4.1. Dry Matter Intake, Milk Yield, and Nutrient Digestibility
The results of this experiment showed that diets with oils rich in PUFA in a moderate amount reduced DM intake, which in turn resulted in lower intakes of other nutrients (OM, CP, NDF, WSC, and starch). Similar observations have been reported previously in experiments with PUFA-rich plant oils [
28,
29], fish oil [
30,
31], or a mixture of fish oil and plant seed oils [
32,
33,
34]. In this experiment, dietary oil supplementation reduced DM intake by 9.7% on average. In line with these results, Shingfield et al. [
33] reported approximately 20.5% reduction in DM intake as a result of adding a mixture of fish and sunflower oils to corn silage-based diet (45 g oil/kg DM and 63 g ether extract (EE)/kg DM) compared with a non-supplemented diet (EE content of 33.5 g/kg DM). However, another study reported no difference in terms of DM intake of cows between the control diet based on alfalfa and corn silage (27.8 g EE/kg DM) and the same diet supplemented with mixture of fish or canola oils (20 g oil/Kg DM and 46.7 g EE/kg DM, respectively; [
31]). It has been well shown that the effect of oil supplementation of a diet on DM intake can be a function of combination of including oil content and diet composition, source of oil, and type of basal diet [
29,
35,
36].
The lack of responses on milk yield due to dietary starch level and unsaturated oil mixture were expected as the diets were designed based on our previous experiences [
10] to cause no effect on milk yield but lower fat and ECM yields due to the combination of high starch level and oil (i.e., HSO diet) known as milk fat depression effect. However, ECM yield was not influenced by dietary treatments despite being noticeably yet numerically lower for HSO diet (2.7 kg/d) compared with the average of other treatments. The reason for numerically lower ECM was the lower milk fat concentration (11% lower compared with LS diet).
The lack of oil supplementation effect on nutrient digestibility, with exception of CP and GE digestibility is consistent with some previous findings [
14,
37,
38].
While in our experiment high-starch diet did not affect DM intake, an increase was observed for CP, OM, and starch intakes with drops in NDF and WSC intakes, which are due to differences in nutrient contents of the diets. The results of our study are consistent with the findings of Pirondini et al. [
39] and Philippeau et al. [
40]. In contrast, a 4.3% drop in DM intake for cows fed high- compared with low-starch diets based on grass silage (212 vs. 116 g starch/kg DM) has been reported [
41]. Starch effect on feed intake can be mediated by a number of factors including starch fermentation rate, forage content of diet, amount of metabolic fuel absorbed from the rumen (for instance VFA), rumen pH, and rumen fermentation parameters [
42,
43]. It should be noted that the source of starch in diet and processing method can also contribute to the mixed results of different experiments. Hatew et al. [
41] attributed the lower DM intake in high starch diets to increased propionate concentration in the rumen since hepatic oxidation of propionate influences DM intake [
44]. Therefore, as in the aforementioned studies, no significant change was found in ruminal propionate concentration which is consistent with unaffected DM intakes.
In our experiment, digestibility of DM, OM, starch, and GE was higher and CP digestibility tended to be higher in cows fed HS than those receiving LS diets. Beckman and Weiss [
45] observed linear increase in DM and OM digestibility as a result of reduction in NDF:starch ratio and linked it to replacement of highly digestible carbohydrates (i.e., starch) with low-digestible carbohydrates (i.e., NDF). In the current experiment, lower NDF and pdNDF digestibility along with improved starch digestibility beside the lack of effect on rumen pH as a factor influencing fiber digestion, might reflect the competition between rumen microbial population to utilize more easily nonstructural carbohydrates when available. As shown by the results of the microbiological analysis, the abundance of
Ruminobacter sp. or
Selenomonas sp. increased in high starch diets while the abundance of amylolytic Prevotella or cellulolytic bacteria Ruminococcus and Fibrobacter in LS-fed cows was not significantly different from that in HS-fed cows. Another explanation might be that observed differences in fiber digestibility reflect changes in the composition of diets; in HS diet rolled barley and ground wheat were used instead of sugar beet pulp and barley feed which might have different fiber characteristics.
It should be noted, however, that in some experiments where starchy feeds were replaced by fibrous by-products, there has been a confounding effect of both starch:NDF ratio and forage proportion of the diet [
45]. However, in our experiment, the forage proportion in both LS and HS diets were fixed to remove such a source of difference between diets.
4.2. Methane Production and Rumen Fermentation
The oil supplementation to both low- and high-starch diets resulted in lower CH
4 production compared with non-supplemented diets (475 vs. 552 g/d, on average). This is in line with the findings of the experiments that used oilseeds [
29,
46,
47], or fish oil [
30] to supplement dairy cow diets. In the current experiment, enteric CH
4 emission (g/d) reduced by on average 4.7% for each 10 g/kg DM unsaturated oil in the diet. Similarly, Martin et al. [
4] indicated that CH
4 production reduces by 3.8% as a result of every 10 g/kg increase in dietary lipid supplementation. The most of this response in our experiment was caused by lower feed intake as CH
4 yield (g/kg OMI) was not affected by the oil supplementation (average numerical reduction of 1.7% in CH
4 yield). Ramin and Huhtanen [
48] in a meta-analysis showed that 1 g/kg of DM increase in dietary EE concentration decreased CH
4 yield by 0.043 L/kg of DM. The equivalent value in our experiment was on average 0.045 g/kg of OM intake which is very close to the reported value. Apparently both biohydrogenation of unsaturated fatty acids and amount of unfermentable organic matter introduced by oil to the diet should have caused the lower CH
4 yield. Oil potential in reducing CH
4 production seems to be a function of factors such as source of oil, fatty acid composition and level of supplementation in the diet [
2,
4]. Several mechanisms are known to influence the impact of fats and oils in reducing CH
4 production including reduced OM fermentation in the rumen, unfavorable effects of C12:0 and C14:0 on protozoa community, and inhibition of methanogens by 18-carbon unsaturated fatty acids [
4,
49,
50] and the competition for using hydrogen for biohydrogenation of unsaturated fatty acids. Patra [
51] reported that C12:0 and C18:3 fatty acids are stronger inhibitors of methanogenesis compared with other fatty acids. Furthermore, as far as reduction in CH
4 production is concerned, MUFA and PUFA are more effective than SFA in the diet. In this experiment, oil supplementation to both low- and high-starch diets resulted in higher intakes of both MUFA and PUFA compared with non-supplemented diets.
Unaffected CH
4 yield for cows fed oil-supplemented diets in our experiment is consistent with the lack of differences in rumen pH, acetate, propionate, and VFA concentration and acetate:propionate ratio, although acetate and propionate concentrations and acetate:propionate ratio decreased numerically. However, oil supplementation is expected to not only reduce CH
4 production but also lower acetate:propionate ratio. The non-significant change in acetate:propionate ratio observed in our experiment can be attributed to the lower DM and concentrate intakes, and the forage portion of our experimental grass silage-based diet. Changes in rumen fermentation pattern due to oil supplementation with diets based on restrictively fermented grass silage (using the silage additive based on formic acid) may be resistant to lipid supplementation [
7].
Even though the intake of GE increased as a result of including oil in LS and HS diets, there were no significant differences between experimental treatments in terms of CH
4 production as a percentage of GE intake while CH
4 intensity calculated as g/kg milk or ECM decreased and tended to decrease by oil supplementation, respectively. In fact, the reduction in CH
4 intensity indicates that net energy is partitioned more towards milk production, leading to lower CH
4 intensity [
3]. The values of CH
4 production as a percentage of GE intake for un-supplemented diets (LS and HS; 7.25 and 7.04%) having DM intake of 23.0 and 23.5 kg/d are higher than 6.4% measured from dairy cows receiving rather similar diets with similar DM intake in respiration chambers (Bayat et al., unpublished data).
Our findings showed that starch level did not influence ruminal CH
4 and CO
2 production (g/d) and emission intensity (g/kg milk yield). Previous studies [
39,
40,
41] have reported lower or tendencies towards lower daily CH
4 production and CH
4 emission intensity with the exception of Hatew et al. [
41] reporting a non-significant CH
4 emission intensity due to increasing dietary starch level. Increased starch content influenced rumen fermentation parameters, with a significant decrease in molar proportion of acetate and a tendency to reduce acetate:propionate ratio. Although high starch diets are expected to reduce acetate and increase propionate molar proportions and to lower CH
4 production [
4], this mechanism may not invariably apply to all experimental conditions. We did not observe any differences in rumen pH between cows fed low- and high-starch diets while it has been shown that high starch content lowers rumen pH, thereby limiting growth or activity of methanogens and cellulolytic bacteria [
4]. The higher rumen ammonia N concentration due to higher starch level can be attributed to slightly higher dietary CP level whereas the higher rumen ammonia N concentration with oil-containing diets might have arisen from adding urea, which is highly rumen-degradable, to the diets in an attempt to make them isonitrogenous.
4.3. Rumen Microbial Ecology
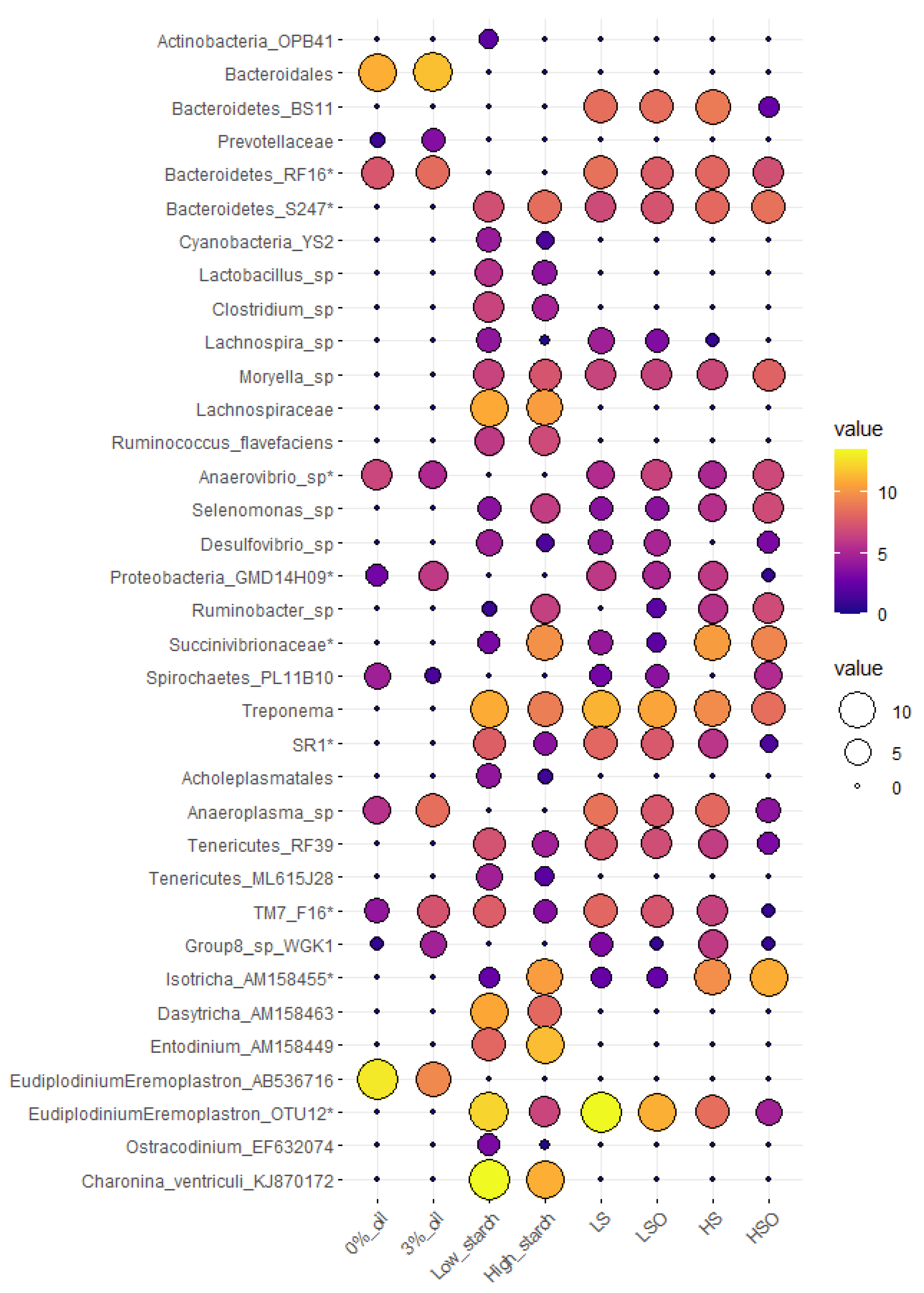
Starch level had stronger effect on bacterial richness compared with those caused by inclusion of oil in the diet. Despite reports of toxic effects of oils on microbial community and, therefore, greater expected changes in the rumen microbial composition [
52], this study did not observe significant oil effect on the reduction of alpha diversity in bacterial, archaeal or ciliate protozoan communities. Effects of oil on the microorganisms may depend on type and amount of oil, type of fatty acids in diet, and type of dietary forage fed. Bayat et al. [
7] showed that supplementation of dairy cow diets with plant oils like rapeseed, safflower, linseed or myristic acid reduced CH
4 production, with each type of oil having a different impact on bacterial community. Furthermore, Martin et al. [
37] reported that adding extruded linseed to hay-based diets or corn silage-based diets for dairy cows reduced CH
4 production without a notable change in abundance of rumen methanogens and cellulolytic bacteria. Looking at individual microbial taxa, addition of oil to both low- and high starch diets in this study, provided a suitable ecological niche for lipid hydrolyzing
Anaerovibrio sp. [
53] and Spirochaetes order PL-11B10. PL-11B10 was detected at significantly higher abundance in the diet with myristic acid supplement [
7] and outside ruminants has been found positively correlated with methane production in methanogenic oil wells [
54]. Nevertheless, our understanding of PL-11B10 ecology in rumen is limited. Oil supplementation also increased relative abundance of ciliate protozoa
Eudiplodinium-Eremoplastron (AB536716). Similarly, a significant increase in
Eremoplastron dilobum abundance was detected in an in vitro experiment with linseed oil but not rapeseed oil additive [
55], suggesting that a positive or negative oil effect is depends on protozoa species and oil type.
High starch diets increased abundances of known starch utilizers like
Ruminobacter sp.,
Selenomonas sp. or ciliate protozoa
Isotricha sp. and
Entodinium sp. that are involved in the utilization of non-structural polysaccharides and soluble sugars [
53]. The Bacteroidetes family S24-7 was also enriched in HS diets. Current research suggests that members of S24-7 family are versatile with respect to complex carbohydrate degradation, but starch utilization trait is common to all family members and increased abundance of S24-7 is correlated with increased propionate production [
56]. Inclusion of oil in LS and HS diets did not change total VFA, acetate, propionate, and butyrate but reduced CH
4 production, suggesting lower H
2 availability in these ruminal ecosystems. S24-7 was significantly more abundant in HSO diet, was strongly negatively correlated with acetate and tended (
p = 0.054) to be positively (R = 0.49) correlated with propionate concentration in the rumen (data not shown). In addition to S24-7, also
Prevotella sp.,
Moryella sp. and members from
Paraprevotellaceae family were positively correlated with propionate concentration in the rumen (
Figure 2). Given that propionate concentration was numerically the highest in HSO diet, these bacteria may have contributed to the sink in reduction of H
2 availability for methanogenesis.
Low starch diets were enriched with bacteria directly or indirectly involved in fiber degradation.
Clostridium,
Treponema, (TM7) F16, (Tenericutes) RF39, and
Desulfovibrio were found to be tightly attached to switchgrass [
57] or wheat straw [
58] during degradation process. In co-cultures with
Fibrobacter succinogenes,
Treponema bryantii has been shown to utilize soluble sugars released from cellulose degradation [
53]. In addition to bacteria, ciliate protozoa
Ostracodinium sp. and
Dasytricha sp., are known to contain cellulolytic and hemicellulolytic activities, respectively, and were significantly enriched in LS diets.
Charonina ventriculi is a holotrich protozoa not frequently observed in the rumen and with limited information about its metabolism. In our experiment,
Charonina ventriculi was significantly enriched in LS diets and was negatively correlated with ammonia-N and butyrate, but positively correlated with acetate concentration in the rumen. Correlation profile of
Charonina ventriculi was similar to
Treponema sp. and
Clostridium sp. (
Figure 2) suggesting that these microorganisms require similar rumen conditions for thriving or are involved in similar metabolic processes.
Methanobrevibacter gottschalkii and Mbb. ruminantium were the predominant archaea without being significantly affected by starch level or oil supplement, although oil numerically reduced their abundance. Methanobrevibacter are hydrogenotrophic methanogens that convert H2 and CO2 produced by protozoa, bacteria, and fungi to CH4. In our study, numerical decreases in both Mbb. gottschalkii and Mbb. ruminantium correlated with decrease in CH4 intensity (g/kg milk) in oil supplemented diets. With higher abundances of bacteria Moryella sp., Anaerovibrio sp., (Bacteroidetes) S24-7, (Spirochaetes) PL11B10, Selenomonas sp., Ruminobacter sp., (Paraprevotellaceae) YRC22 and ciliate protozoa Anoplodinium-Diplodinium which had a tendency to be positively correlated with propionate concentration in the rumen, we can hypothesize having an ecosystem with less hydrogen available for methanogenesis.
Reduced daily CH
4 emissions (g/d) were positively associated with reduction in
Entodinium caudatum. Although
Entodinium was the most abundant ciliate protozoa in all diets and smaller
Entodinium spp. have been suggested to contribute more to CH
4 production compared with larger protozoa in in vitro studies [
59], a deeper subdivision of
Entodinium into OTUs suggests functional versatility and differences in host dependency inside this genus Contrary to our results, Belanche et al. [
60] investigated holotrich protozoa role in CH
4 production compared to the natural flora and concluded that holotrichs were responsible for increased methanogenesis more than the entodiniomorphids. In our study, OTU affiliated with
Isotricha prostoma was negatively correlated with daily CH
4 emissions (OTU detected in 10 samples and therefore not included in
Figure 2), while other OTUs affiliated with holotrichs did not produce significant associations with CH
4 production. Discrepancies in observations could relate to the differences in the basal diet and dietary treatment as well as host impact on the general microbial community composition.
Based on the results of this experiment, it can be argued that inclusion of the mixture of fish oil and sunflower oil at 30 g/kg of diet DM does not have profound toxic effects on bacteria, archaea, or ciliate protozoa, and it is possible that the minor reduction in CH4 yield caused by inclusion of oil in diet may be linked more to the functional networks of microbiome possibly leading to a lower availability of hydrogen in the rumen which is required for CH4 production.


 ,
, 



{kind=link}
{kind=link}