
Dietary Synbiotics Can Help Relieve the Impacts of Deltamethrin Toxicity of Nile Tilapia Reared at Low Temperatures

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
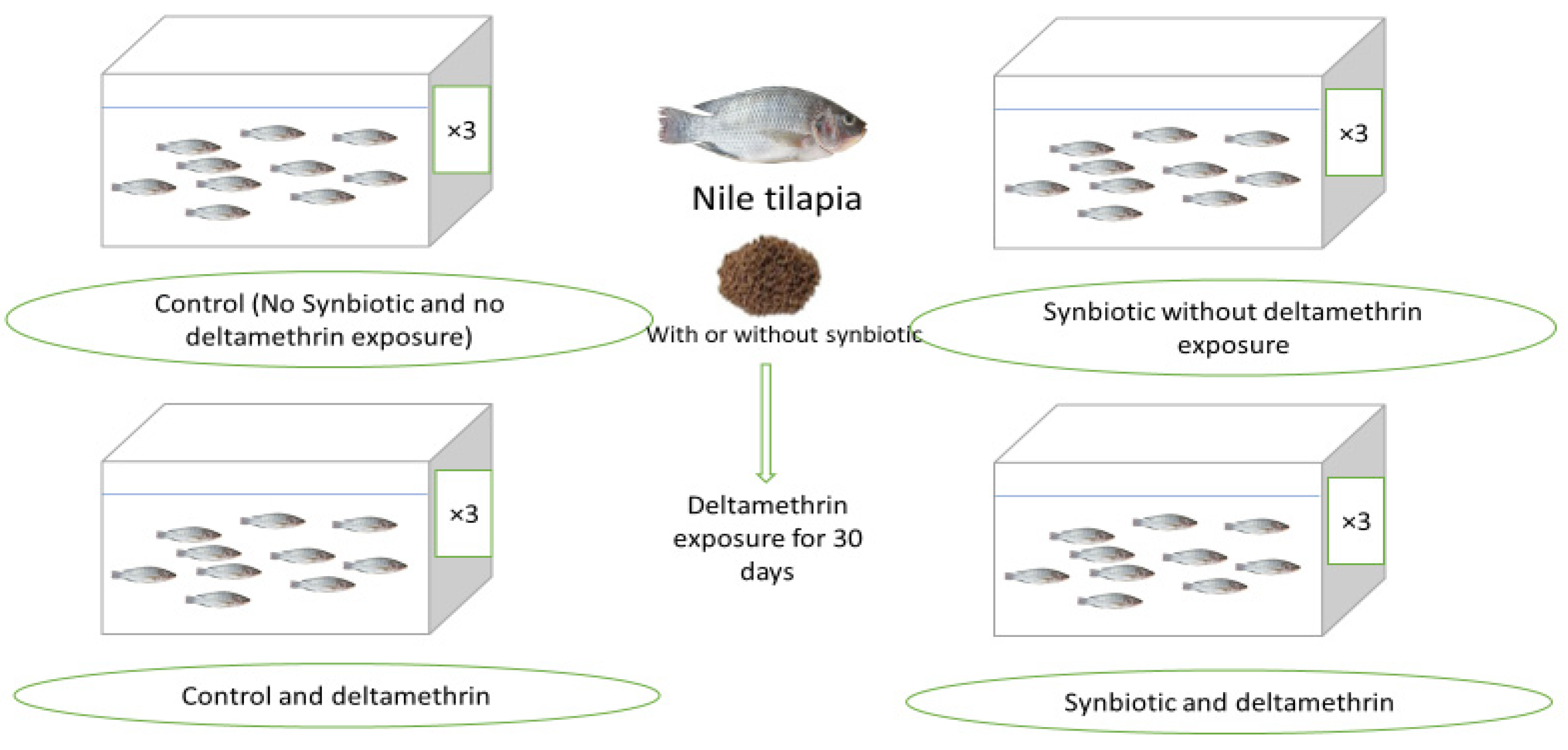
2.1. Fish, Diets, and Experimental Design
2.2. Histopathology Study
2.3. Transcriptome Assay
2.4. Statistical Analysis
3. Results
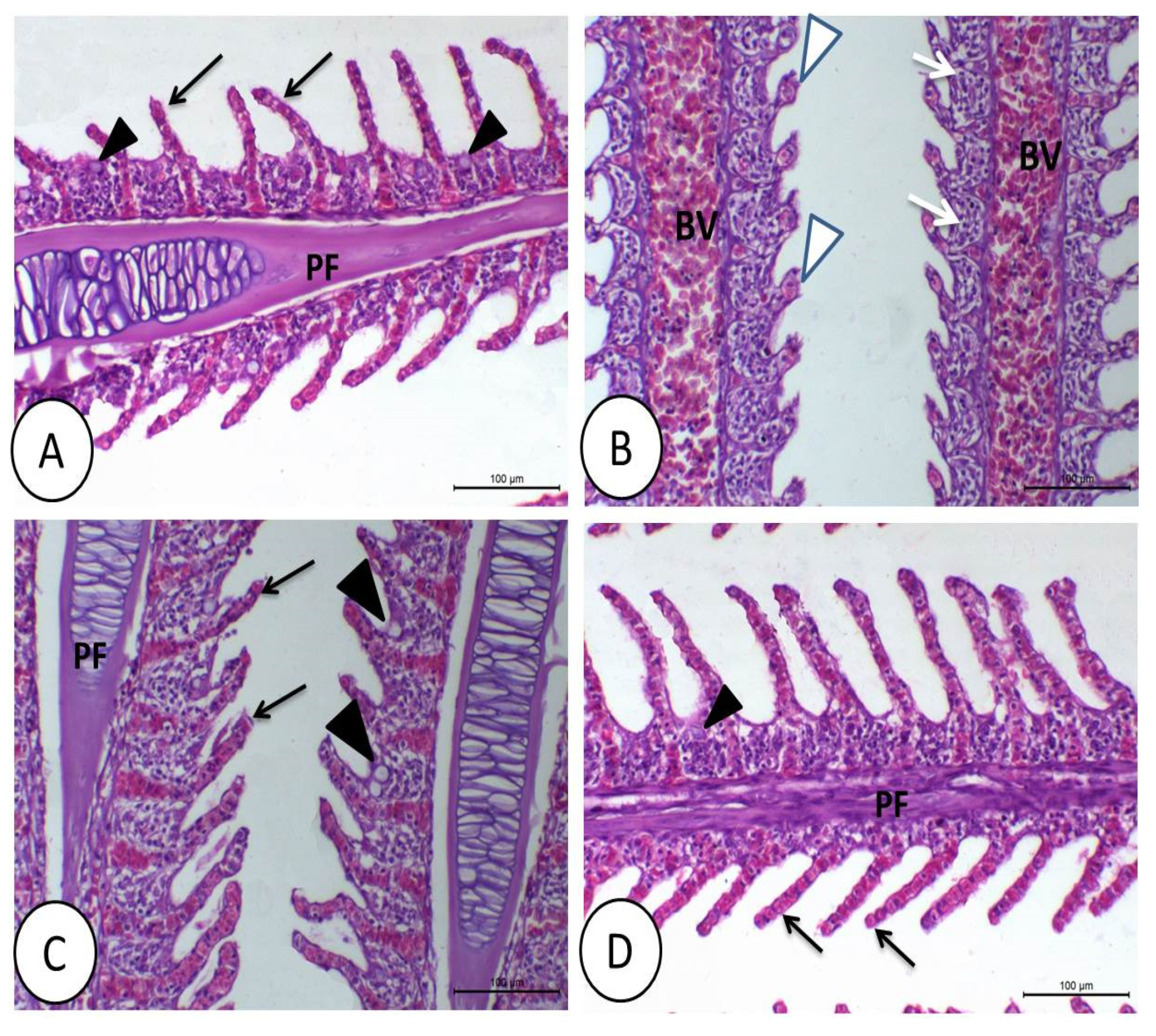
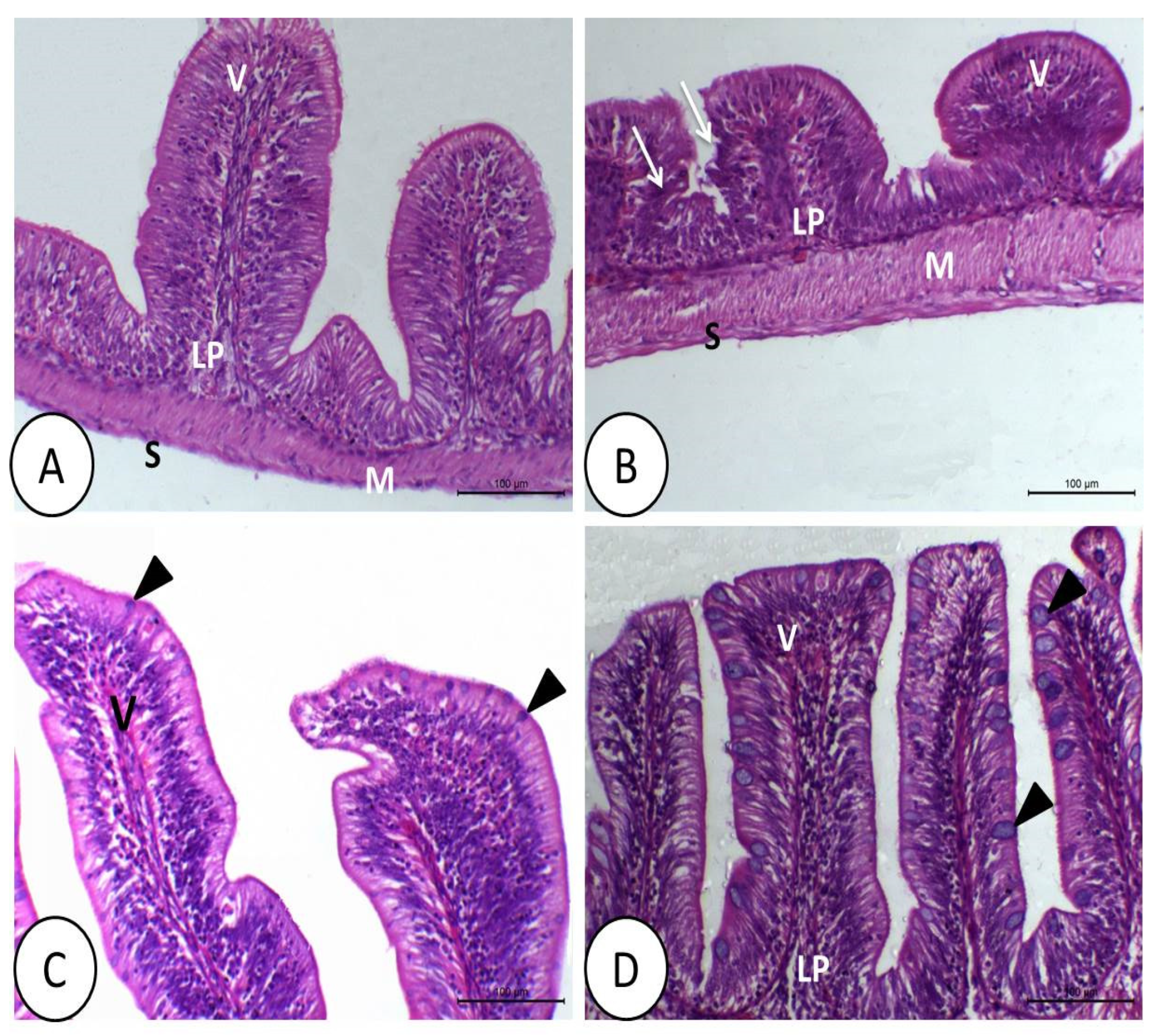
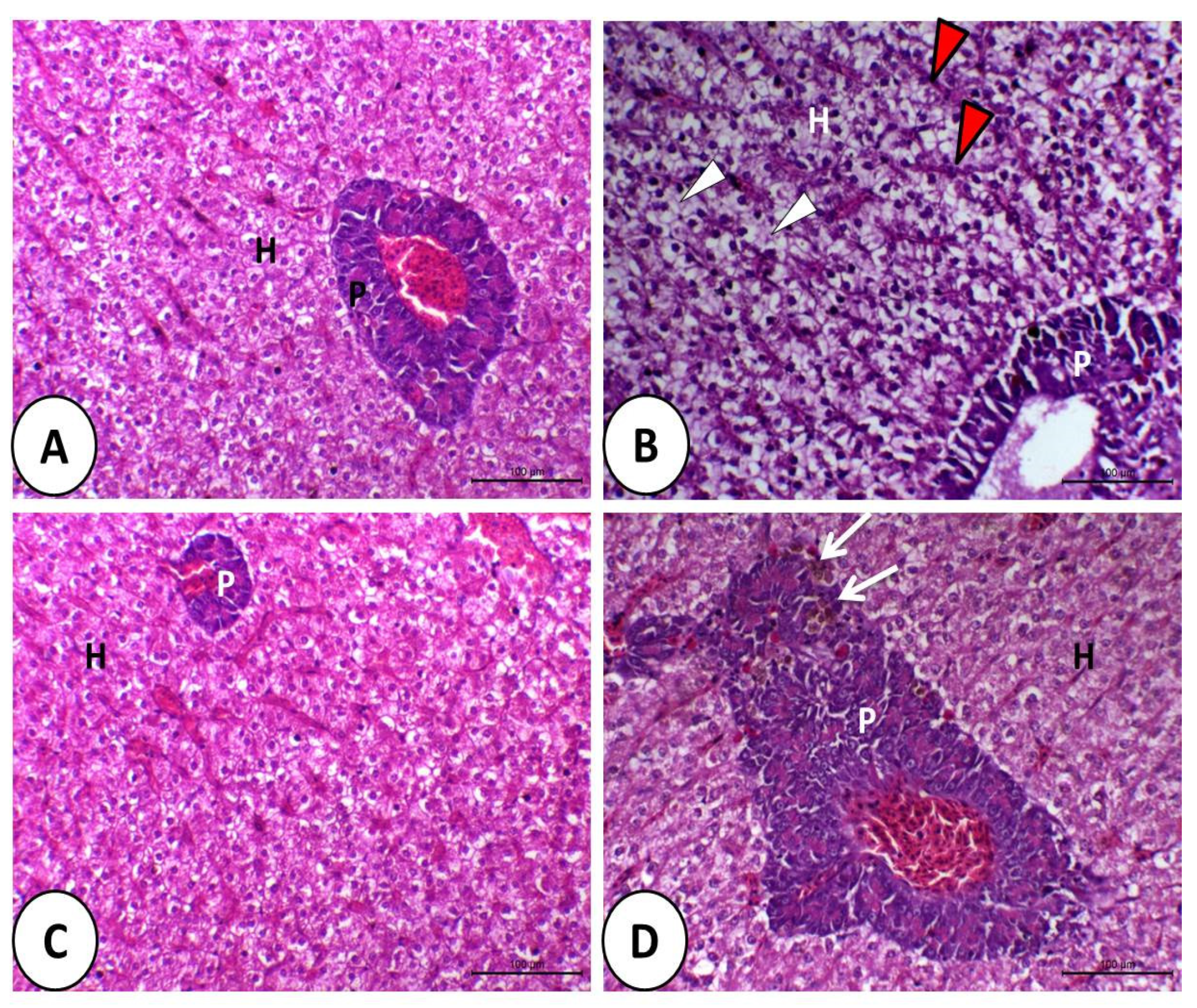
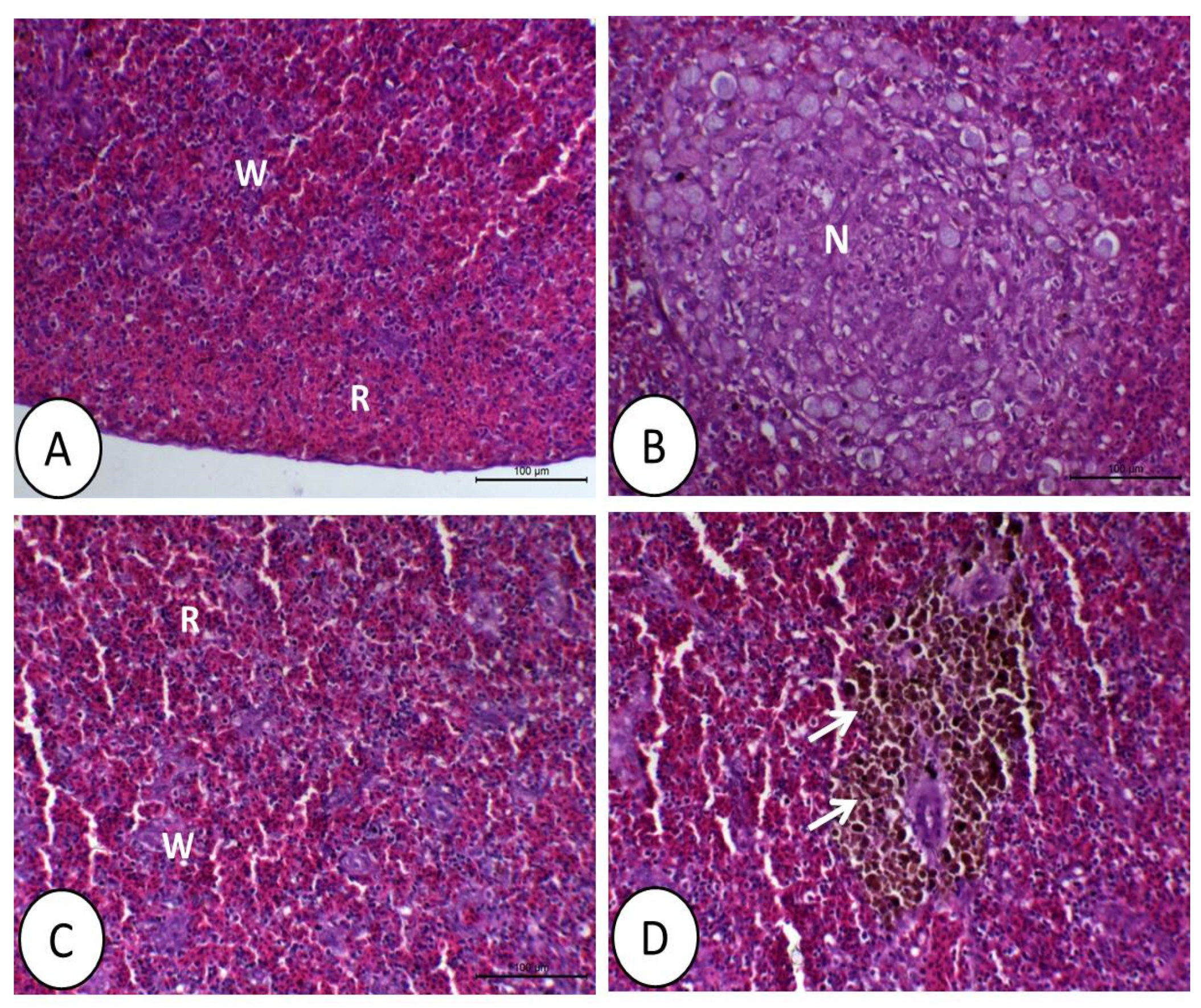
3.1. Histopathological Image
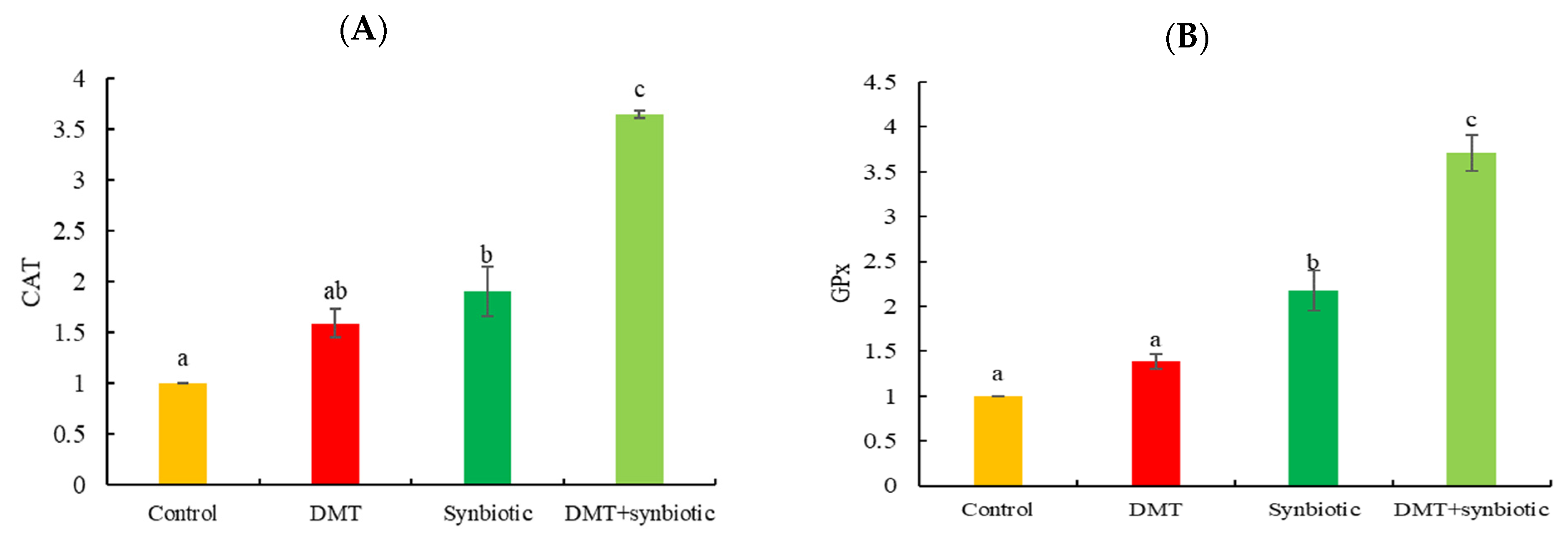
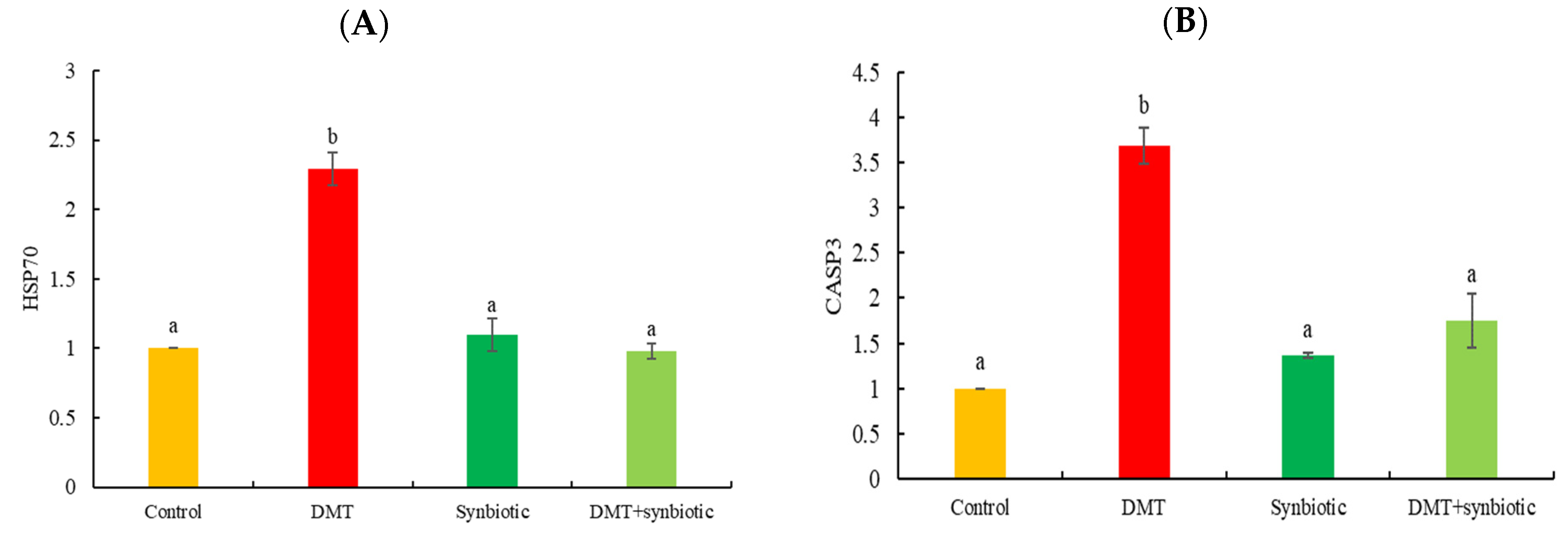
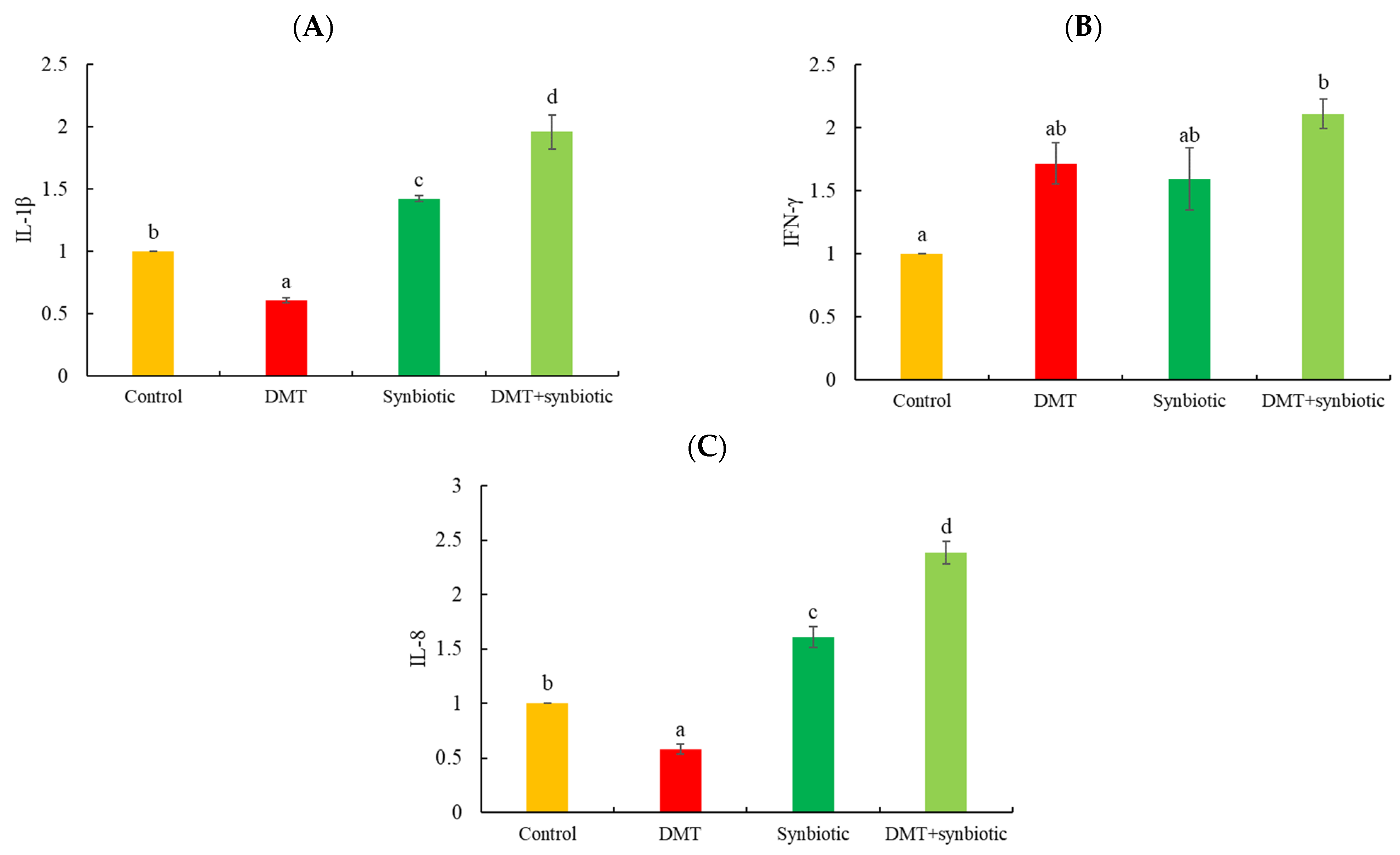
3.2. Gene Transcription
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmadifar, E.; Dawood, M.A.O.; Moghadam, M.S.; Sheikhzadeh, N.; Hoseinifar, S.H.; Musthafa, M.S. Modulation of immune parameters and antioxidant defense in zebrafish (Danio rerio) using dietary apple cider vinegar. Aquaculture 2019, 513, 734412. [Google Scholar] [CrossRef]
- El Megid, A.A.; Al Fatah, M.E.A.; El Asely, A.; El Senosi, Y.; Moustafa, M.M.A.; Dawood, M.A.O. Impact of pyrethroids and organochlorine pesticides residue on IGF-1 and CYP1A genes expression and muscle protein patterns of cultured Mugil capito. Ecotoxicol. Environ. Saf. 2020, 188, 109876. [Google Scholar] [CrossRef] [PubMed]
- Soliman, N.F.; Yacout, D.M.M. Aquaculture in Egypt: Status, constraints and potentials. Aquac. Int. 2016, 24, 1201–1227. [Google Scholar] [CrossRef]
- El-Murr, A.E.I.; El Hakim, Y.A.; Neamat-Allah, A.N.F.; Baeshen, M.; Ali, H.A. Immune-protective, antioxidant and relative genes expression impacts of β-glucan against fipronil toxicity in Nile tilapia, Oreochromis niloticus. Fish Shellfish. Immunol. 2019, 94, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Suvetha, L.; Saravanan, M.; Hur, J.-H.; Ramesh, M.; Krishnapriya, K. Acute and sublethal intoxication of deltamethrin in an Indian major carp, Labeo rohita: Hormonal and enzymological responses. J. Basic Appl. Zool. 2015, 72, 58–65. [Google Scholar] [CrossRef][Green Version]
- Dawood, M.A.O.; Moustafa, E.M.; Gewaily, M.S.; Abdo, S.E.; AbdEl-Kader, M.F.; SaadAllah, M.S.; Hamouda, A.H. Ameliorative effects of Lactobacillus plantarum L-137 on Nile tilapia (Oreochromis niloticus) exposed to deltamethrin toxicity in rearing water. Aquat. Toxicol. 2020, 219, 105377. [Google Scholar] [CrossRef]
- Datta, M.; Kaviraj, A. Ascorbic acid supplementation of diet for reduction of deltamethrin induced stress in freshwater catfish Clarias gariepinus. Chemosphere 2003, 53, 883–888. [Google Scholar] [CrossRef]
- Fanta, E.; Rios, F.S.A.; Romão, S.; Vianna, A.C.C.; Freiberger, S. Histopathology of the fish Corydoras paleatus contaminated with sublethal levels of organophosphorus in water and food. Ecotoxicol. Environ. Saf. 2003, 54, 119–130. [Google Scholar] [CrossRef]
- Bennour, E.E.G.; Timoumi, R.; Annaibi, E.; Mokni, M.; Omezzine, A.; Bacha, H.; Abid-Essefi, S. Protective effects of kefir against deltamethrin-induced hepatotoxicity in rats. Environ. Sci. Pollut. Res. 2019, 26, 18856–18865. [Google Scholar] [CrossRef]
- Jindal, R.; Sinha, R.; Brar, P. Evaluating the protective efficacy of Silybum marianum against deltamethrin induced hepatotoxicity in piscine model. Environ. Toxicol. Pharmacol. 2019, 66, 62–68. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abdo, S.E.; Gewaily, M.S.; Moustafa, E.M.; SaadAllah, M.S.; AbdEl-Kader, M.F.; Hamouda, A.H.; Omar, A.A.; Alwakeel, R.A. The influence of dietary β-glucan on immune, transcriptomic, inflammatory and histopathology disorders caused by deltamethrin toxicity in Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2020, 98, 301–311. [Google Scholar] [CrossRef]
- Srichaiyo, N.; Tongsiri, S.; Hoseinifar, S.H.; Dawood, M.A.O.; Esteban, M.Á.; Ringø, E.; Van Doan, H. The effect of fishwort (Houttuynia cordata) on skin mucosal, serum immunities, and growth performance of Nile tilapia. Fish Shellfish. Immunol. 2020, 98, 193–200. [Google Scholar] [CrossRef]
- Moustafa, E.M.; Dawood, M.A.O.; Assar, D.H.; Omar, A.A.; Elbialy, Z.I.; Farrag, F.A.; Shukry, M.; Zayed, M.M. Modulatory effects of fenugreek seeds powder on the histopathology, oxidative status, and immune related gene expression in Nile tilapia (Oreochromis niloticus) infected with Aeromonas hydrophila. Aquaculture 2019, 515, 734589. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish. Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Maradonna, F.; Gioacchini, G.; Falcinelli, S.; Bertotto, D.; Radaelli, G.; Olivotto, I.; Carnevali, O. Probiotic Supplementation Promotes Calcification in Danio rerio Larvae: A Molecular Study. PLoS ONE 2013, 8, e83155. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Bruni, L.; Randazzo, B.; Vargas, A.; Gioacchini, G.; Truzzi, C.; Annibaldi, A.; Riolo, P.; Parisi, G.; Cardinaletti, G.; et al. Partial Dietary Inclusion of Hermetia illucens (Black Soldier Fly) Full-Fat Prepupae in Zebrafish Feed: Biometric, Histological, Biochemical, and Molecular Implications. Zebrafish 2018, 15, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish. Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Modanloo, M.; Soltanian, S.; Akhlaghi, M.; Hoseinifar, S.H. The effects of single or combined administration of galactooligosaccharide and Pediococcus acidilactici on cutaneous mucus immune parameters, humoral immune responses and immune related genes expression in common carp (Cyprinus carpio) fingerlings. Fish Shellfish. Immunol. 2017, 70, 391–397. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J.; Tak, J.Y.; Lee, B.-J.; Kim, K.W.; Hur, S.W.; Han, H.-S.; Kim, B.-S.; Min, D.-H.; Kim, S.-K.; et al. Effects of Lactococcuslactis subsp. lactis I2 with Beta-Glucooligosaccharides on Growth, Innate Immunity and Streptococcosis Resistance in Olive Flounder (Paralichthys olivaceus). J. Microbiol. Biotechnol. 2018, 28, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Yu, R.; Zhang, Y.; Hu, B.; Jian, S.; Wen, C.; Kajbaf, K.; Kumar, V.; Yang, G. Effects of dietary supplementation with β-glucan and Bacillus subtilis on growth, fillet quality, immune capacity, and antioxidant status of Pengze crucian carp (Carassius auratus var. Pengze). Aquaculture 2019, 508, 106–112. [Google Scholar] [CrossRef]
- Devi, G.; Harikrishnan, R.; Paray, B.A.; Al-Sadoon, M.K.; Hoseinifar, S.H.; Balasundaram, C. Effect of symbiotic supplemented diet on innate-adaptive immune response, cytokine gene regulation and antioxidant property in Labeo rohita against Aeromonas hydrophila. Fish Shellfish. Immunol. 2019, 89, 687–700. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Ringø, E.; Masouleh, A.S.; Esteban, M.Á. Probiotic, prebiotic and synbiotic supplements in sturgeon aquaculture: A review. Rev. Aquac. 2016, 8, 89–102. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Choudhury, T.G.; Kamilya, D. Paraprobiotics: An aquaculture perspective. Rev. Aquac. 2019, 11, 1258–1270. [Google Scholar] [CrossRef]
- Singh, S.T.; Kamilya, D.; Kheti, B.; Bordoloi, B.; Parhi, J. Paraprobiotic preparation from Bacillus amyloliquefaciens FPTB16 modulates immune response and immune relevant gene expression in Catla catla (Hamilton, 1822). Fish Shellfish. Immunol. 2017, 66, 35–42. [Google Scholar] [CrossRef]
- Van Nguyen, N.; Onoda, S.; van Khanh, T.; Hai, P.D.; Trung, N.T.; Hoang, L.; Koshio, S. Evaluation of dietary Heat-killed Lactobacillus plantarum strain L-137 supplementation on growth performance, immunity and stress resistance of Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 498, 371–379. [Google Scholar] [CrossRef]
- Dawood, M.A.; Magouz, F.I.; Salem, M.F.; Abdel-Daim, H.A. Modulation of digestive enzyme activity, blood health, oxidative responses and growth-related gene expression in GIFT by heat-killed Lactobacillus plantarum (L-137). Aquaculture 2019, 505, 127–136. [Google Scholar] [CrossRef]
- Yassine, T.; Khalafalla, M.M.; Mamdouh, M.; Elbialy, Z.I.; Salah, A.S.; Ahmedou, A.; Mamoon, A.; El-Shehawi, A.M.; van Doan, H.; Dawood, M.A.O. The enhancement of the growth rate, intestinal health, expression of immune-related genes, and resistance against suboptimal water temperature in common carp (Cyprinus carpio) by dietary paraprobiotics. Aquac. Rep. 2021, 20, 100729. [Google Scholar] [CrossRef]
- Leyva-López, N.; Lizárraga-Velázquez, C.E.; Hernández, C.; Sánchez-Gutiérrez, E.Y. Exploitation of Agro-Industrial Waste as Potential Source of Bioactive Compounds for Aquaculture. Foods 2020, 9, 843. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Abdel-Kader, M.F.; Moustafa, E.M.; Gewaily, M.S.; Abdo, S.E. Growth performance and hemato-immunological responses of Nile tilapia (Oreochromis niloticus) exposed to deltamethrin and fed immunobiotics. Environ. Sci. Pollut. Res. 2020, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Magouz, F.I.; Salem, M.F.I.; Elbialy, Z.I.; Abdel-Daim, H.A. Synergetic Effects of Lactobacillus plantarum and β-Glucan on Digestive Enzyme Activity, Intestinal Morphology, Growth, Fatty Acid, and Glucose-Related Gene Expression of Genetically Improved Farmed Tilapia. Probiotics Antimicrob. Proteins 2019, 12, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Moustafa, E.M.; Elbialy, Z.I.; Farrag, F.; Lolo, E.E.E.; Abdel-Daim, H.A.; Abdel-Daim, M.M.; van Doan, H. Lactobacillus plantarum L-137 and/or β-glucan impacted the histopathological, antioxidant, immune-related genes and resistance of Nile tilapia (Oreochromis niloticus) against Aeromonas hydrophila. Res. Veter. Sci. 2020, 130, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Interaction effects of dietary supplementation of heat-killed Lactobacillus plantarum and β-glucan on growth performance, digestibility and immune response of juvenile red sea bream, Pagrus major. Fish Shellfish. Immunol. 2015, 45, 33–42. [Google Scholar] [CrossRef]
- Amin, A.; El Asely, A.; El-Naby, A.S.A.; Samir, F.; El-Ashram, A.; Sudhakaran, R.; Dawood, M.A.O. Growth performance, intestinal histomorphology and growth-related gene expression in response to dietary Ziziphus mauritiana in Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 512, 734301. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Tilapia Culture; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Bansemer, M.S.; Forder, R.E.A.; Howarth, G.S.; Suitor, G.M.; Bowyer, J.; Stone, D.A.J. The effect of dietary soybean meal and soy protein concentrate on the intestinal mucus layer and development of subacute enteritis in Yellowtail Kingfish (Seriola lalandi) at suboptimal water temperature. Aquac. Nutr. 2015, 21, 300–310. [Google Scholar] [CrossRef]
- Jacquin, L.; Gandar, A.; Aguirre-Smith, M.; Perrault, A.; Le Hénaff, M.; De Jong, L.; Paris-Palacios, S.; Laffaille, P.; Jean, S. High temperature aggravates the effects of pesticides in goldfish. Ecotoxicol. Environ. Saf. 2019, 172, 255–264. [Google Scholar] [CrossRef]
- Murosaki, S.; Yamamoto, Y.; Ito, K.; Inokuchi, T.; Kusaka, H.; Ikeda, H.; Yoshikai, Y. Heat-killed Lactobacillus plantarum L-137 suppresses naturally fed antigen-specific IgE production by stimulation of IL-12 production in mice. J. Allergy Clin. Immunol. 1998, 102, 57–64. [Google Scholar] [CrossRef]
- Gewaily, M.S.; Shukry, M.; Abdel-Kader, M.F.; Alkafafy, M.; Farrag, F.A.; Moustafa, E.M.; Doan, H.V.; Abd-Elghany, M.F.; Abdelhamid, A.F.; Eltanahy, A.; et al. Dietary Lactobacillus plantarum relieves Nile tilapia (Oreochromis niloticus) juvenile from oxidative stress, immunosuppression, and inflammation induced by deltamethrin and Aeromonas hydrophila. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Eweedah, N.M.; Moustafa, E.M.; Shahin, M.G. Synbiotic efects of Aspergillus oryzae and β-glucan on growth and oxidative and immune responses of Nile tilapia, Oreochromis niloticus. Probiotics Antimicrob. Proteins 2020, 12, 172–183. [Google Scholar] [CrossRef]
- AOAC. Method 2007-04; Association of Official Analytical Chemists: Washington, DC, USA, 2007. [Google Scholar]
- Cengiz, E.I.; Bayar, A.S.; Kızmaz, V.; Başhan, M.; Satar, A. Acute Toxicity of Deltamethrin on the Fatty Acid Composition of Phospholipid Classes in Liver and Gill Tissues of Nile tilapia. Int. J. Environ. Res. 2017, 11, 377–385. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: New York, NY, USA, 2008. [Google Scholar]
- Hassan, A.; El Nahas, A.; Mahmoud, S.; Barakat, M.; Ammar, A. Thermal stress of ambient temperature modulate expression of stress and immune-related genes and DNA fragmentation in Nile tilapia (Oreochromis niloticus (Linnaeus, 1758)). Appl. Ecol. Environ. Res. 2017, 15, 1343–1354. [Google Scholar] [CrossRef]
- Standen, B.T.; Peggs, D.L.; Rawling, M.D.; Foey, A.; Davies, S.J.; Santos, G.A.; Merrifield, D.L. Dietary administration of a commercial mixed-species probiotic improves growth performance and modulates the intestinal immunity of tilapia, Oreochromis niloticus. Fish Shellfish. Immunol. 2016, 49, 427–435. [Google Scholar] [CrossRef]
- Goes, E.S.D.R.; Goes, M.D.; de Castro, P.L.; de Lara, J.A.F.; Vital, A.C.P.; Ribeiro, R.P. Imbalance of the redox system and quality of tilapia fillets subjected to pre-slaughter stress. PLoS ONE 2019, 14, e0210742. [Google Scholar] [CrossRef] [PubMed]
- Afifi, M.; Saddick, S.; Abu Zinada, O.A. Toxicity of silver nanoparticles on the brain of Oreochromis niloticus and Tilapia zillii. Saudi J. Biol. Sci. 2016, 23, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Qiang, J.; He, J.; Yang, H.; Xu, P.; Habte-Tsion, H.M.; Ma, X.Y.; Zhu, Z.X. The changes in cortisol and expression of immune genes of GIFT tilapia Oreochromis niloticus (L.) at different rearing densities under Streptococcus iniae infection. Aquac. Int. 2016, 24, 1365–1378. [Google Scholar] [CrossRef]
- Abo-Al-Ela, H.G.; El-Nahas, A.F.; Mahmoud, S.; Ibrahim, E.M. Vitamin C Modulates the Immunotoxic Effect of 17α-Methyltestosterone in Nile Tilapia. Biochemistry 2017, 56, 2042–2050. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Stara, A.; Zuskova, E.; Vesely, L.; Kouba, A.; Velisek, J. Single and combined effects of thiacloprid concentration, exposure duration, and water temperature on marbled crayfish Procambarus virginalis. Chemosphere 2020, 273, 128463. [Google Scholar] [CrossRef]
- Dsikowitzky, L.; Nguyen, T.M.I.; Konzer, L.; Zhao, H.; Wang, D.R.; Yang, F.; Schwarzbauer, J. Occurrence and origin of triazine herbicides in a tropical coastal area in China: A potential ecosystem threat. Estuarine Coast. Shelf Sci. 2020, 235, 106612. [Google Scholar] [CrossRef]
- Biggar, J.W.; Riggs, R.L. Apparent solubility of organochlorine insecticides in water at various temperatures. Hilgardia 1974, 42, 383–391. [Google Scholar] [CrossRef]
- Lopes, T.O.M.; Passos, L.S.; Vieira, L.V.; Pinto, E.; Dorr, F.; Scherer, R.; Salustriano, N.D.A.; Carneiro, M.T.W.D.; Postay, L.F.; Gomes, L.C. Metals, arsenic, pesticides, and microcystins in tilapia (Oreochromis niloticus) from aquaculture parks in Brazil. Environ. Sci. Pollut. Res. 2020, 27, 20187–20200. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Abdel-Tawwab, M.; Abdel-Latif, H.M. Lycopene reduces the impacts of aquatic environmental pollutants and physical stressors in fish. Rev. Aquac. 2020, 12, 2511–2526. [Google Scholar] [CrossRef]
- Magouz, F.I.; Dawood, M.A.O.; Salem, M.F.I.; El-Ghandour, M.; van Doan, H.; Mohamed, A.A.I. The role of a digestive enhancer in improving the growth performance, digestive enzymes activity, and health condition of Nile tilapia (Oreochromis niloticus) reared under suboptimal temperature. Aquaculture 2020, 526, 735388. [Google Scholar] [CrossRef]
- Azimirad, M.; Meshkini, S.; Ahmadifard, N.; Hoseinifar, S.H. The effects of feeding with synbiotic (Pediococcus acidilactici and fructooligosaccharide) enriched adult Artemia on skin mucus immune responses, stress resistance, intestinal microbiota and performance of angelfish (Pterophyllum scalare). Fish Shellfish. Immunol. 2016, 54, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Song, S.K.; Beck, B.R.; Kim, D.; Park, J.; Kim, J.; Kim, H.D.; Ringø, E. Prebiotics as immunostimulants in aquaculture: A review. Fish Shellfish. Immunol. 2014, 40, 40–48. [Google Scholar] [CrossRef]
- Rjeibi, I.; Ben Saad, A.; Hfaiedh, N. Oxidative damage and hepatotoxicity associated with deltamethrin in rats: The protective effects of Amaranthus spinosus seed extract. Biomed. Pharmacother. 2016, 84, 853–860. [Google Scholar] [CrossRef]
- Guzman-Villanueva, L.T.; Tovar-Ramírez, D.; Gisbert, E.; Cordero, H.; Guardiola, F.A.; Cuesta, A.; Meseguer, J.; Ascencio-Valle, F.; Esteban, M.A. Dietary administration of β-1,3/1,6-glucan and probiotic strain Shewanella putrefaciens, single or combined, on gilthead seabream growth, immune responses and gene expression. Fish Shellfish. Immunol. 2014, 39, 34–41. [Google Scholar] [CrossRef]
- Aliko, V.; Qirjo, M.; Sula, E.; Morina, V.; Faggio, C. Antioxidant defense system, immune response and erythron profile modulation in gold fish, Carassius auratus, after acute manganese treatment. Fish Shellfish. Immunol. 2018, 76, 101–109. [Google Scholar] [CrossRef]
- Yao, J.; Wang, J.-Y.; Liu, L.; Li, Y.-X.; Xun, A.-Y.; Zeng, W.-S.; Jia, C.-H.; Wei, X.-X.; Feng, J.-L.; Zhao, L.; et al. Anti-oxidant Effects of Resveratrol on Mice with DSS-induced Ulcerative Colitis. Arch. Med. Res. 2010, 41, 288–294. [Google Scholar] [CrossRef]
- Sala, A.; Recio, M.; Schinella, G.R.; Máñez, S.; Giner, R.M.; Cerdá-Nicolás, M.; Ríos, J.-L. Assessment of the anti-inflammatory activity and free radical scavenger activity of tiliroside. Eur. J. Pharmacol. 2003, 461, 53–61. [Google Scholar] [CrossRef]
- Petit, J.; Wiegertjes, G.F. Long-lived effects of administering β-glucans: Indications for trained immunity in fish. Dev. Comp. Immunol. 2016, 64, 93–102. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Sadegh, T.H.; Dawood, M.A.; Dadar, M.; Sheikhzadeh, N. The effects of dietary Pediococcus pentosaceus on growth performance, hemato-immunological parameters and digestive enzyme activities of common carp (Cyprinus carpio). Aquaculture 2020, 516, 734656. [Google Scholar] [CrossRef]
- Ming, J.; Xie, J.; Xu, P.; Liu, W.; Ge, X.; Liu, B.; He, Y.; Cheng, Y.; Zhou, Q.; Pan, L. Molecular cloning and expression of two HSP70 genes in the Wuchang bream (Megalobrama amblycephala Yih). Fish Shellfish. Immunol. 2010, 28, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Craig, E.A. The Heat-Shock Proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Forsyth, R.B.; Candido, E.P.M.; Babich, S.L.; Iwama, G.K. Stress Protein Expression in Coho Salmon with Bacterial Kidney Disease. J. Aquat. Anim. Health 1997, 9, 18–25. [Google Scholar] [CrossRef]
- Voll, R.E.; Herrmann, M.; Roth, E.A.; Stach, C.; Kalden, J.R.; Girkontaite, I. Immunosuppressive effects of apoptotic cells. Nature 1997, 390, 350–351. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Tapingkae, W.; Seel-Audom, M.; Jaturasitha, S.; Dawood, M.A.; Wongmaneeprateep, S.; Thu, T.T.N.; Esteban, M.Á. Boosted growth performance, mucosal and serum immunity, and disease resistance Nile tilapia (Oreochromis niloticus) fingerlings using corncob-derived xylooligosaccharide and Lactobacillus plantarum CR1T5. Probiotics Antimicrob. Proteins 2020, 12, 400–411. [Google Scholar] [CrossRef]
- Yang, P.; Yang, W.; He, M.; Li, X.; Leng, X.-J. Dietary synbiotics improved the growth, feed utilization and intestinal structure of largemouth bass (Micropterus salmoides) juvenile. Aquac. Nutr. 2020, 26, 590–600. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 1–20. [Google Scholar] [CrossRef]
- Sherif, A.H.; Mahfouz, M.E. Immune status of Oreochromis niloticus experimentally infected with Aeromonas hydrophila following feeding with 1, 3 β-glucan and levamisole immunostimulants. Aquaculture 2019, 509, 40–46. [Google Scholar] [CrossRef]
- Cavalcante, R.B.; Telli, G.S.; Tachibana, L.; Dias, D.D.C.; Oshiro, E.; Natori, M.M.; da Silva, W.F.; Ranzani-Paiva, M.J. Probiotics, Prebiotics and Synbiotics for Nile tilapia: Growth performance and protection against Aeromonas hydrophila infection. Aquac. Rep. 2020, 17, 100343. [Google Scholar] [CrossRef]
- Dawood, M.A.O. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2021, 13, 642–663. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | (%) | Composition | (%) |
|---|---|---|---|
| Fish meal | 8 | Dry matter | 90.66 |
| Soybean meal | 42 | Crude protein | 30.05 |
| Wheat bran | 10 | Ether extract | 6.22 |
| Yellow corn | 20 | Crude fibers | 4.95 |
| Gluten | 6 | Total ash | 3.95 |
| Fish oil | 3 | Gross energy (KJ/g) * | 18.98 |
| Dicalcium phosphate | 1 | ||
| Vitamin and mineral mixture | 2 | ||
| Vitamin C | 0.08 | ||
| Starch | 7.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gewaily, M.S.; Abdo, S.E.; Moustafa, E.M.; AbdEl-kader, M.F.; Abd El-Razek, I.M.; El-Sharnouby, M.; Alkafafy, M.; Raza, S.H.A.; El Basuini, M.F.; Van Doan, H.; et al. Dietary Synbiotics Can Help Relieve the Impacts of Deltamethrin Toxicity of Nile Tilapia Reared at Low Temperatures. Animals 2021, 11, 1790. https://doi.org/10.3390/ani11061790
Gewaily MS, Abdo SE, Moustafa EM, AbdEl-kader MF, Abd El-Razek IM, El-Sharnouby M, Alkafafy M, Raza SHA, El Basuini MF, Van Doan H, et al. Dietary Synbiotics Can Help Relieve the Impacts of Deltamethrin Toxicity of Nile Tilapia Reared at Low Temperatures. Animals. 2021; 11(6):1790. https://doi.org/10.3390/ani11061790
Chicago/Turabian StyleGewaily, Mahmoud S., Safaa E. Abdo, Eman M. Moustafa, Marwa F. AbdEl-kader, Ibrahim M. Abd El-Razek, Mohamed El-Sharnouby, Mohamed Alkafafy, Sayed Haidar Abbas Raza, Mohammed F. El Basuini, Hien Van Doan, and et al. 2021. "Dietary Synbiotics Can Help Relieve the Impacts of Deltamethrin Toxicity of Nile Tilapia Reared at Low Temperatures" Animals 11, no. 6: 1790. https://doi.org/10.3390/ani11061790
APA StyleGewaily, M. S., Abdo, S. E., Moustafa, E. M., AbdEl-kader, M. F., Abd El-Razek, I. M., El-Sharnouby, M., Alkafafy, M., Raza, S. H. A., El Basuini, M. F., Van Doan, H., & Dawood, M. A. O. (2021). Dietary Synbiotics Can Help Relieve the Impacts of Deltamethrin Toxicity of Nile Tilapia Reared at Low Temperatures. Animals, 11(6), 1790. https://doi.org/10.3390/ani11061790