The most difficult factor in either investigating or interpreting the protein requirement systems for the maintenance and growth of bovine animals is finding evidence that those requirements are influenced by inherent genetic variation and capability of animals to obtain greater protein synthesis as a considerably energy intake. To the best of the author′s knowledge, the current data show for the first time an investigation that predicts a protein requirement system for the maintenance and growth of 18–24 months old Thai swamp buffaloes. Therefore, the discussion will be based on providing different protein supplies with an abundance of energy supply for the maintenance and growth of Thai swamp buffalo. Protein requirement, nutrient utilization, and microorganism profile are included to elaborate the influences mentioned.
3.1. Body Weight and Daily Gain Performance
The different protein supplies with abundance of energy supply fed to Thai swamp buffaloes resulted in improving numbers of final weight and average of daily gain (
p < 0.05;
Table 2). The average of daily gain (ADG) throughout the 90 days of feeding trial was a gradually increased (
p < 0.05) in buffaloes receiving a greater protein level in diets. Despite buffaloes receiving a level of 5.4% protein in their diet, the present study indicating that those buffaloes had a higher protein efficiency, where buffaloes that received a range of 6.6–10.5% crude protein (CP) were able to get impressive ADG. Our results were in agreement with Chumpawadee, et al. [
32] who reported that an increase in ADG of Thai-indigenous yearling heifers was successfully obtained after those heifers were fed diets containing high CP (6.6–13.6%). The assumption that more protein supply is conducive to more weight gain might seem to be true according to the abovementioned. Other bull observations, nevertheless, concluded that excessively dietary CP had no effect on ADG in bull animal performance. For instance, Devant, et al. [
33] increased dietary CP level in diets ranging from 14.4–17.3%, and Promkot and Wanapat [
34] attempted to decrease the level of protein in diets ranging from 10.5–14.4%, and those observations reported that no gaining weight of crossed heifers occurred. Similarly, Basra, et al. [
35] suggested no shift of ADG, when CP level in diets were adjusted from 12.1% to 18.2% of DM in Nili-Ravi buffaloes; thus, suggesting that dietary CP over 10.0% fed to bull animals did not give a positive impact on gaining weight. Dietary CP in the range of 6.6–8.5% constituted an alternative strategy to improve body weight and ADG, without negatively affecting animal performance and farm budgeting. Other discussion had previously reviewed the efficiency of protein-rich diets to contribute to food security, employment, and rural economies [
36].
As expected, the aforementioned results of gained weights were derived from a greater feed efficiency (ADG/DMI, CPI or TDNI,
p < 0.01,
Table 2) as a result of receiving more protein supplies in diets. These results were similar with a prior study [
37], where Nili-Ravi buffaloes were fed diets containing CP (9.1%) and had a relatively abundant energy intake. These achievements, in turn, lead to lowering the feed efficiency and ADG pattern. If the reduction in a feed intake represents a shift of caloric restriction allowing hyperthermic animals to reduce heat generation [
38], a reduction in feed efficiency or ADG pattern was obtained from the limitation of nutrient digestion and metabolism in the rumen host permitting an alleviation of the protein synthesis rate, as indicated from the insufficient energy intake [
12].
3.2. Nutrient Intake and Nutrient Digestibility
Nutrient intakes beneath requirements by bull animals results in deferred maturity of the reproduction system and slowed down growth rates [
9,
11,
12]. Generally, all of the nutrient intakes on dry-matter (DM) basis increased, when dietary crude protein increased (
p < 0.01,
Table 3). CP intake (g/kg W
0.75) was a gradually increased in a range of 3.55–9.44 g/kg W
0.75. It could be noted that no buffalo had restricted the nutrient access per group per experimental design (
n = 4) in the present study. Thus, the latter outcomes could determine more investigation for nutrient or protein requirement of maintenance and growth in Thai swamp buffaloes. Moreover, the rate of nutrient intakes could be influenced by several factors such as rumen capacity, ruminal metabolic level (factual VFAs), digestion rate, physiological animal, and nutrient requirement setting [
39]. We know of few serial observations regarding dietary DM intake influenced by dietary CP as similar as the present study. Comparative studies observed in Nili-Ravi buffaloes [
35,
37], Thai-indigenous heifers [
32,
40], and Murrah buffaloes [
7,
11] showed that either protein or nitrogen intake was varied when those bull animals were fed to the extent of the dietary CP. In addition, the rumen capacity, including limited passage rate, suggested that the microbial turnover was increasing and leading to a reduced efficiency of microbial protein [
12]. In this case, low feed intake might have occurred that led to animals′ severe nutrient deficiency.
Nutrient digestibility, especially the digestibility of protein, is essential—for both bacterial crude protein (BCP) and undegradable intake protein (UIP) during absorbing the metabolizable protein [
12]. In the present study, digestibility rates of DM, OM, total digestible nutrient (TDN), neutral detergent fiber (NDF) were lower, whereas digestibility of protein was higher when dietary crude protein increased (
p < 0.05,
Table 3). Although, the present results were in contrast with previous study [
11] that Murrah buffaloes showed the greater digestibility rates of DM, OM, and protein. Diets were consisted of berseem, wheat straw, and concentrate mixture without inclusion of urea. Previously, the inclusion of urea from 10 to 30 g/kg in the diet composition led to increase the digestibility rates both of OM and protein [
41]. To note, the present study provided dietary CP increases with simultaneously increasing urea levels in the range of 3.7–8.0 g/kg DM in the buffalo diets (
Table 1). This suggests that the presence of urea in bull animal diets might enhance deamination and modulate the apparent of protein; however, the exceeding urea supplementation of the offered diets might have had considerably impact on ureagenesis. A generous hepatic ureagenesis becomes indispensable for ruminants to refrain poisoning from absorbed ammonia [
42]. Furthermore, considering carbohydrate digestion (TDN) in the rumen could be the most accurate predictor of BCP synthesis [
12]. The present study expected to provide the protein and carbohydrate digestions, so that the provision of 6.6% protein, the inclusion of 5.5 g/kg urea and the availability of energy intake at 2.2 Mcal/kg DM in the diet (
Table 1) suggested the optimum level for buffaloes to obtain greater digestibility rates of protein and TDN.
3.3. Ruminal Fermentation and Blood Urea Nitrogen
Ruminal fermentation end-product (pH, NH
3-N, VFAs) and blood metabolite such as blood urea nitrogen (BUN) are crucial parameters to assess whether animals are meeting their nutrient requirement without reducing the animal responses [
7,
11,
35,
37,
43]. Increasing the protein content in Thai swamp buffalo diets enriched the concentration of NH
3-N in the rumen (
p < 0.05), but surprisingly did not affect the ruminal pH (
Table 4). The ruminal pH of Thai swamp buffaloes fed the diets in the present study was unvaried, as the provision of different protein contents and the pH rate values (6.7–7.1) were expected. The current result was similar to prior observations [
32,
40]. To note, inclusion of urea in diets of the present study suggested a similar finding by Wanapat, et al. [
44] who added urea to a buffalo diet in a range of 15–30 g/kg concentrate. The ruminal pH was in the normal range from 6.8 to 6.9, which provided the optimal rumen circumstance for the rumen host to grow and to ferment nutrient digestion, especially protein. Hence, it might be that the rumen-buffering capacity did not occurred, but that even the ruminal NH
3-N changed. Further studies are needed to better explain the regression between rumen-buffering capacity, pH, and ruminal NH
3-N on manipulating protein supply in buffalo diets.
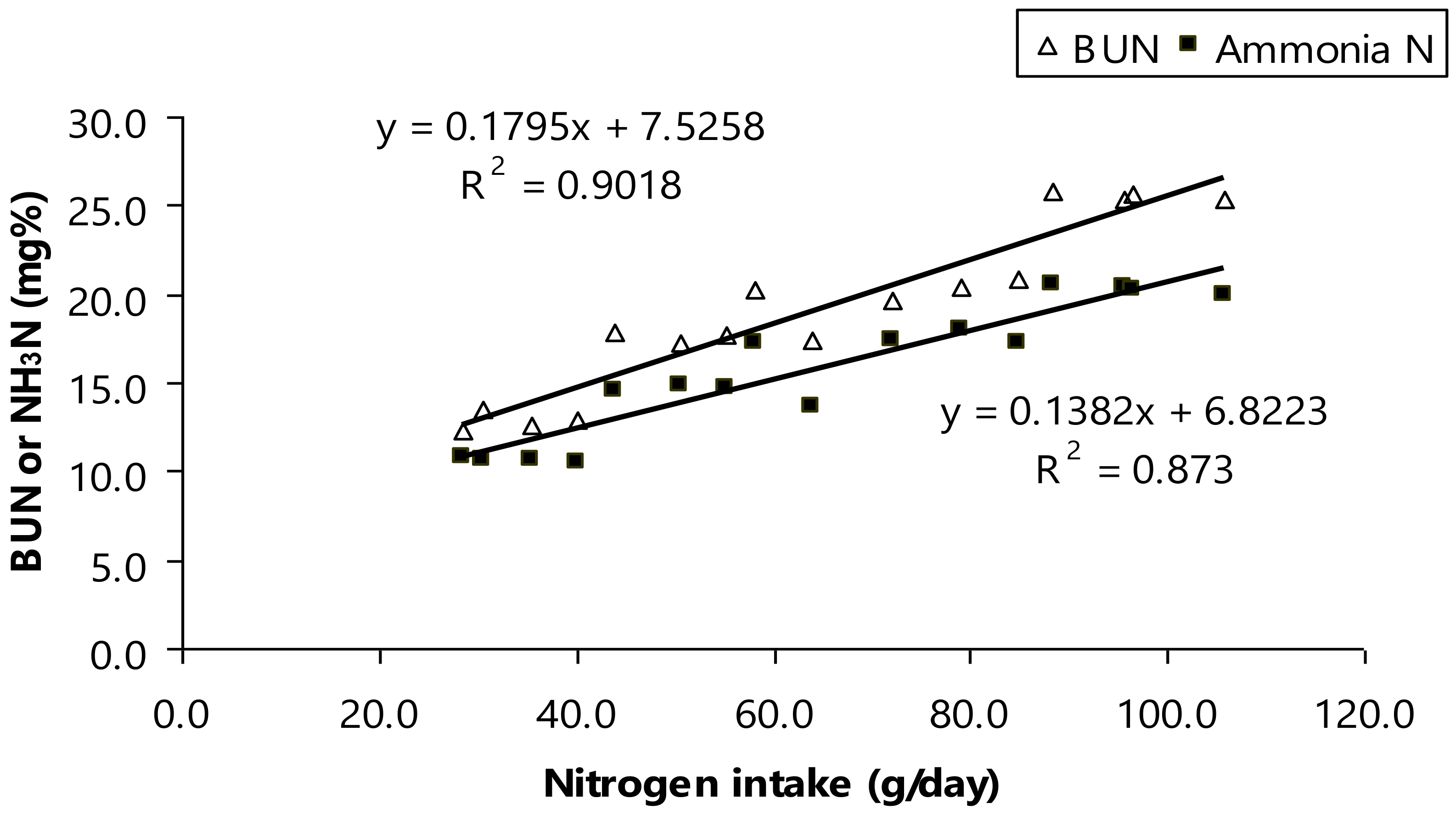
Furthermore, BUN concentration was shifted by dietary CP (
p < 0.01;
Table 4). As aforementioned a shift of ruminal NH
3-N, the mean of ruminal NH
3-N was at normal level (11.3–22.2 mg/dL) and considered safe for rumen host to avoid microbial turnover increasing [
45]. However, another study suggested that a shift of ruminal NH
3-N related to altered BUN concentration as modulating dietary CP for bull animals [
40,
46,
47]. Here, the mean of BUN in the present study was varied (
p < 0.01, 13.7–26.8 mg/dL) and those numbers were still considerably safe for ruminal bull animals [
40,
47]. Moreover, there was high relationship between nitrogen intake as varied protein intake, ruminal NH
3-N, and BUN (
Figure 2). Ruminal NH
3-N concentration increases when protein intake increases that relates to faster protein degradation than synthesis, higher dietary rumen-degradable protein (RDP) [
47], or an imbalance of fermentable energy [
41,
47,
48]. This occur suggesting that ammonia would be accumulated to rumen fluid and result in exceedingly concentration. As aforementioned, all of the ammonia including urea retention absorbed in lumen of the gut was removed by the liver with, as a result, a net splanchnic flux of zero to perform detoxification of ammonia by the liver [
42], and finally excreted via urine in to high levels of urine nitrogen [
40,
41].
Total VFA in the present study was varied in the range of 79.0–85.0 mM, and the VFA fraction including acetate and propionate was changed but the butyrate remained unchanged, where samples of rumen fluids observed at 0 and 4 h post-feedings by dietary CP (
Table 4). In general, the influences of the dietary CP occurred in the buffaloes fed the diet that consisted of 10.5% protein at 4 h post-feeding (
p < 0.05). A shift of VFAs relates to the ruminal host on digesting and metabolizing the nutrient source in diet, here, buffaloes could manifest their efficiency of feed utilization that is represented by increasing a number of VFAs. A higher VFAs might be due to a greater digestibility by VFA-producing bacteria. More CP intake at 10.5% showed the highest protein digestibility compared with other protein supplementations (
Table 3). However, those effects are time dependent. Since fermentable energy availability (TDN, g/kg W
0.75) was obtained in a similar pattern, the present results reflect that the dietary CP allowed the VFA-producing bacteria to rapidly accelerate the protein breakdown on nutrient digestion and the latter fermentable products were forwarded to hydrolysis and stored at a shift of VFAs as a main source of buffalo energy [
44,
47]. Additionally, there was a shift in VFAs produced in the rumen toward more propionate with corresponding to reduce in acetate and remain in butyrate (
Table 4). The acetate portion decreased when the propionate increased, indicating that the dietary CP in the present study altered the fermentable carbohydrate to be a major substrate for acetate fraction; however, a higher propionate suggested that propionate-producing bacteria was dominant to synthesize fermentable carbohydrate more propionate in gluconeogenesis by the pentose phosphate pathway produces a nicotinamide adenine dinucleotide phosphate [
18,
49,
50]. These occurrences might relate to the reduction of NDF digestion (
Table 3). The observed NDF digestion playing a role in the VFA-producing substrate is line with data by Vorlaphim, et al. [
51], who reported no changes for propionate portion in rumen fluids at 0 h, 2 h, and 4 h after feedings when NDF intake (g/d) was similar. It, thus, can be suggested that this declining availability of fermentable carbohydrate might relate to decrease acetate portion as the first major VFA absorbed from the rumen to have somewhat distinctive metabolic shift [
28,
49,
50]. This achievement was similar with the previous study [
32] and was expected in the present study because of the altered carbohydrate digestion.
3.4. Nitrogen Balance, Purine Derivative Excretion, Microbial-Related N Characteristic, and Rumen Microorganim Population
The provision of increased dietary CP in Thai swamp buffaloes increased the size of feces (
p < 0.05,
Table 5). This occurrence was expected due to the efficiency of dietary CP which was suggested, below 10% DM CP in the diet, as aforementioned [
32,
33,
34,
35]. Hence, the present results could be corroborating previous reports. This possibly occurred due to a higher dietary CP; the increasing fecal size might also be due to the likely enlarged DM intake and slightly dwindled number of DM digestibility (
Table 3), so as to excrete lowered undigested dietary CP formed in a higher volume of feces. Therefore, a significant difference in fecal N among diets in the present study could be attributed to the endogenous losses from digestive tracts; however, it might not be varied among bull animals [
15].
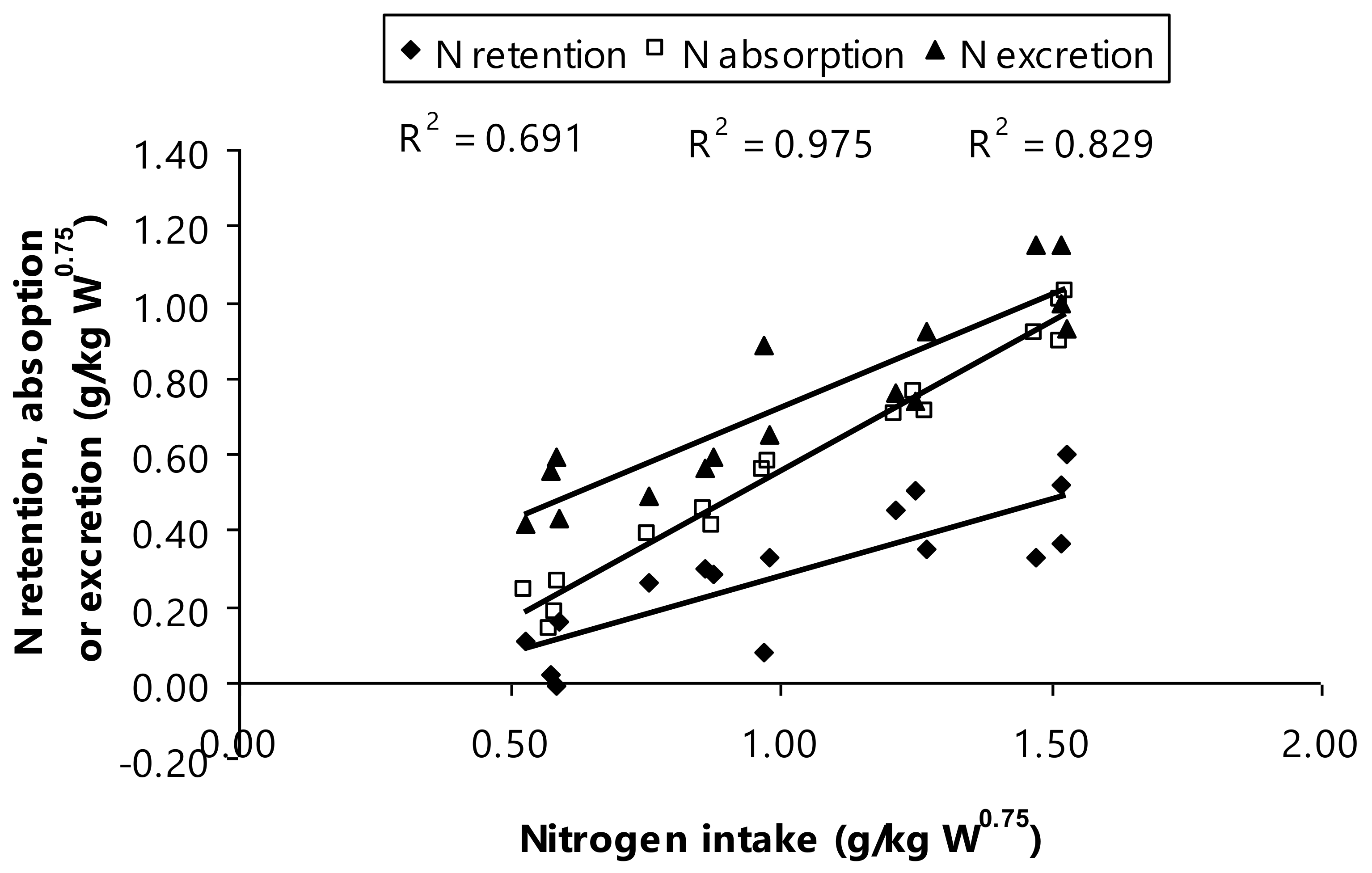
Concurrently, the current scenario resulted in N contents determined in N intake, N excretion, N absorption, and N retention as indicated a higher N amount of urinary and fecal (
p < 0.01). It has been well-documented that increasing protein content in diet inevitably increases N intake and this surpasses the numbers of N retention and N excretion in Thai-indigenous heifers [
9,
40], Murrah buffaloes [
11], and Nili-Ravi buffaloes [
46]. Notably, Thai swamp buffaloes fed to increased dietary CP as a depiction in the present study showing that there is relationship between N intake, N retention, and both of N absorption and excretion (g/kg W
0.75;
Figure 3). These recent findings interpreting that N retention was ranged from 0.07 to 0.45 g N/kg W
0.75 and N absorption ranged from 0.21 to 0.96 g N/kg W
0.75, when the Thai swamp buffaloes consumed the increased dietary CP levels. This relationship gave evidence that the provision of increased dietary CP among bull animals resulted in an increase in the N amount in feces, nitrogen fecal, and urine, if the fermentable energy availability was insufficient supply.
It has been discussed that N excretion via urine is increased with an increasing protein level in diet (N intake), which may relate to the RDP ration in dietary CP [
41,
52]. To note, high RDP ratios in dietary CP have been showing due to the increase of N in urine, but this occur led to a decrease N balance [
46]. The utilized nitrogen parameter such as nitrogen or protein degradability has a main effect on urinary N output because of excess soluble N in the rumen from diets with high RDP [
41]. Several factors, namely N origin, degree of N intake, degree of energy intake, age, sex, and physiological animal related to metabolic perspective could be considered as profoundly influencer on shifting nitrogen balance [
53]. Nitrogen balance defines the animal’s status to meet their protein requirement, as metabolized protein excess (supply minus requirements; NRC [
12]). However, excessively N retaining no longer gives a metabolic benefit due to those N absorbed from the lumen of the gut into the portal vein, which must be partially removed by the liver to avoid a large hepatic ureagenesis as discussed above [
42].
The urinary purine derivative (PD) excretion (mmol/d or µmol/kg W
0.75) increased when dietary CP in Thai swamp buffaloes increased (
p < 0.05;
Table 6). Concentration of PD parameters (allantoin, uric acid, xanthine, and hypoxanthine) in the present study was in the range of 16.3–31.3 mmol/d and it was expected to be a similar range as reported in other buffalo investigations [
26,
54]. In addition, there was a significant increase (
p < 0.05) in allantoin and total PD, and non-significant responses or remaining unchanged in relatively uric acid, xanthine and hypoxanthine excretions that observed in urine as influence of increased dietary CP. These achievements in the present study were similar to previous studies [
26,
41], which sizes of allantoin and uric acid excretions were relatively normal for bull animals as influenced protein content in diets. Moreover, the present study by adjusting dietary CP seemed to have no influence on urinary excretion of creatinine (µmol/kg W
0.75). However, dietary CP shifted ratio of A:C, PD:C and PDC index (
p < 0.01;
Table 6). Creatinine excretion might be influenced by the amount of protein intake by animals and the present study showed increased pattern of DM intake as well as concomitantly protein content in diets increased (
Table 1 and
Table 3). However, the recent findings were not in line with previous studies that were observed in Zebu cattle and swamp buffaloes [
55], and Murrah buffaloes [
56]. To compare, those bull animals had been to show the difference of creatinine excretions due to the marginal variations among studies. Variation of animals, diets, experimental managements were expected to differ the characteristic and profile in creatinine shift [
55]. We speculated also that breed or species specific and more closely correlated with muscle mass than body weight might differ the creatinine size, and not depend on dietary intake. Thus, the present study could be assumed to reconfirm that the dietary intake of animals did not attribute to vary the creatinine excretion, if those fed to similar in diet concern and group of selected breeds.
To determine and predict the protein requirement for the maintenance and growth of Thai swamp buffaloes on any given feeding program, it is necessary to establish the characteristic of microbial nitrogen supply and microbial nitrogen efficiency. Adjusting dietary CP was expected to increase linearly microbial purine base (MPB) flow (mmol/d), microbial N supply (g N/d), and microbial N efficiency (g N/kg nutrient utilization) in urinary Thai swamp buffaloes (
p < 0.01;
Table 6). However, a solely microbial N efficiency in terms of CP intake remained unchanged. Our results were in agreement with the previous study of Paengkoum et al. [
41]; their group assessed that microbial N synthesis grew up by adding urea from 10 to 30 g/kg steam-treated oil palm fronds. As mentioned above, urea stimulated to increase protein availability in rumen. Kim et al. [
47] reported rumen richer in fermentable protein derived from dietary CP (9.2 vs. 11.2%DM) and solely RDP fraction (52.3 vs. 79.8%CP) resulting in plentiful microbial N supply to omasum. It may indicate microbial N turnover decreasing due to a greater N source. Exceedingly high CP supplementation at 13.5 to 19.4% DM had been reported to show an inclination of omasal flow of total non-NH
3-N bacteria from 425 to 480 g/d [
57]. More efficiency gains had been achieved by only increasing RDP level (10.6–13.2%) in the diets tending to enrich total non-NH
3-N bacteria [
58]. It could be assessed that impressive numbers of total non-NH
3-N bacteria flowing from rumen to omasum due to dietary CP increased as protein intake and its degradability increased, but this efficiency of microbial protein synthesis had seemed to depend on the age of rumen itself. The age-related succession of rumen microbial communities, especially
Bacteroidetes and
Proteobacteria populations, defined the shift of ruminant animal productivity as growing higher in presence of older age rumen [
59]. In other words, animal age might have influence on differing microbial efficiency. Moreover, the present study had provided the experimental design in numerous levels of CP and considerably energy supply. This was prepared because we still assumed that any protein catabolism converting to amino acid seemed to have sufficient fermentable energy availability. The traditional meta-analysis regarding microbial efficiency in rumen had been previously discussed to relate protein synthesis is based on ruminal carbohydrate as a main energy source in rumen [
13]. The high-energy and low-protein diets shifted the available N for microbial growth in limited microbial protein synthesis [
60]. Of note, therefore, several factors such as the availability of carbohydrates and N in rumen, ruminal pH, physiological effects, sources and levels of N components, and other stabilizing ruminal fermentations had been discussed to have a substantial role on modifying the efficiency of microbial protein in rumen [
13,
60,
61].
Despite similar pH in rumen, dietary CP increased relatively fungal zoospore and total bacteria in rumen of Thai swamp buffaloes (
p < 0.05;
Table 7). Cellulolytic bacterial population had quadratic and cubic effects (
p < 0.05). Amylolytic bacterial population had a linear effect (
p < 0.05). However, increasing CP content in diets did not affect proteolytic bacterial population in the rumen of Thai swamp buffaloes neither observed in 0 and 4 h post-feedings. The inclination of increased numbers in those recently mentioned findings were similar patterns as a well-documented in those prior reports [
62,
63]. There may be a big question mark in which the proteolytic bacterial population did not get affection by adjusting dietary CP, while the digestibility of CP (
Table 3) was a greater appearance. The possibly reason might relate to the protein metabolism itself in rumen as present objective delivered. Protein metabolism in the rumen is metabolically manifested by activity of ruminal microorganisms to have a corresponding effect towards the nutrient utilization. Here, protein utilization is depending on the structure of nutrient and it is defining whether having a susceptibility to microbial proteases and, thus, its degradability. Bach, et al. [
64] had pointed out several findings by reviewing the nitrogen metabolism in the rumen. Ruminal protein degradation is shifted by pH and the major microbial population. Proteolytic population represents the protein degrader in rumen and its activity changes as pH changes with high-forage dairy cattle-type rations, but not in high-concentrate beef-type rations. It, thus, indicating that proteolytic population degrades the protein substrates in post-feeding and finally shifts to the amino acid in rumen. However, some amino acid, such as Ile, Leu, and Phe which are synthesized by rumen microorganisms tend to limit the protein degradation in rumen [
9,
53,
64]. The second reason pointing to a lack of determining proteolytic population was in a recent alternative and complementary technique of the present study, where the measurement of proteolytic counting used the roll-tube technique observed in rumen culture that is no longer accurate. It was recently suggested that bacterial cells, including N distribution therein, could be affected by indigenously factor in rumen such as rate of fermentation [
64]. As consequence, further studies are needed to greater explain the protein utilization by proteolytic population in rumen determined by using the sequence technique to define extracellular prokaryotic diversity in bovine rumen [
65].
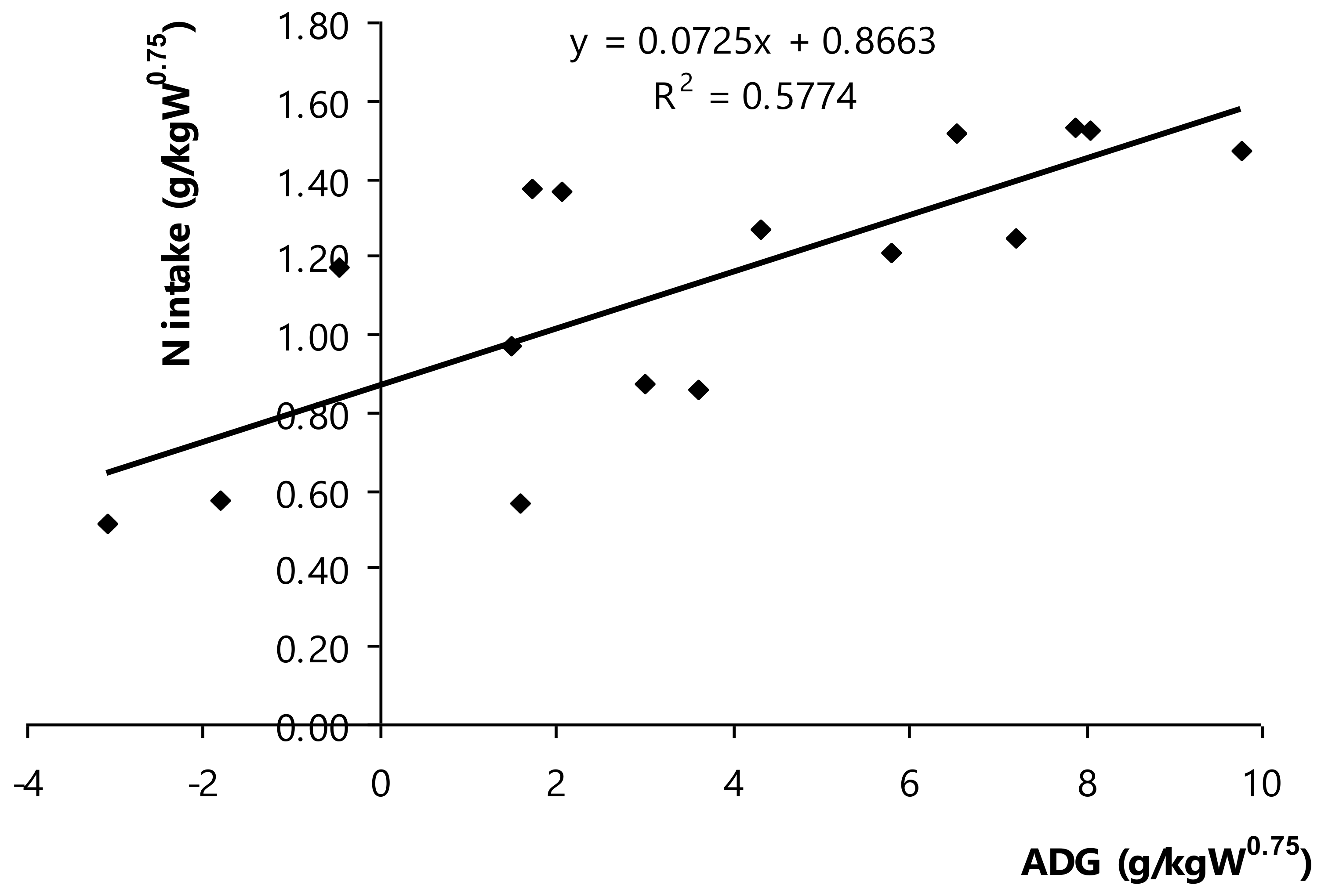
3.5. Nitrogen or Crude Protein Requirement
The net nitrogen or crude protein requirements for maintenance and growth of Thai swamp buffaloes were determined based on ADG (g ADG/kg W
0.75) and N intake (g N/kg W
0.75;
Figure 4). It depicted the linear regression between dietary nitrogen or crude protein and the net requirements for maintenance and growth of Thai swamp buffaloes, as equation = 0.0725ADG + 0.8663 (R
2 = 0.577,
p < 0.001,
n = 16). Nitrogen requirement could be estimated, the nitrogen intake in which ADG (equally at 0) was nitrogen requirement for maintenance, ADG index (at slope) was nitrogen requirement for growth of Thai swamp buffaloes. Of note, it could be mathematically calculated following to 0.866 g N/kg W
0.75 or equivalent to 5.41 g CP/kg W
0.75/d (maintenance requirement), and 0.073 g N/g ADG or equivalent to 0.46 g CP/g ADG/d (growth). The recently findings regard of the net CP requirement for growth of Thai swamp buffaloes was higher than it reported in NRC [
12], but the maintenance was lower. Besides, the present results were slightly higher for maintenance of buffaloes and lower to roughly at 24.0% for growth of buffaloes compared to those reported in Kearl′s report (maintenance, (5.24 g CP/kg W
0.75; growth, 0.65 g CP/g ADG), as recommendation CP requirements of ruminants in developing countries [
15]. It is possible that these differences were at the UIP presence of the applied diet as a deciding factor to have influence on the unaffected nutrient apparent tract digestibility and the population of ammonia-producer bacteria. Rumen fermentation alleviated with the greater number of UIP in diet, but UIP intensified ammonia retention as higher efficiency of nitrogen utilization in rumen. For instance, Paengkoum et al. [
9] had estimated the metabolized protein (MP) requirement to increase one g/kg BW
0.75 that was 0.34 g MP/kg BW
0.75 of Thai-indigenous growing cattle and MP requirement for maintenance was 2.77 g/kg BW
0.75. It was suggested to supplement not over 10.0% CP DM in diet with proportional ratio of UIP and degradable intake protein (DIP, 35:65), which resulted in the optimum growth performance for growing Thai-indigenous beef cattle. Several previous studies also had been testing to prove the net nitrogen requirement for maintenance and growth among bull animals having a age-related succession of rumen microbial communities [
59], breed and sex [
35,
37], and balanced content of protein and energy [
7,
56]. Collectively, the present findings could corroborate other possibly factors, protein requirement of bull animals might relate to specific domestication and climatic condition as Thai swamp buffaloes investigated in the present study.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}