
Effects of L-Histidine and Sodium Acetate on β-Casein Expression in Nutrient-Restricted Bovine Mammary Epithelial Cells


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Experimental Design
2.3. Cell Viability
2.4. Protein Quantification
2.5. Gene Expression
2.6. Statistical Analysis
3. Results
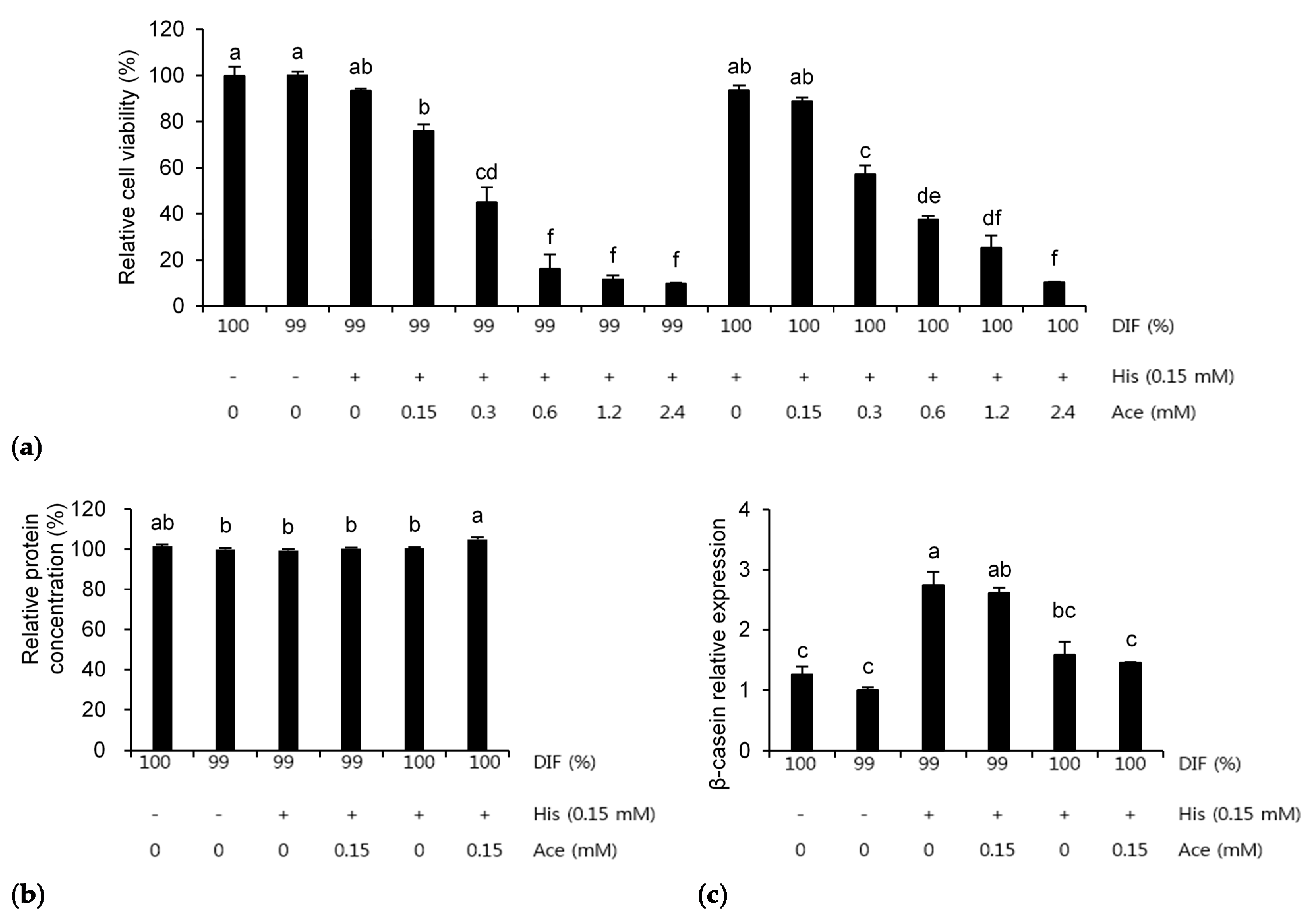
3.1. Effect of NR on MAC-T Cell Viability
3.2. Individual Effects of His and Ace under the NR Condition
3.3. Combination Effects of His and Ace in the NR Condition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.G.; Buel, G.R.; Blenis, J. Nutrient regulation of the mTOR complex 1 signaling pathway. Mol. Cells 2013, 35, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Appuhamy, J.A.; Nayananjalie, W.A.; England, E.M.; Gerrard, D.E.; Akers, R.M.; Hanigan, M.D. Effects of AMP-activated protein kinase (AMPK) signaling and essential amino acids on mammalian target of rapamycin (mTOR) signaling and protein synthesis rates in mammary cells. J. Dairy Sci. 2014, 97, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhou, Z.; Wang, L.; Saremi, B.; Helmbrecht, A.; Wang, Z.; Loor, J. Increasing the availability of threonine, isoleucine, valine, and leucine relative to lysine while maintaining an ideal ratio of lysine: Methionine alters mammary cellular metabolites, mammalian target of rapamycin signaling, and gene transcription. J. Dairy Sci. 2018, 101, 5502–5514. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Zhao, S.; Zheng, N.; Zhang, Y.; Wang, S.; Zhou, X.; Wang, J. Combination of histidine, lysine, methionine, and leucine promotes β-casein synthesis via the mechanistic target of rapamycin signaling pathway in bovine mammary epithelial cells. J. Dairy Sci. 2017, 100, 7696–7709. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine mammary protein synthesis during the lactation cycle. Bioinform. Biol. Insights 2011, 5, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, C.; Yoshino, K.; Yonezawa, K. mTOR integrates amino acid- and energy-sensing pathways. Biochem. Biophys. Res. Commun. 2004, 313, 443–446. [Google Scholar] [CrossRef]
- Wang, X.; Proud, C.G. The mTOR pathway in the control of protein synthesis. Physiology 2006, 21, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Broderick, G.A.; Stevenson, M.J.; Patton, R.A.; Lobos, N.E.; Olmos Colmenero, J.J. Effect of supplementing rumen-protected methionine on production and nitrogen excretion in lactating dairy cows. J. Dairy Sci. 2008, 91, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Giallongo, F.; Hristov, A.N.; Oh, J.; Frederick, T.; Weeks, H.; Werner, J.; Lapierre, H.; Patton, R.A.; Gehman, A.; Parys, C. Effects of slow-release urea and rumen-protected methionine and histidine on performance of dairy cows. J. Dairy Sci. 2015, 98, 3292–3308. [Google Scholar] [CrossRef]
- Lee, C.; Hristov, A.N.; Cassidy, T.W.; Heyler, K.S.; Lapierre, H.; Varga, G.A.; de Veth, M.J.; Patton, R.A.; Parys, C. Rumen-protected lysine, methionine, and histidine increase milk protein yield in dairy cows fed a metabolizable protein-deficient diet. J. Dairy Sci. 2012, 95, 6042–6056. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.N.; Hu, H.; Zheng, N.; Wang, J.Q. Leucine and histidine independently regulate milk protein synthesis in bovine mammary epithelial cells via mTOR signaling pathway. J. Zhejiang Univ. Sci. B 2015, 16, 560–572. [Google Scholar] [CrossRef]
- Sheng, R.; Yan, S.; Qi, L.; Zhao, Y.; Jin, L.; Guo, X. Effect of the ratios of acetate and β-hydroxybutyrate on the expression of milk fat-and protein-related genes in bovine mammary epithelial cells. Czech J. Anim. Sci. 2015, 60, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yan, S.; Chen, L.; Shi, B.; Guo, X. Effect of interaction between leucine and acetate on the milk protein synthesis in bovine mammary epithelial cells. Anim. Sci. J. 2019, 90, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, J.-E.; Lee, J.-S.; Park, J.-S.; Moon, J.-O.; Lee, H.-G. Phenylalanine and valine differentially stimulate milk protein synthetic and energy-mediated pathway in immortalized bovine mammary epithelial cells. J. Anim. Sci. Technol. 2020, 62, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Jeon, S.W.; Jung, U.S.; Kim, M.J.; Lee, H.G. l-Lactate Dehydrogenase B Chain Associated with Milk Protein Content in Dairy Cows. Animals 2019, 9, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Lim, J.N.; Bok, J.D.; Kim, J.H.; Kang, S.K.; Lee, S.B.; Hwang, J.H.; Lee, K.H.; Kang, H.S.; Choi, Y.J.; et al. Association of protein expression in isolated milk epithelial cells and cis-9, trans-11 conjugated linoleic acid proportions in milk from dairy cows. J. Sci. Food Agric. 2014, 94, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Loor, J.J. Identification of reference genes for quantitative real-time PCR in the bovine mammary gland during the lactation cycle. Physiol. Genom. 2007, 29, 312–319. [Google Scholar] [CrossRef]
- Jedrzejczak, M.; Szatkowska, I. Bovine mammary epithelial cell cultures for the study of mammary gland functions. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Dessauge, F.; Lollivier, V.; Ponchon, B.; Bruckmaier, R.; Finot, L.; Wiart, S.; Cutullic, E.; Disenhaus, C.; Barbey, S.; Boutinaud, M. Effects of nutrient restriction on mammary cell turnover and mammary gland remodeling in lactating dairy cows. J. Dairy Sci. 2011, 94, 4623–4635. [Google Scholar] [CrossRef] [Green Version]
- Herve, L.; Quesnel, H.; Veron, M.; Portanguen, J.; Gross, J.J.; Bruckmaier, R.M.; Boutinaud, M. Milk yield loss in response to feed restriction is associated with mammary epithelial cell exfoliation in dairy cows. J. Dairy Sci. 2019, 102, 2670–2685. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Zhao, F.; Liu, J.; Liu, H. ASCT2 is involved in SARS-mediated β-casein synthesis of bovine mammary epithelial cells with methionine supply. J. Agric. Food Chem. 2019, 68, 13038–13045. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, S.; Zou, Y.; Zhao, F.Q.; Liu, J.; Liu, H. Lysine Stimulates Protein Synthesis by Promoting the Expression of ATB0,+ and Activating the mTOR Pathway in Bovine Mammary Epithelial Cells. J. Nutr. 2018, 148, 1426–1433. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Guo, X.; Yan, S.; Shi, B.; Sheng, R. Acetate regulates milk fat synthesis through the mammalian target of rapamycin/eukaryotic initiation factor 4E signaling pathway in bovine mammary epithelial cells. J. Dairy Sci. 2021, 104, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Xiao, C.; Guo, C.; Zhang, X.; Wang, Y.; Wang, J.; Yang, Z.; Fu, Y. Sodium acetate inhibits Staphylococcus aureus internalization into bovine mammary epithelial cells by inhibiting NF-κB activation. Microb. Pathog. 2017, 107, 116–121. [Google Scholar] [CrossRef]
- Jeon, S.W.; Conejos, J.R.; Kim, J.; Kim, M.J.; Lee, J.E.; Lee, B.S.; Park, J.S.; Moon, J.O.; Lee, J.S.; Lee, H.G. Supplementing conjugated and non-conjugated L-methionine and acetate alters expression patterns of CSN2, proteins and metabolites related to protein synthesis in bovine mammary cells. J. Dairy Res. 2020, 87, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Q. Biology of glucose transport in the mammary gland. J. Mammary Gland Biol. Neoplasia 2014, 19, 3–17. [Google Scholar] [CrossRef]
- Zhao, F.Q.; Keating, A.F. Expression and regulation of glucose transporters in the bovine mammary gland. J. Dairy Sci. 2007, 90 (Suppl. 1), E76–E86. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Sun, X.; Hou, X.; Qu, B.; Gao, X.; Li, Q. Effects of glucose on lactose synthesis in mammary epithelial cells from dairy cow. BMC Vet. Res. 2016, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Boutinaud, M.; Ben Chedly, M.H.; Delamaire, E.; Guinard-Flament, J. Milking and feed restriction regulate transcripts of mammary epithelial cells purified from milk. J. Dairy Sci. 2008, 91, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.C.; Zhao, S.G.; Wang, S.S.; Luo, C.C.; Gao, H.N.; Zheng, N.; Wang, J.Q. d-Glucose and amino acid deficiency inhibits casein synthesis through JAK2/STAT5 and AMPK/mTOR signaling pathways in mammary epithelial cells of dairy cows. J. Dairy Sci. 2018, 101, 1737–1746. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, Z.; Peng, J.; Loor, J.J. Methionine and valine activate the mammalian target of rapamycin complex 1 pathway through heterodimeric amino acid taste receptor (TAS1R1/TAS1R3) and intracellular Ca2+ in bovine mammary epithelial cells. J. Dairy Sci. 2018, 101, 11354–11363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Chen, Y.; Cortes, I.M.; Coleman, D.N.; Dai, H.; Liang, Y.; Parys, C.; Fernandez, C.; Wang, M.; Loor, J.J. Supply of methionine and arginine alters phosphorylation of mechanistic target of rapamycin (mTOR), circadian clock proteins, and alpha-s1-casein abundance in bovine mammary epithelial cells. Food Funct. 2020, 11, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Salama, A.A.K.; Duque, M.; Wang, L.; Shahzad, K.; Olivera, M.; Loor, J.J. Enhanced supply of methionine or arginine alters mechanistic target of rapamycin signaling proteins, messenger RNA, and microRNA abundance in heat-stressed bovine mammary epithelial cells in vitro. J. Dairy Sci. 2019, 102, 2469–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; van Baal, J.; Ma, L.; Loor, J.J.; Wu, Z.L.; Dijkstra, J.; Bu, D.P. Short communication: Relationship between lysine/methionine ratios and glucose levels and their effects on casein synthesis via activation of the mechanistic target of rapamycin signaling pathway in bovine mammary epithelial cells. J. Dairy Sci. 2019, 102, 8127–8133. [Google Scholar] [CrossRef] [PubMed]
- Prizant, R.L.; Barash, I. Negative effects of the amino acids Lys, His, and Thr on S6K1 phosphorylation in mammary epithelial cells. J. Cell. Biochem. 2008, 105, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Loor, J.J.; Liu, H.Y.; Liu, L.; Hosseini, A.; Zhao, W.S.; Liu, J.X. Optimal ratios of essential amino acids stimulate beta-casein synthesis via activation of the mammalian target of rapamycin signaling pathway in MAC-T cells and bovine mammary tissue explants. J. Dairy Sci. 2017, 100, 6676–6688. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Accession No. | Type 2 | Sequence (5′-3′) |
|---|---|---|---|
| ASCT2 | NM_174601.2 | F | TGCCGCTGATGATGAAGTGT |
| R | AGTCCACGGCCAAGATCAAG | ||
| GLUT1 | NM_174602.2 | F | TCGCTTCATCATCGGTGTGT |
| R | GCTTCTTCAGCACGCTCTTG | ||
| LAT1 | AF174615 | F | TACTTCCTTGGGGTCTGGTG |
| R | GTATCTGCGGACATCCACCT | ||
| mTOR | XM_002694043.6 | F | ATGCTGTCCCTGGTCCTTAT |
| R | GGGTCAGAGAGTGGCCTTCA | ||
| RPS6 | NM_001015548.2 | F | TGAAGCAGGGTGTCTTGACC |
| R | TCCAGTCCTCCTTGGTCTGT | ||
| S6K1 | NM_205816.1 | F | GGACATGGCAGGGGTGTTT |
| R | GGTATTTGCTCCTGTTACTT | ||
| UXT | NM_001037471.2 | F | GCGCTACGAGGCTTTCATCT |
| R | CCAAGGGCCACATAGATCCG | ||
| β-actin | NM_173979.3 | F | GCATGGAATCCTGCGGC |
| R | GTAGAGGTCCTTGCGGATGT | ||
| β-casein | XM_015471671.2 | F | GAGCCTGACTCTCACTGATGTTGAA |
| R | GACAGCACGGACTGAGGAGGAA |
| Treatment 1,2 | ||||||||
|---|---|---|---|---|---|---|---|---|
| 99%DIF + His | 100%DIF + His | |||||||
| Gene 3 | 99%DIF | 100%DIF | Ace − | Ace + | Ace − | Ace + | SEM 4 | p-Value |
| ASCT2 | 1.00 | 1.09 | 0.79 | 0.86 | 1.11 | 1.32 | 0.11 | 0.314 |
| LAT1 | 1.00 b | 1.39 b* | 0.86 b | 0.82 b | 1.02 b | 1.84 a# | 0.13 | 0.004 |
| GLUT1 | 1.00 | 1.45 * | 0.85 | 1.08 | 0.99 | 1.38 | 0.13 | 0.068 |
| mTOR | 1.00 | 1.05 | 0.87 | 0.93 | 0.97 | 1.16 | 0.06 | 0.582 |
| S6K1 | 1.00 | 0.99 | 0.98 | 1.02 | 0.96 | 1.38 # | 0.05 | 0.221 |
| RPS6 | 1.00 | 0.97 | 1.06 | 0.95 | 1.00 | 1.00 | 0.02 | 0.590 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Lee, H.-G. Effects of L-Histidine and Sodium Acetate on β-Casein Expression in Nutrient-Restricted Bovine Mammary Epithelial Cells. Animals 2021, 11, 1444. https://doi.org/10.3390/ani11051444
Kim J, Lee H-G. Effects of L-Histidine and Sodium Acetate on β-Casein Expression in Nutrient-Restricted Bovine Mammary Epithelial Cells. Animals. 2021; 11(5):1444. https://doi.org/10.3390/ani11051444
Chicago/Turabian StyleKim, Jungeun, and Hong-Gu Lee. 2021. "Effects of L-Histidine and Sodium Acetate on β-Casein Expression in Nutrient-Restricted Bovine Mammary Epithelial Cells" Animals 11, no. 5: 1444. https://doi.org/10.3390/ani11051444