1. Introduction
In a fish farming context, fish are susceptible to a wide range of pathogens, since seawater is a good growing media for many bacteria and virus that can ruin an entire fish stock. Once present in water, pathogens are in direct contact with the first lines of defense of fish and can easily spread, particularly if the animals are not well-nourished and prepared [
1,
2]. As such, health maintenance is of utmost importance in modern fish farming, and establishing strategies to improve fish immune status and welfare is essential.
Recently, there has been an effort to adopt the use of supplements that can boost fish immune status (e.g., yeast extracts, probiotics, prebiotics, and amino acids) in the development of functional aquafeeds, which can contribute to significantly reduce the abusive use of antimicrobials (e.g., antibiotics) and disinfectants [
3]. Functional diets are those that extend beyond satisfying the basic nutritional requirements of the farmed fish, contributing toward optimal growth, health, and survival through the inclusion of specific additives with health and growth-promoting properties [
4]. Moreover, it is known that amino acid (AA) requirements are expected to increase under challenging rearing conditions, so dietary AA supplementation beyond the nominal requirements for each species could benefit animals (e.g., improved immune status and increased tolerance to environmental stresses), as already reviewed elsewhere [
5,
6]. Thus, research on dietary AA supplementation as potential functional additives (i.e., supplied at levels beyond the species’ requirements) in fish, with particular emphasis on their effects on the immune system, should be better explored.
Histidine (His) is an essential AA (EAA) abundant in plasma albumin and skeletal muscle in fish [
7]. His participates in one-carbon unit metabolism, consequently affecting DNA and protein synthesis [
8]. Directly or through its derivative compounds, histidine plays an important role in homeostatic maintenance, osmoregulation, muscle pH buffering, and detoxification of reactive carbonyl species [
9,
10]. In humans, His also contributes significantly to the buffering capacity of plasma and tissue proteins [
11]. The protein buffer effect is the result of their dissociable side groups. For most proteins including hemoglobin, the most important of these dissociable groups is the imidazole ring of His residues. The His metabolite carnosine (beta-alanyl-L-histidine) also combats intramuscular acidosis by maintaining intracellular and extracellular buffering in muscle tissue pH [
12]. Moreover, various studies indicate that His and its derivatives act as antioxidants or can mitigate the impact of oxidative stress [
13,
14,
15]. Histidine and its imidazole derivatives such as carnosine and anserine have been proven to scavenge reactive oxygen species, and also contribute to improve taste, texture, and overall fillet quality [
9,
11,
16]. Additionally, a study with juvenile grass carp fed a His-deficient diet demonstrated significantly increased osmotic fragility of erythrocytes [
16]. Thus, histidine supplementation is expected to modulate both fish antioxidant status and immune response.
Threonine (Thr) is often considered the third limiting AA after lysine and methionine for growing fish fed low fishmeal feed formulations [
17,
18]. It is also involved in many physiological and biochemical processes including growth and immune functions [
18,
19,
20]. Duval, et al. [
21] discovered that Thr-enriched cell culture medium prevented apoptosis, stimulated mouse hybridoma cell growth, and promoted antibody production in lymphocytes through protein synthesis and cellular signaling mechanisms. Additionally, Thr deficiency upregulated nitric oxide levels in blood monocytes of broilers [
22]. Dietary Thr inclusion at 13.9 mg/g diet improved growth, digestive and absorptive capacity as well as the antioxidant status in intestine and hepatopancreas of sub-adult grass carp [
23]. Thr is also a major component of mucin in the small intestine of animals, which suggests its importance in the regulation of intestinal barrier integrity and function [
24,
25]. Therefore, the expectation is that a dietary Thr surplus will modulate fish mucosal health.
Taurine (Tau) is a conditionally essential nutrient that, although technically considered as an amino sulfonic acid due to its chemical structure [
7,
26], is often referenced to as an amino acid (in the sense of a small aminated acid). Tau can be synthesized from methionine and cysteine in fish, but the rate of synthesis is usually too low to satisfy the nutritional requirements of (at least) carnivorous fish [
27]. Hence, growth depression is one of the first clear observed signs reported during Tau deficiency [
26]. In fact, Tau supplementation in rainbow trout (
Oncorhynchus mykiss) fed a fishmeal-free diet improved growth and feed efficiency [
28]. Taurine’s role across numerous biological processes in fish has already been described, with a range of physiological problems and histological changes having been reported when Tau levels are reduced in the diet, namely green liver syndrome, reduced hematocrit, anemia, and reduced disease resistance [
26]. Taurine has also been described to play an important osmoregulatory role in fish [
29]. At the moment, information suggesting that Tau plays a role in fish health and immunity is still very fragmented. Nevertheless, Maita, et al. [
30] reported that yellowtail (
Seriola quinqueradiata) fed a fishmeal-free diet supplemented with Tau displayed improved fish survival in response to an artificial bacterial challenge, reaching similar levels as those fed a fishmeal-containing control diet, suggesting it may have immunoregulatory properties in fish. Hence, taurine supplementation is expected to possibly improve fish growth and feed efficiency, while modulating the immune system.
Several seabream species are farmed worldwide due to their savory meat and to meet its growing consumption trend. Among Sparidae, the gilthead seabream (
Sparus aurata, L.) is one of the main carnivorous farmed fish species in the Mediterranean region [
31]. A sustainable and profitable aquaculture relies on the production of healthy fish, which requires balanced feeds manufactured with high-quality ingredients [
32,
33]. The main goal of this study was to evaluate the effects of His, Thr, and Tau on gilthead seabream (
Sparus aurata) immune function when added as a supplement to a practical aquafeed formulation during a transient (two weeks) and short-term (four weeks) feeding period.
2. Materials and Methods
2.1. Diet Formulations
Extruded feeds were based on plant proteins sources and limiting fish meal inclusion to 12%. This mimics most of the currently used commercial diets for gilthead seabream. Using this basal formulation, three other experimental diets were produced at SPAROS Lda. (Olhão, Portugal) through the inclusion of crystalline His, Thr, and Tau.
Briefly, a control (CTRL) diet was formulated to meet current commercial formulations for this species as well as its known nutritional requirements. Three other diets were identical to the CTRL diet but supplemented with 0.4% His, 0.75% Thr, or 0.5% of Tau of feed (
Table 1). These inclusion levels were chosen to be at least 50% above the requirement levels established for gilthead seabream [
34,
35]. A single common base formulation was prepared and extruded, with the four different diets being obtained by the application of different vacuum coatings post-extrusion: the CTRL diet was subjected to vacuum coating with oil, while the other three (TAU, HIS, and THR) were coated with a mixture of oil and the corresponding AA, in order to attain the target AA supplementation levels. Main ingredients were ground (below 250 μm) in a Hosakawa, model #1 micropulverizer hammer mill (Hosokawa Micron Ltd., Preston Brook-United Kingdom). These ground ingredients were then mixed according to the target formulation in a Double-helix Mixture TGC, model 500 L (TGC Extrusion, Roullet-Saint-Estèphe, France), to attain a basal mixture (no oils were added at this stage). All diets were manufactured by extrusion (pellet size 2.0 mm) by means of a pilot-scale twin-screw extruder CLEXTRAL BC45 (Clextral, Firminy, France) with a screw diameter of 55.5 mm and temperature ranging 105–110 °C. Upon extrusion, all batches of extruded feeds were dried in a convection oven (OP 750-EF, LTE Scientifics, Greenfield, UK) for 2 h at 60 °C. After this process, pellets were left to cool at room temperature, and subsequently the AA (taurine, histidine, and threonine for each of the correspondent experimental diets) were mixed with the fish oil fraction according to each target formulation and added under vacuum coating conditions in a Pegasus vacuum mixer (PG-10VCLAB, Dinnisen, Sevenum, Limburg, The Netherland). Proximate composition analysis was performed by the following methods: dry matter, by drying at 105 °C for 24 h; ash, by combustion at 550 °C for 12 h; crude protein (N × 6.25), by a flash combustion technique followed by gas chromatographic separation and thermal conductivity detection (LECO FP428); fat, after petroleum ether extraction, by the Soxhlet method; total phosphorus, according to the ISO/DIS 6491 method, using the vanado-molybdate reagent; and gross energy, in an adiabatic bomb calorimeter (IKA).
Total AA content of diets was determined by hydrolyzation in 6 M HCl at 116 °C for 2 h in nitrogen-flushed glass vials. Samples were then pre-column derivatized with Waters AccQ Fluor Reagent (6-aminoqui-nolyl-N-hydroxysuccinimidyl carbamate) using the AccQ Tag method (Waters, Milford, MA, USA). Analyses were done by ultra-high performance liquid chromatography in a Waters reverse-phase AA analysis system using norvaline as an internal standard. During acid hydrolysis, asparagine is converted to aspartate and glutamine to glutamate, so the reported values for these AAs represent the sum of the respective amine and acid. The resultant peaks were analyzed with EMPOWER software (Waters, Milford, MA, USA). Tryptophan was independently determined by HPLC, after alkaline hydrolysis (Silliker Portugal, S.A. Vila Nova de Gaia-Portugal) The AA profiles of the experimental diets are presented in
Table 2.
2.2. Rearing Conditions
The current trial was conducted by trained scientists (following FELASA category C recommendations) and according to the animal experimentation guidelines on the protection of animals used for scientific purposes from the European directive 2010/63/UE at the experimental facilities of I3S (Instituto de Investigação e Inovação em Saúde, Porto, Portugal).
Gilthead seabream juveniles with an average weight of 8.77 ± 0.44 g were acquired from a certified hatchery (SONRÍONANSA; Cantabria, Spain) and were randomly distributed into 12 tanks (200 L;
n = 70) for an acclimatization period of three weeks being fed the CTRL diet. Fish were held in a recirculation seawater system in which oxygen concentration (7.3 ± 0.01 mg L
−1) and photoperiod were automatically controlled (10 h dark and 14 h light). Temperature was maintained by a water heater/cooler system at 20 ± 0.5 °C and kept unchanged throughout the experiment. Both nitrite and ammonium levels were daily recorded, and its levels controlled by a water ozonizer system. Water renovations and system cleanings were performed twice a week. After the quarantine period, the experiment was started by feeding each group (three tanks) with the respective feed three times a day
ad libitum. The control (CTRL) group was fed a control diet, which met the EAA requirement levels estimated for gilthead seabream [
34,
35]. The other three groups were fed diets with the inclusion of Thr, Tau, or His (THR, TAU, and HIS dietary treatments, respectively), as stated in
Table 2 for one month. No deaths during the experimental trial were observed.
2.3. Experimental Procedures
An initial sampling point was set, and 10 fish fed the CTRL diet were euthanized and designated as time zero (TØ) to assess their immune status prior to the feeding trial.
The feeding trial lasted four weeks in order to assess the effect of short- and mid-term dietary AA supplementation. Feed intake was recorded daily and body weight of fish fed dietary treatments was measured at the beginning and at the end of the feeding trial. Growth was monitored by obtaining the initial body weight (IBW) and final body weight (FBW) and used to calculate growth performance parameters.
Zootechnical performance measures were calculated as:
where g = (ln (Final body weight) − ln (Initial body weight))/days, calculated as a tank average.
At two and four weeks after feeding the experimental diets, 36 fish from each group (i.e., 12 per replicate) were euthanized by anesthetic overdose with 2-phenoxyethanol and individually weighed. Samples were obtained for immunological and transcriptomic studies. Skin mucus was collected by gently scraping the fish dorsal-lateral surface using a cell scraper with enough care to avoid contamination with blood, urogenital, and intestinal excretions, snap frozen in liquid nitrogen and stored at −80 °C. Blood was collected from the caudal vein using heparinized syringes, centrifuged at 10,000× g for 10 min and plasma pools were stored. Blood from four fish per tank was also used to perform blood smears. Head-kidney was collected and snap frozen for gene expression analysis. All samples were immediately kept at −80 °C until further processing.
2.4. Hematological Procedures
Blood was collected from the caudal vein using heparinized syringes. The hematological profile consisted of total white (WBC) and red (RBC) blood cells counts and blood smears for differential WBC counting as described by Machado, et al. [
36].
Blood smears were initially fixed with formol-ethanol (formaldehyde at 4%) and afterward stained with Wright’s stain (Haemacolor; Merck, Darmstadt, Germany). Neutrophils were identified according to their peroxidase activity, which was detected using the method described by Afonso, et al. [
37]. The slides were examined under oil immersion (1000×) and at least 200 leucocytes per slide were counted and classified as thrombocytes, lymphocytes, monocytes, and neutrophils. The relative proportion of each cell type was subsequently calculated.
2.5. Immune Parameters
Plasma bactericidal activity was measured using
Vibrio anguillarum according to Graham, et al. [
38], adapted by Machado, Azeredo, Diaz-Rosales, Afonso, Peres, Oliva-Teles and Costas [
36], with some modifications. Succinctly, 20 µL of plasma was added as duplicate to wells of a U-shaped 96-well plate. Hanks’ Balanced Salt solution (HBSS) was added to four wells instead of plasma and served as a positive control. To each well, 20 µL of
V. anguillarum (1 × 10
6 cfu mL
−1) were added and the plate was incubated for 3 h at 25 °C. To each well, 25 µL of iodonitrotetrazolium chloride, INT (2-(4-iodophenyl)-3-(4-nitrophenyl)-5-phenyl-2H-tetrazolium chloride; 1 mg.mL
−1; Sigma-Alrich, Steinheim am Albuch, Germany) was added to allow the formation of formazan. Plates were then centrifuged at 2000×
g for 10 min and the precipitate was dissolved in 200 µL of dimethyl sulfoxide (Sigma). The absorbance of the dissolved formazan was measured at 490 nm in a Synergy HT microplate reader (Biotek, Winooski, Vermont, USA). Total bactericidal activity is expressed as the percentage of killed bacteria, calculated from the difference between the samples (surviving bacteria) and the positive control (100% living bacteria).
IgM in plasma was measured by an Enzyme-Linked Immunosorbent Assay (ELISA assay). Succinctly, 4 µL of plasma were previously diluted (1:100) in 396 µL of Na2CO3 (50 mM, pH = 9.6) buffer and then 100 µL of these diluted plasma samples were added to the 96 well in duplicates using 100 µL of buffer (Na2CO3) as a negative control (added to four wells). The samples (antigen) were allowed to stand at 22 °C for 1 h, being subsequently removed by means of an aspirator with a pipet tip. Then, 300 µL of blocking buffer (5% low fat milk powder in 0.1% Tween 20) was added to each well and incubated for 1 h at 22 °C. This mixture was then removed by aspiration and followed by three consecutive washes with 300 µL of T-TBS (0.1% Tween 20). T-TBS (0.1% Tween) is a solution made with 20 mM Tris Base (Nzytech) and 137 mM NaCl (Sigma), distilled water, and TWEEN 20 (Sigma). After properly cleaning and drying the wells, 100 µL of the anti-gilthead seabream primary IgM monoclonal antibody previously diluted in blocking buffer (1:100) was added to each well and incubated for 1 h at 22 °C. The primary antibody was then removed by aspiration, with three consecutive washes being performed. Afterward, the anti-mouse IgG-HRP secondary antibody diluted 1:1000 in blocking buffer was added and incubated for 1 h at 22 °C, then being recovered by aspiration. The wells were then washed three times and 100 µL of 3,3′,5,5′-tetramethylbenzidine hydrochloride (TMB) substrate solution for ELISA (BioLegend #421101), previously prepared, was added to each well and incubated for 5 min. The color change reaction was stopped after 5 min by adding 100 µL of 2 M sulfuric acid and the optical density was read at 450 nm.
The antiprotease activity was determined as described by Ellis [
39] adapted by Machado, Azeredo, Diaz-Rosales, Afonso, Peres, Oliva-Teles and Costas [
36]. Briefly, 10 µL of plasma were incubated with the same volume of trypsin solution (5 mg.mL
−1 in NaHCO
3, 5 mg.mL
−1, pH 8.3) for 10 min at 22 °C in polystyrene microtubes. To the incubation mixture, 100 µL of phosphate buffer (NaH
2PO
4, 13.9 mg.mL
−1, pH 7.0) and 125 µL of azocasein (20 mg.mL
−1 in NaHCO
3, 5 mg.mL
−1, pH 8.3) were added and incubated for 1 h at 22 °C. Finally, 250 µL of trichloroacetic acid were added to each microtube and incubated for 30 min at 22 °C. The mixture was centrifuged at 10,000×
g for 5 min at room temperature. Afterward, 100 µL of the supernatant was transferred in duplicates to a 96-well plate that previously contained 100 µL of NaOH (40 mg.mL
−1) per well. The OD was read at 450 nm. Phosphate buffer was added to some wells instead of plasma and trypsin and served as a blank, whereas the reference sample was phosphate buffer instead of plasma. The percentage of inhibition of trypsin activity compared to the reference sample was calculated.
Total peroxidase activity in plasma was measured according to the procedures described by Quade and Roth [
40]. Briefly, 15 µL of plasma in duplicates were diluted with 135 µL of HBSS without Ca
2+ and Mg
2+ in flat-bottomed 96-well plates. Then, 50 µL of 20 mM 3,3′,5,5′-tetramethylbenzidine hydrochloride (TMB; Sigma, Alrich, Steinheim am Albuch, Germany) and 50 mL of 5 mM H
2O
2 were added. The color change reaction was stopped after 2 min by adding 50 mL of 2 M sulfuric acid and the optical density was read at 450 nm. Wells without plasma were used as blanks. One unit of peroxidase activity (units.mL
−1 plasma) was defined by the quantity of peroxidase that produces an absorbance change of 1 OD.
2.6. Gene Expression Analysis
Head-kidney were taken from fish fed the experimental diets for two and four weeks. Genes were analyzed using the seabream PCR-array platform of the IATS Nutrigenomics group (
http://nutrigroup-iats.org accessed on 7 March 2021). Total RNA was extracted using a MagMAX
TM-96 total RNA Isolation Kit (Life Technologies, Carlsbad, CA, USA). RNA yield was 50–100 μg with 260 to 280 nm UV absorbance ratios (A260/280) of 1.9–2.1. Reverse transcription (RT) of 500 ng total RNA was performed with random decamers using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA), according to the manufacturer’s instructions. Negative control reactions were run without reverse transcriptase.
Real-time quantitative PCR was carried out on a CFX96 Connect
TM Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) using a 96-well PCR array layout designed for simultaneously profiling of a panel of 29 genes for head-kidney (summarized by biological processes in
Table 3). Selected genes were associated with health biological processes, like interleukins and cytokines (9), macrophage and monocyte chemokines (3), immunoglobulins (4), antiprotease (1), antimicrobial peptide/iron recycling (1), T-cell markers (6), and pattern recognition receptors (5). Controls of general PCR performance were included on each array with all the pipetting operations performed by means of the EpMotion 5070 Liquid Handling Robot (Eppendorf, Hamburg, Germany). Briefly, RT reactions were diluted to convenient concentrations and the equivalent of 660 pg of total input RNA was used in a 25 μL volume for each PCR reaction.
PCR-wells contained a 2× SYBR Green Master Mix (Bio-Rad) and specific primers at a final concentration of 0.9 μM were used to obtain amplicons of 50–150 bp in length (
Table S1). The program used for PCR amplification included an initial denaturation step at 95 °C for 3 min, followed by 40 cycles of denaturation for 15 s at 95 °C and annealing/extension for 60 s at 60 °C. The efficiency of PCR reactions was always higher than 90%, and negative controls without sample templates were routinely performed for each primer set. The specificity of reactions was verified by analysis of melting curves (ramping rates of 0.05 °C/s over a temperature range of 55–95 °C), and linearity of serial dilutions of RT reactions. Fluorescence data acquired during the PCR extension phase were normalized using the delta–delta Ct method [
41]. β-Actin was tested for gene expression stability using GeNorm software (M score = 0.21) and used as a housekeeping gene in the normalization procedure. Fold-change calculations were done in reference to the mean response of CTRL fish. To compare the mRNA gene expression level of a panel of genes in a given dietary treatment, all data values were expressed in reference to the expression level of
casp3 in CTRL fish, which was arbitrarily assigned a value of 1.
2.7. Statistical Analysis
All results were expressed as means ± standard error (SE). Univariate statistic evaluation of the data was accomplished by two-way ANOVA with sampling time and dietary treatment as main factors. A significance of p < 0.05 was applied to all statistical tests. Gross deviations from the ANOVA assumptions of error normality and homoscedasticity were evaluated through residual analysis (using QQ-plots and “residuals vs. fitted” scatter plots). All tests were run with SPSS statistical analysis software (SPSS ver.26.0; Chicago, IL, USA).
For gene expression data, unsupervised principle component analysis (PCA) was first performed on data as an unbiased statistical method to observe intrinsic trends in the dataset using EZ-INFO
® v3.0 (Umetrics, Malmö, Sweden). To achieve the maximum separation between groups, supervised partial least-squares discriminant analysis (PLS-DA) was subsequentially applied. Potential differential genes were selected according to the Variable Importance in the Projection (VIP) values. Variables with VIP > 1 were considered to be influential for the separation of samples in PLS-DA analysis [
42,
43,
44].
4. Discussion
The present study showed that some modulation of leucocyte response was observed in fish fed a TAU dietary treatment, with the number of circulating neutrophils being increased regardless of sampling time. There is no published information about Tau involvement in differentiation of this cell-type, but it is known that neutrophils contain very high concentrations of taurine due to its uptake from the blood [
45]. Hence, some stimulation of Tau levels on neutrophil numbers could be postulated and it could also be speculated that Tau requirement may increase upon infection. Neutrophils from the reserve pools are quickly attracted to infectious foci by microbial products and chemotactic substances released by host cells [
46].
Moreover, plasma antiprotease increased in fish fed THR dietary treatment compared to those fed the CTRL diet, which could indicate a potential capacity of this particular AA to boost the host’s innate immune defenses. Antiproteases play a crucial role in the inhibition of certain proteases either by “trapping” proteases to avoid protein hydrolysis or by binding to their active sites [
47,
48]. The observed effect of a dietary Thr surplus may also translate into an improved resistance against bacteria in animals fed higher levels of this AA. Additionally, several pathogens (mainly bacteria) express a variety of non-specific proteases that degrade important proteins involved in fish innate immunity [
49,
50]. Hence, the presence of stronger antiprotease activity in seabream fed the THR dietary treatment may indicate an improvement in fish immune condition.
Skin mucus is an important physical and chemical barrier against invading pathogens. It serves as an important component of the innate immune machinery by: (i) its continuous renewal, preventing stable colonization of the skin by pathogens; and (ii) containing a number of innate immune proteins and enzymes (e.g., lysozyme, complement proteins, and antibacterial proteins and petites) including antiproteases [
51]. In the present study, seabream fed HIS dietary treatment showed an increase of total antiprotease activity measured in skin mucus. In contrast, fish fed the HIS diet had a lower bactericidal activity in skin mucus compared to those fed the CTRL diet after two weeks, which seems to point to a differential immune status in fish fed this diet. One should take into account that the bactericidal activity is a multifactorial indicator since it evaluates a wide range of innate immune mechanisms and molecular defenses against bacterial invasion such as proteins of the complement system, antimicrobial peptides, acute phase proteins, immunoglobulins, lysozyme, and cytokines [
38,
52]. Therefore, and considering the decrease in lymphocyte numbers observed at four weeks, we suggest that dietary His supplementation can improve certain key aspects of innate immunity in detriment of factors contributing to acquired immunity at the skin mucus level.
The results obtained in terms of bactericidal activity are comparable with those of previous works where the impact of a Tau-rich diet on lysozyme and alternative complement system was evaluated. Han, et al. [
53] observed that Tau supplementation did not influence blood parameters (e.g., hematocrit, hemoglobin, glucose, total protein, total cholesterol, total bilirubin, and triglycerides) of Japanese flounder (
Paralichthys olivaceus). This was corroborated by Magalhães, et al. [
54] in a similar study with white seabream (
Diplodus sargus) juveniles, where no differences were observed in the plasma alternative complement pathway and lysozyme levels. Nonetheless, a diet rich in taurine boosted the lysozyme activity and other immune parameters such as phagocytic index, respiratory burst, and total immunoglobulin content, in yellow catfish (
Pelteobagrus fulvidraco) [
55]. This highlights the importance of further studies to clarify the effects of Tau on fish immune system, since it could be species and context-dependent.
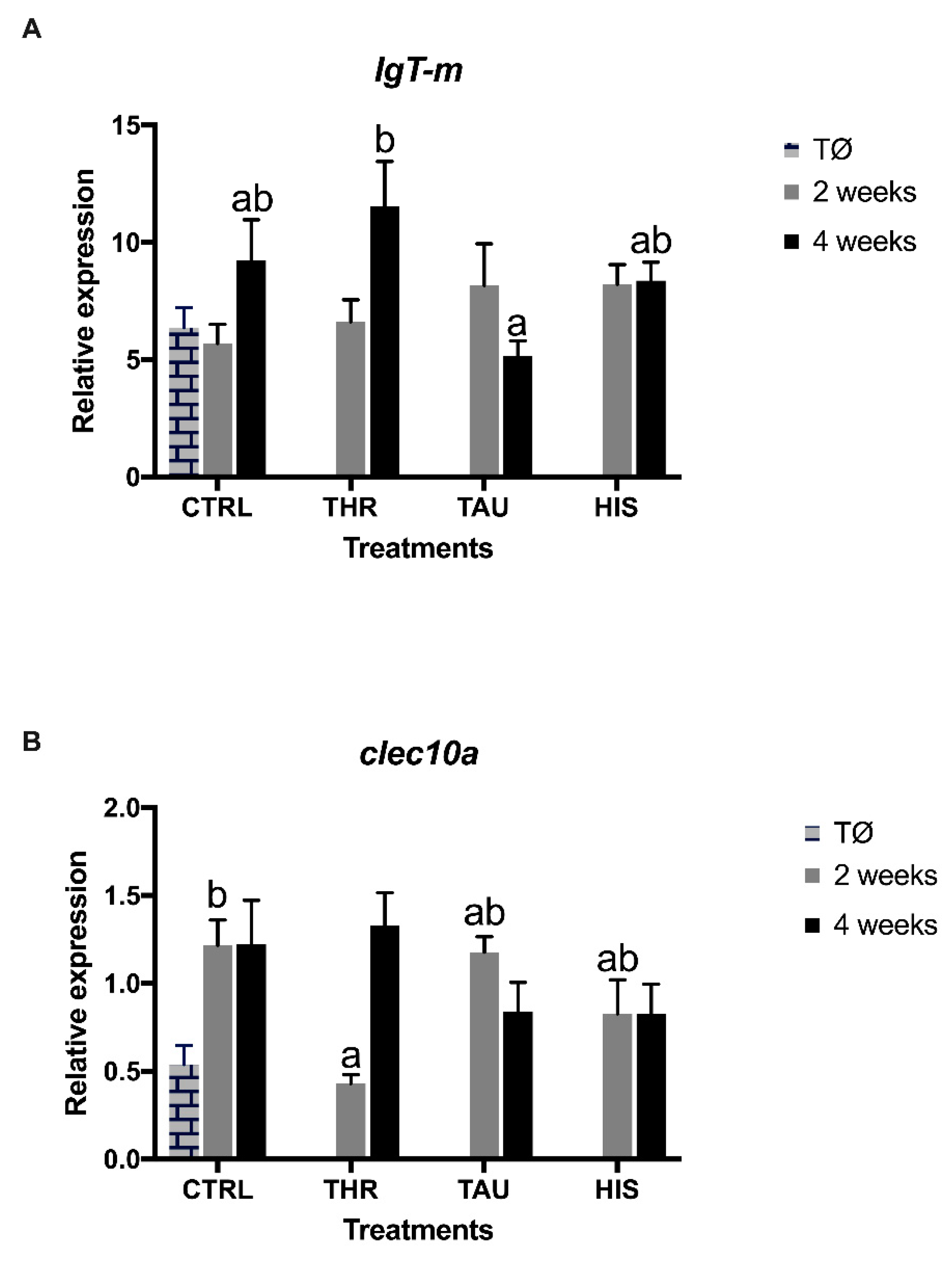
In the present study, dietary treatments did not induce strong transcriptional changes, as the observed differences were mostly time-dependent. Nonetheless, some mild effects were observed with the downregulation of clec10, a member of the C-type lectin (CTLs) superfamily, in seabream fed the THR dietary treatment for two weeks.
Lectins are a family of glycoprotein pattern-recognition receptors highly represented in fish, typically being multivalent proteins that recognize and bind specific carbohydrate recognition domains (CRD) [
56]. The presence of several CRDs, in combination with other proteins domains, enables not only the recognition of specific carbohydrates on the pathogen’s surface, but also on the surface of immunocompetent cells. Some lectins induce the synthesis of pro-inflammatory cytokines including
il1-β1,
il1-β2,
tnf-α1,
tnf-α2, and
il8 in rainbow trout macrophages and fibroblast-like cells [
57]. Lectins play an active role in innate immunity in PAMP recognition, opsonization, phagocytosis, and complement activation [
58,
59]. C-type lectins, in particular, have several functions in innate immunity and contribute to the homing of leucocytes and immune cell trafficking as well as pathogen recognition and subsequent T cell activation [
60]. As such, CLEC10 is likely to play a role in regulating adaptive and innate immune responses and, thus, in this study, the downregulation of this CTL in seabream fed THR diet for two weeks could be suggested, and the concomitant decrease in blood total WBC, as a possible inhibition of immune alertness (e.g., pathogen recognition and innate immunity).
Overall, some unexpected variation in the AA concentrations measured in feeds were observed. On the other hand, given that all diets were produced from the same basal formulation, we expected true differences in non-supplemented AA to be smaller than that measured. Thus, we expected the observed effects to be mostly attributable to the increase in threonine, histidine, and taurine.
Moreover, Ramos-Pinto, Azeredo, Silva, Conceição, Dias, Montero, Torrecillas, Silva and Costas [
32] described that arginine and citrulline, particularly when supplemented at a 1% of feed inclusion level, induced a stimulation of the fish immune system after a short-term feeding period, verified mostly by a modulation of the gilthead seabream plasma proteome and health-biomarkers after four weeks of exposure. Hence, the fact that arginine concentration was higher in the CTRL diet compared to the others could have translated into an increased gilthead seabream immune status. It can therefore be speculated that an eventual immune modulation in fish fed the CTRL diet due to higher arginine levels could have attenuated more positive effects regarding dietary threonine, histidine, or taurine supplementation.
In the present study, data from the analyzed responses of fish fed supplemented diets suggest that the effects of Thr, His, and Tau supplementation on fish immune system seem to be of an indirect nature, when compared with other EAAs previously studied such as tryptophan, methionine, or arginine [
32,
61,
62], which present stronger and more direct effects. In a previous work where the effects of dietary tryptophan supplementation were also explored in gilthead seabream, only a transient immune enhancement was observed in fish fed an extreme formulation (no FM inclusion) over a short-term feeding period (two weeks), suggesting that these putative advantageous effects seem to disappear over a longer feeding period (13 weeks) [
33]. The present study also underlines the fact that “feeding time”, besides “dosage”, seems to be a crucial factor in determining the AA efficacy. It seems that supplements, in this case AA, may lose their effectiveness after being available for a long feeding period. A possible explanation can be assumed: fish becomes adapted to this supplementation level through some physiological/metabolic readjustment. Therefore, the importance of “exposure time” as a central factor in determining the apparent functional effects of additives seems to be of major importance. In the present study, this fact seems to be particularly relevant for the THR dietary treatment. While a decrease in total WBC numbers and
clec10 transcripts was observed after two weeks of feeding, the acquired arm of the immune system was stimulated by dietary Thr supplementation with an enhancement of peripheral lymphocytes and
IgT-m mRNA expression level in the head-kidney after four weeks of feeding.


 ,
, 


{kind=link}