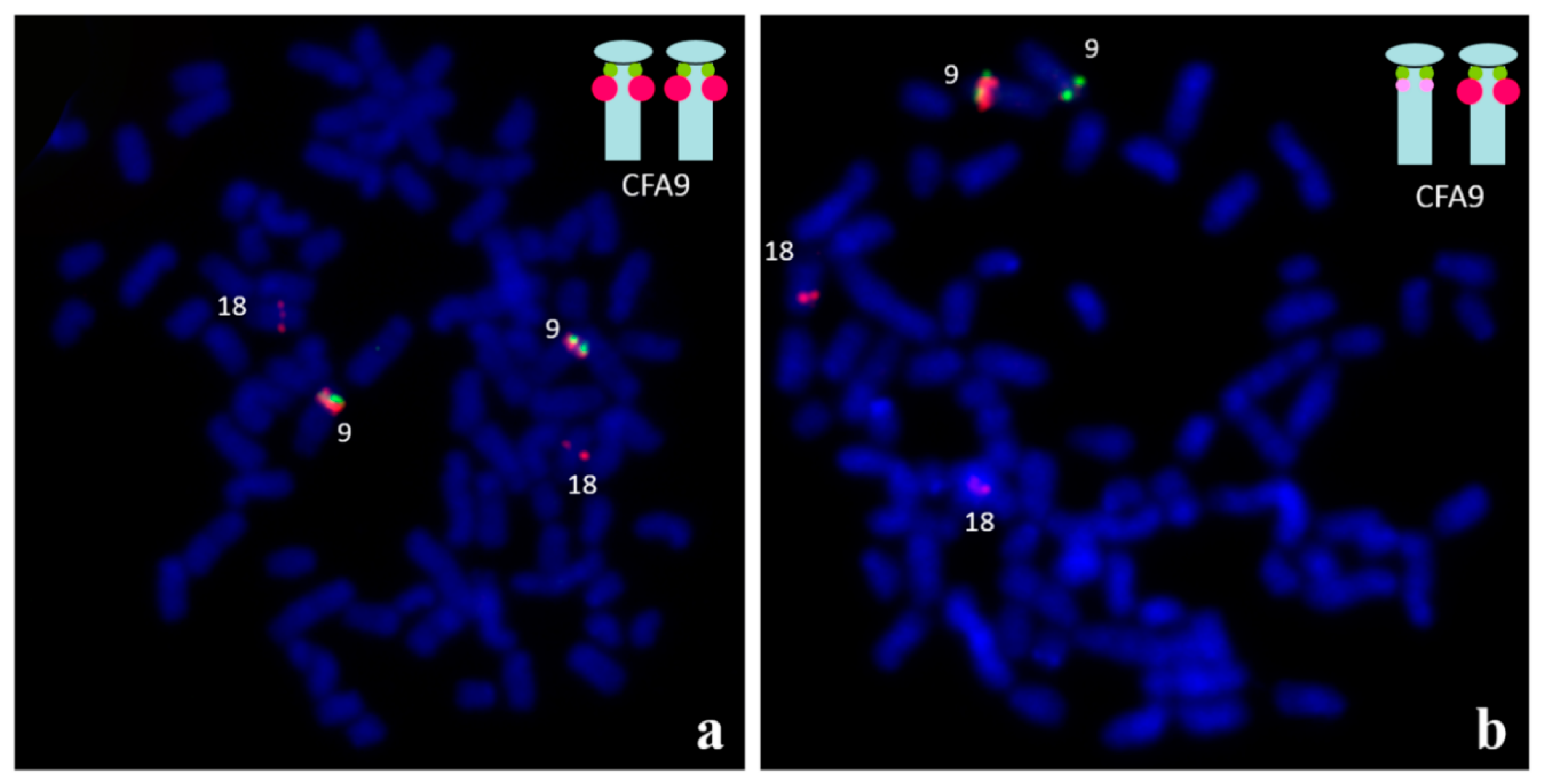
Figure 5.
FISH for identification of three copies of SOX9 gene (green signals) in two interphase nuclei (a,b). Moreover, multiple copies at CNV region upstream SOX9, as well as homologous region of CFA18 (red signals), are visible. For details, see [64].
Figure 5.
FISH for identification of three copies of SOX9 gene (green signals) in two interphase nuclei (a,b). Moreover, multiple copies at CNV region upstream SOX9, as well as homologous region of CFA18 (red signals), are visible. For details, see [64].
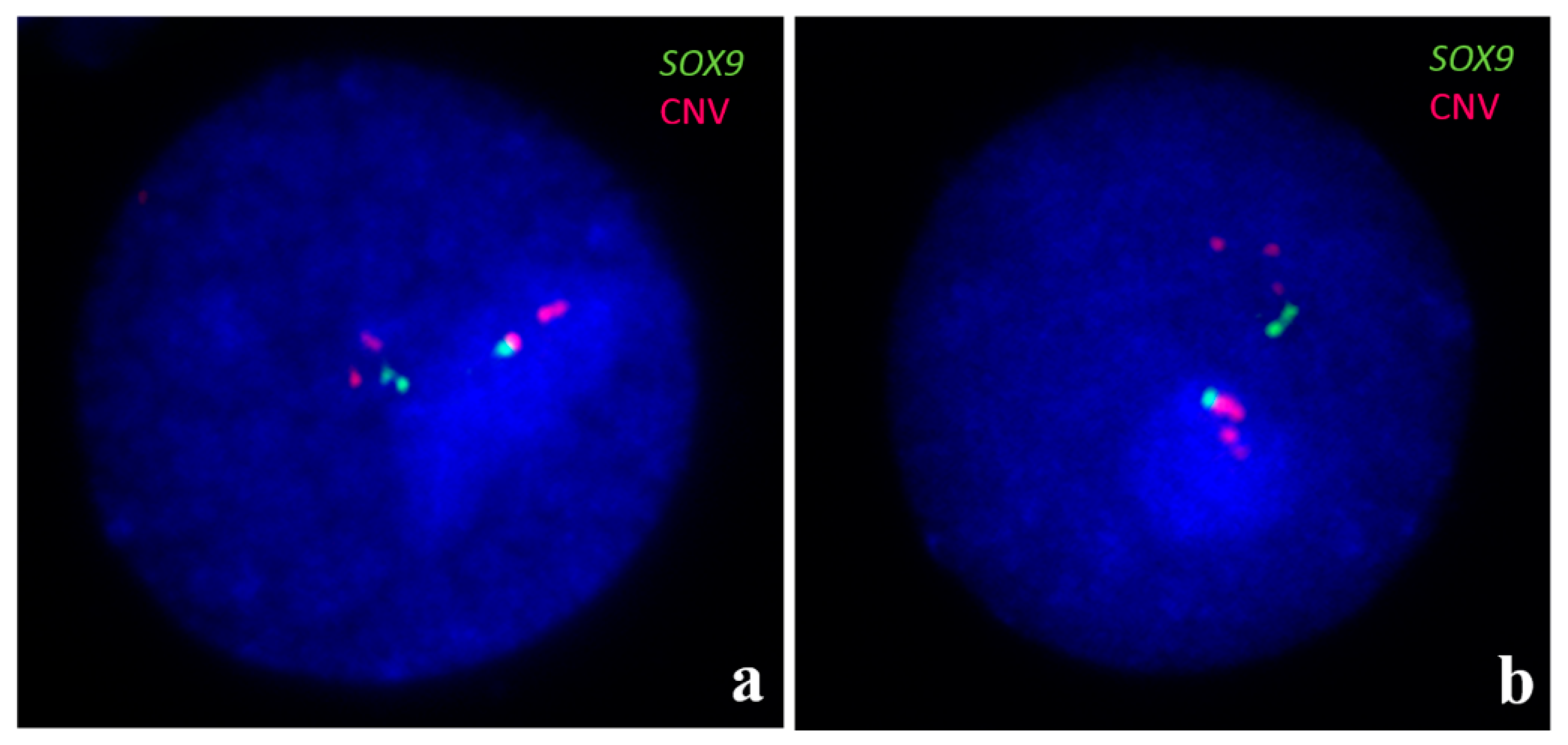
Table 1.
Cases of X monosomy reported in dogs.
| Karyotype | No. of Cells Analyzed | Breed | Characteristic Feature of Phenotype | Reference |
|---|---|---|---|---|
| 77,X | Lack of information | Doberman Pinscher | Small stature, excessive skin in the ventrum of the neck, no signs of estrus, small ovaries consisting primarily of interstitial-type cells and solid epithelial cords | [8] |
| 77,X | 60 | Miniature American Eskimo | Juvenile appearance, signs of proestrus, small and fibrous ovaries, no evidence of ovarian follicle development or corpora lutea | [9] |
| 77,X[95%]/78,XX[5%] | 40 | Toy Poodle | Abnormal estrus cycle and apparently persistent follicles, gonadal dysgenesis | [11] |
| 77,X[5%]/78,XX[95%] | 220 | Munsterlander | Infertility, vertical septum in vagina | [10] |
| 77,X[6%]/78,XX[94%] | 473 | Bearded Collie | Infertility, irregular and poorly manifested estrus cycles | [10] |
Table 2.
Cases of X trisomy reported in dogs.
| Karyotype | Breed | Characteristic Feature of Phenotype | Reference |
|---|---|---|---|
| 79,XXX | Airedale Terrier | Primary anestrus, ovaries with solid epithelial cords and large masses of interstitial cells, lack of follicles and corpora lutea | [15] |
| 79,XXX | Mixed breed | Infertility, normal reproductive organs, ovaries with primary follicles and corpora lutea, dental anomalies, abnormal behavior (lack of barking and fearfulness) | [16] |
| 79,XXX | Labrador Retriever | Primary anestrus, chronic dermatitis, abnormal behavior (coprophagy) | [17] |
| 79,XXX | Silky Terrier | Infertility, abnormal estrous cycles, hypoplastic ovaries, absence of normal follicular structures, shy and timid behavior | [18] |
| 79,XXX | Labrador Retriever | Infertility, abnormal estrous cycles, hypoplastic ovaries, absence of normal follicular structures | [18] |
| 79,XXX/78,XX | Boston Terrier | Estrus symptoms occurred once, ovary with corpora lutea | [14] |
Table 3.
Cases of the XXY complement reported in dogs.
| Karyotype | Breed | Characteristic Feature of Phenotype | Reference |
|---|---|---|---|
| 79,XXY | German Shorthair Pointer | Testicular hypoplasia, lack of spermatogenesis, ventricular septal defect, congenital heart abnormalities | [19] |
| 79,XXY | Great Dane | Female external and internal genitalia, structure reminiscent of a vestigial scrotal sac | [23] |
| 79,XXY | Norwich Terrier | Testicular dysgenesis, azoospermia | [24] |
| 79,XXY | West Highland White Terrier | High stature, rugae of the dermis and hypodermis, low level of testosterone, Sertoli cell tumor | [22] |
| 79,XXY/78,XY | Miniature Schnauzer | Alopecia, gynecomastia, bilateral cryptorchidism, Sertoli cell tumor | [20] |
| 79,XXY[18%]/78,XY[82%] | Poodle | Bilateral cryptorchidism, testes with vacuolation of the seminal cells and small nests of Leydig cells, total absence of sperm cells | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szczerbal, I.; Switonski, M. Clinical Cytogenetics of the Dog: A Review. Animals 2021, 11, 947. https://doi.org/10.3390/ani11040947
AMA Style
Szczerbal I, Switonski M. Clinical Cytogenetics of the Dog: A Review. Animals. 2021; 11(4):947. https://doi.org/10.3390/ani11040947
Chicago/Turabian StyleSzczerbal, Izabela, and Marek Switonski. 2021. "Clinical Cytogenetics of the Dog: A Review" Animals 11, no. 4: 947. https://doi.org/10.3390/ani11040947
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.