
Impacts of Supplementing Broiler Diets with Biological Curcumin, Zinc Nanoparticles and Bacillus licheniformis on Growth, Carcass Traits, Blood Indices, Meat Quality and Cecal Microbial Load



 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
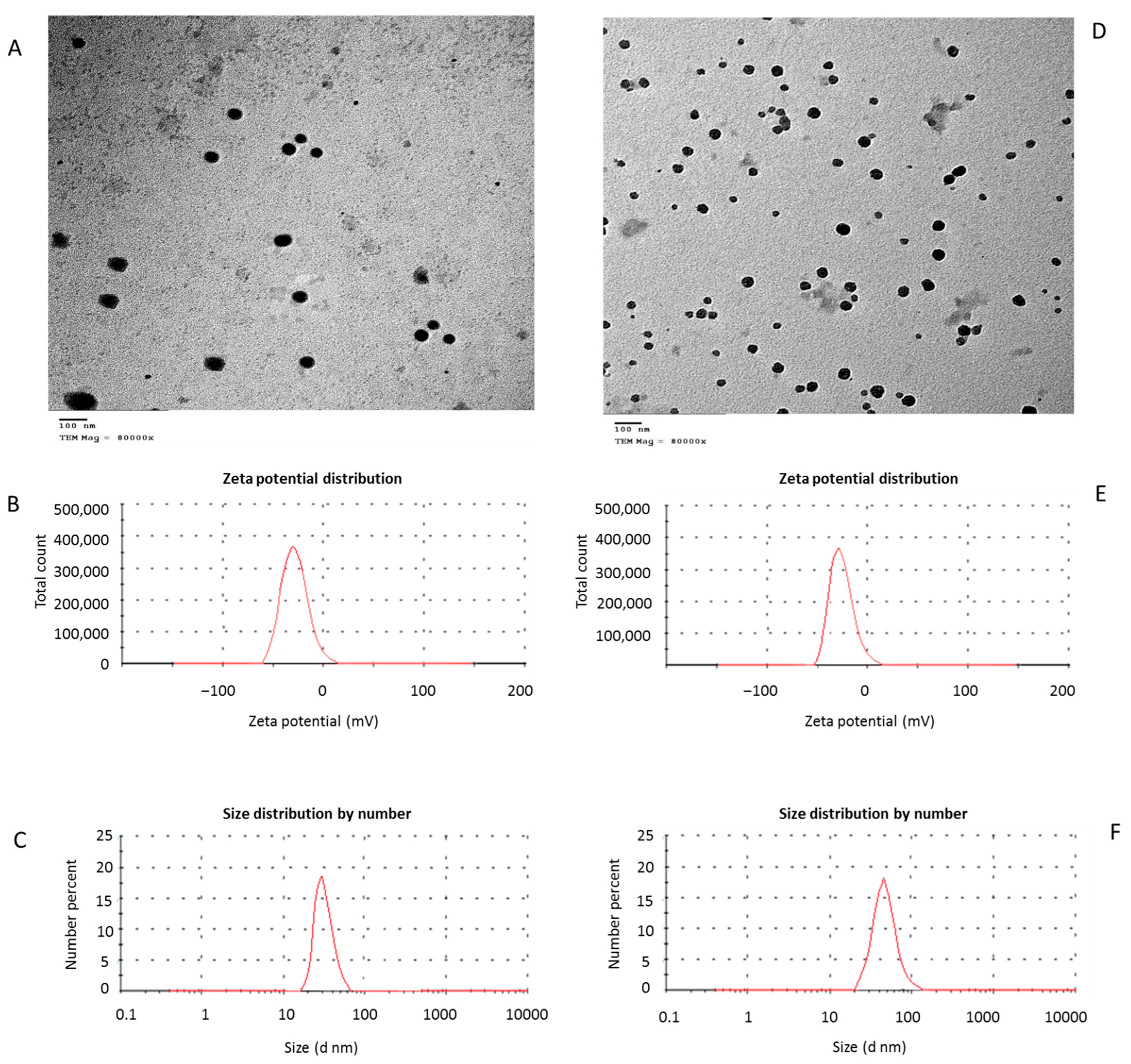
2.1. Nanoparticles Biosynthesis and Antimicrobial Properties
2.1.1. Bacterial Isolates, Biosynthesis and Characterization of ZnNPs and CurNPs
2.1.2. Antimicrobial Activity of CurNPs and ZnNPs
2.2. Birds, Experimental Design and Diets
2.3. Traits Measured
2.3.1. Performance, Carcass, and Blood Biochemical Parameters
2.3.2. Breast Meat Quality and Sensory Evaluation
2.3.3. Microbial Count in Diet and Cecal Samples
2.4. Statistical Analysis
3. Results
3.1. Antimicrobial Activity of CurNPs and ZnNPs
3.2. Growth Performance
3.3. Carcass Traits
3.4. Blood Biochemical Indices
3.5. Meat Quality
3.6. Microbial Count in Diet and Cecal Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, R.U.; Naz, S.; Javdani, M.; Nikousefat, Z.; Selvaggi, M.; Tufarelli, V.; Laudadio, V. The use of turmeric (Curcuma longa) in poultry feed. Worlds Poult. Sci. J. 2012, 68, 97–103. [Google Scholar] [CrossRef]
- European Commission. Ban on Antibiotics as Growth Promoters in Animal Feed Enters into Effect; European Commision: Brussels, Belgium, 2005. [Google Scholar]
- Alagawany, M.; Elnesr, S.; Farag, M. Use of liquorice (Glycyrrhiza glabra) in poultry nutrition: Global impacts on performance, carcass and meat quality. Worlds Poult. Sci. J. 2019, 75, 293–304. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Tiwari, R.; Yatoo, M.; Bhatt, P. Use of licorice (Glycyrrhiza glabra) herb as a feed additive in poultry: Current knowledge and prospects. Animals 2019, 9, 536. [Google Scholar] [CrossRef] [Green Version]
- Khafaga, A.F.; Abd El-Hack, M.E.; Taha, A.E.; Elnesr, S.S.; Alagawany, M. The potential modulatory role of herbal additives against Cd toxicity in human, animal, and poultry: A review. Environ. Sci. Pollut. Res. 2019, 26, 4588–4604. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.; Ashour, E.; Elaraby, G.M.; Osman, A.; Arif, M. Influences of dietary supplementation of peanut skin powder (Arachis Hypogaea) on growth performance, carcass traits, blood chemistry, antioxidant activity and meat quality of broilers. Anim. Prod. Sci. 2018, 58, 965–972. [Google Scholar] [CrossRef]
- Ashour, E.A.; Abd El-Hack, M.E.; Swelum, A.A.; Osman, A.O.; Taha, A.E.; Alhimaidi, A.R.; Ismail, I.E. Does the dietary graded levels of herbal mixture powder impact growth, carcass traits, blood indices and meat quality of the broilers? Ital. J. Anim. Sci. 2020, 19, 1226–1235. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alkhatib, F.M.; Alzahrani, S.O.; Shafi, M.E.; Abdel-Hamid, S.E.; Taha, T.F.; Aboelenin, S.M.; Soliman, M.M.; Ahmed, N.H. Impact of mycogenic zinc nanoparticles on performance, behavior, immune response, and microbial load in Oreochromis niloticus. Saudi J. Biol. Sci. 2021, in press. [Google Scholar] [CrossRef]
- Akl, B.; Nader, M.M.; El-Saadony, M.T. Biosynthesis of silver nanoparticles by Serratia marcescens ssp sakuensis and its antibacterial application against some pathogenic bacteria. J. Agric. Chem. Biotechnol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Najjar, A.A.; Alzahrani, S.O.; Alkhatib, F.M.; Shafi, M.E.; Selem, E.; Desoky, E.-S.M.; Fouda, S.E.; El-Tahan, A.M.; et al. The use of biological selenium nanoparticles to suppress Triticum aestivum L. crown and root rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 2021, in press. [Google Scholar] [CrossRef]
- Li, X.; Xu, H.; Chen, Z.; Chen, G. Biosynthesis of nanoparticles by microorganisms and their applications. J. Nanomater. 2011, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wijnhoven, S.W.; Peijnenburg, W.J.; Herberts, C.A.; Hagens, W.I.; Oomen, A.G.; Heugens, E.H.; Roszek, B.; Bisschops, J.; Gosens, I.; Van De Meent, D. Nano-silver–a review of available data and knowledge gaps in human and environmental risk assessment. Nanotoxicology 2009, 3, 109–138. [Google Scholar] [CrossRef]
- Liao, C.-D.; Hung, W.-L.; Jan, K.-C.; Yeh, A.-I.; Ho, C.-T.; Hwang, L.S. Nano/sub-microsized lignan glycosides from sesame meal exhibit higher transport and absorption efficiency in Caco-2 cell monolayer. Food Chem. 2010, 119, 896–902. [Google Scholar] [CrossRef]
- Jahanian, R.; Moghaddam, H.N.; Rezaei, A. Improved broiler chick performance by dietary supplementation of organic zinc sources. Asian-australas. J. Anim. Sci. 2008, 21, 1348–1354. [Google Scholar]
- NRC. National Research Council. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994.
- Feng, J.; Ma, W.; Niu, H.; Wu, X.; Wang, Y. Effects of zinc glycine chelate on growth, hematological, and immunological characteristics in broilers. Biol. Trace Elem. Res. 2010, 133, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Sagar, P.D.; Mandal, A.; Akbar, N.; Dinani, O. Effect of different levels and sources of zinc on growth performance and immunity of broiler chicken during summer. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 459–471. [Google Scholar] [CrossRef]
- Reda, F.M.; El-Saadony, M.T.; El-Rayes, T.K.; Attia, A.I.; El-Sayed, S.A.; Ahmed, S.Y.; Madkour, M.; Alagawany, M. Use of biological nano zinc as a feed additive in quail nutrition: Biosynthesis, antimicrobial activity and its effect on growth, feed utilisation, blood metabolites and intestinal microbiota. Ital. J. Anim. Sci. 2021, 20, 324–335. [Google Scholar] [CrossRef]
- Akhavan-Salamat, H.; Ghasemi, H. Effect of different sources and contents of zinc on growth performance, carcass characteristics, humoral immunity and antioxidant status of broiler chickens exposed to high environmental temperatures. Livest. Sci. 2019, 223, 76–83. [Google Scholar] [CrossRef]
- Ahmadi, F.; Ebrahimnezhad, Y.; Sis, N.M.; Ghiasi, J. The effects of zinc oxide nanoparticles on performance, digestive organs and serum lipid concentrations in broiler chickens during starter period. Int. J. Biosci. 2013, 3, 23–29. [Google Scholar] [CrossRef]
- Zhao, C.-Y.; Tan, S.-X.; Xiao, X.-Y.; Qiu, X.-S.; Pan, J.-Q.; Tang, Z.-X. Effects of dietary zinc oxide nanoparticles on growth performance and antioxidative status in broilers. Biol. Trace Elem. Res. 2014, 160, 361–367. [Google Scholar] [CrossRef]
- Toghyani, M.; Toghyani, M.; Gheisari, A.; Ghalamkari, G.; Eghbalsaied, S. Evaluation of cinnamon and garlic as antibiotic growth promoter substitutions on performance, immune responses, serum biochemical and haematological parameters in broiler chicks. Livest. Sci. 2011, 138, 167–173. [Google Scholar] [CrossRef]
- Rukkumani, R.; Aruna, K.; Varma, P.S.; Rajasekaran, K.N.; Menon, V.P. Comparative effects of curcumin and an analog of curcumin on alcohol and PUFA induced oxidative stress. J. Pharm. Pharm. Sci. 2004, 7, 274–283. [Google Scholar]
- Zhai, S.; Ruan, D.; Zhu, Y.; Li, M.; Ye, H.; Wang, W.; Yang, L. Protective effect of curcumin on ochratoxin A–induced liver oxidative injury in duck is mediated by modulating lipid metabolism and the intestinal microbiota. Poult. Sci. 2020, 99, 1124–1134. [Google Scholar] [CrossRef]
- Reda, F.M.; El-Saadony, M.T.; Elnesr, S.S.; Alagawany, M.; Tufarelli, V. Effect of dietary supplementation of biological curcumin nanoparticles on growth and carcass traits, antioxidant status, immunity and caecal microbiota of Japanese quails. Animals 2020, 10, 754. [Google Scholar] [CrossRef]
- Shaikh, J.; Ankola, D.; Beniwal, V.; Singh, D.; Kumar, M.R. Nanoparticle encapsulation improves oral bioavailability of curcumin by at least 9-fold when compared to curcumin administered with piperine as absorption enhancer. Eur. J. Pharm. Sci. 2009, 37, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Rajput, N.; Muhammah, N.; Yan, R.; Zhong, X.; Wang, T. Effect of dietary supplementation of curcumin on growth performance, intestinal morphology and nutrients utilization of broiler chicks. J. Poult. Sci. 2012, 0120065. [Google Scholar] [CrossRef] [Green Version]
- Sayrafi, R.; Hosseini, S.; Ahmadi, M. The protective effects of nanocurcumin on liver toxicity induced by salinomycin in broiler chickens. Rev. Méd. Vét. 2017, 168, 136–142. [Google Scholar]
- Partovi, R.; Seifi, S.; Pabast, M.; Babaei, A. Effects of dietary supplementation with nanocurcumin on quality and safety of meat from broiler chicken infected with Eimeria species. J. Food Saf. 2019, 39, e12703. [Google Scholar] [CrossRef]
- Ahuja, K.; Mamtani, K. Poultry Probiotic Ingredients Market Size by Product. In (Lactobacilli, Bifidobacterium, Streptococcus, Bacillus), by Application (Broilers, Layers, Turkeys, Breeders, Chicks & Poults), Regional Outlook, Application Potential, Price Trends, Competitive Market Share & Forecast, 2019–2025; Global Market Insights: Selbyville, DE, USA, 2019. [Google Scholar]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (direct-fed microbials) in poultry nutrition and their effects on nutrient utilization, growth and laying performance, and gut health: A systematic review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Attia, Y.; Zeweil, H.; Alsaffar, A.; El-Shafy, A. Effect of non-antibiotic feed additives as an alternative to flavomycin on productivity, meat quality and blood parameters in broilers. Arch. Geflügelk 2011, 75, 40–48. [Google Scholar]
- Santoso, U.; Tanaka, K.; Ohtani, S.; Sakaida, M. Effect of fermented product from Bacillus subtilis on feed conversion efficiency, lipid accumulation and ammonia production in broiler chicks. Asian Australas. J. Anim. Sci. 2001, 14, 333–337. [Google Scholar] [CrossRef]
- Abou-Kassem, D.; Elsadek, M.; Abdel-Moneim, A.; Mahgoub, S.; Elaraby, G.; Taha, A.; Elshafie, M.; Alkhawtani, D.; Abd El-Hack, M.; Ashour, E. Growth, carcass characteristics, meat quality, and microbial aspects of growing quail fed diets enriched with two different types of probiotics (Bacillus toyonensis and Bifidobacterium bifidum). Poult. Sci. 2021, 100, 84–93. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Abd El-Hack, M.E.; Taha, A.E.; Fouda, M.M.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; Elshaer, N. Ecofriendly synthesis and insecticidal application of copper nanoparticles against the storage pest Tribolium castaneum. Nanomaterials 2020, 10, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Saadony, M.T.; El-Wafai, N.A.; El-Fattah, H.I.A.; Mahgoub, S.A. Biosynthesis, optimization and characterization of silver nanoparticles using a soil isolate of Bacillus pseudomycoides MT32 and their antifungal activity against some pathogenic fungi. Adv. Anim. Vet. Sci. 2019, 7, 238–249. [Google Scholar] [CrossRef] [Green Version]
- Kannan, M.N.; Sethi, S.; Badoni, A.; Chamoli, V.; Bahuguna, N.C. Isolation and characterization of bacterial isolates from agriculture field soil of Roorkee region. J. Pharm. Phytochem. 2018, 5, 108–110. [Google Scholar]
- El-Saadony, M.T.; Desoky, E.-S.M.; Saad, A.M.; Eid, R.S.; Selem, E.; Elrys, A.S. Biological silicon nanoparticles improve Phaseolus vulgaris L. yield and minimize its contaminant contents on a heavy metals-contaminated saline soil. J. Environ. Sci. 2021, 106, 1–14. [Google Scholar] [CrossRef]
- Elbeshehy, E.K.; Elazzazy, A.M.; Aggelis, G. Silver nanoparticles synthesis mediated by new isolates of Bacillus spp., nanoparticle characterization and their activity against Bean Yellow Mosaic Virus and human pathogens. Front. Microbiol. 2015, 6, 453. [Google Scholar] [CrossRef] [Green Version]
- Saad, A.M.; El-Saadony, M.T.; El-Tahan, A.M.; Sayed, S.; Moustafa, M.A.; Taha, A.E.; Taha, T.F.; Ramadan, M.M. Polyphenolic extracts from pomegranate and watermelon wastes as substrate to fabricate Sustainable Silver nanoparticles with larvicidal effect against Spodoptera littoralis. Saudi J. Biol. Sci. 2021, in press. [Google Scholar] [CrossRef]
- Mishra, M.; Paliwal, J.S.; Singh, S.K.; Selvarajan, E.; Subathradevi, C.; Mohanasrinivasan, V. Studies on the inhibitory activity of biologically synthesized and characterized zinc oxide nanoparticles using lactobacillus sporogens against Staphylococcus aureus. J. Pure. Appl. Microbiol. 2013, 7, 1–6. [Google Scholar]
- Gulluce, M.; Sahin, F.; Sokmen, M.; Ozer, H.; Daferera, D.; Sokmen, A.; Polissiou, M.; Adiguzel, A.; Ozkan, H. Antimicrobial and antioxidant properties of the essential oils and methanol extract from Mentha longifolia L. ssp. longifolia. Food Chem. 2007, 103, 1449–1456. [Google Scholar] [CrossRef]
- Rahmoun, N.M.; Ziane, H.; Boucherit-Otmani, Z. Antibacterial and antifungal screening of four medicinal plants. J. Coast. Life Med. 2014, 2, 975–979. [Google Scholar]
- Eucast. Determination of minimum inhibitory concentrations (MICs) of antimicrobial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, 1–7. [Google Scholar]
- CLSI. Performance standards for Antimicrobial Susceptibility Testing, 18th Informational Supplement. M100-S18; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Ali, S.S.; Morsy, R.; El-Zawawy, N.A.; Fareed, M.F.; Bedaiwy, M.Y. Synthesized zinc peroxide nanoparticles (ZnO2-NPs): A novel antimicrobial, anti-elastase, anti-keratinase, and anti-inflammatory approach toward polymicrobial burn wounds. Int. J. Nanomed. 2017, 12, 6059. [Google Scholar] [CrossRef] [Green Version]
- Usman, J.G.; Sodipo, O.; Sandabe, U. In vitro antimicrobial activity of Cucumis metuliferus E. Mey. Ex. Naudin fruit extracts against Salmonella gallinarum. Int. J. Phytomed. 2014, 6, 268. [Google Scholar]
- Fidan, E.D.; Nazlıgül, A.; Türkyılmaz, M.K.; Aypak, S.Ü.; Kilimci, F.S.; Karaarslan, S.; Kaya, M. Effect of photoperiod length and light intensity on some welfare criteria, carcass, and meat quality characteristics in broilers. Rev. Bras. Zootec. 2017, 46, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Trela, J.; Kierończyk, B.; Hautekiet, V.; Józefiak, D. Combination of bacillus licheniformis and salinomycin: Effect on the growth performance and git microbial populations of broiler chickens. Animals 2020, 10, 889. [Google Scholar] [CrossRef] [PubMed]
- IR-BroilerHandbook2018-EN.pdf. 2018. A.I.R.B.M.H. Available online: http://www.aviagen.com (accessed on 27 May 2018).
- Abdelnour, S.A.; Swelum, A.A.; Salama, A.; Al-Ghadi, M.Q.; Qattan, S.Y.; Abd El-Hack, M.E.; Khafaga, A.F.; Alhimaidi, A.R.; Almutairi, B.O.; Ammari, A.A.; et al. The beneficial impacts of dietary phycocyanin supplementation on growing rabbits under high ambient temperature. Ital. J. Anim. Sci. 2020, 19, 1046–1056. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A.; Gharib, A.; Chobert, J.-M.; Haertlé, T. Preliminary assessment of potential toxicity of methylated soybean protein and methylated β-lactoglobulin in male Wistar rats. Food Chem. Toxicol. 2013, 59, 618–625. [Google Scholar] [CrossRef]
- Salvaggio, A.; Periti, M.; Miano, L.; Tavanelli, M.; Marzorati, D. Body mass index and liver enzyme activity in serum. Clin. Chem. 1991, 37, 720–723. [Google Scholar] [CrossRef]
- Owen, J.; Iggo, B.; Scandrett, F.; Stewart, C. The determination of creatinine in plasma or serum, and in urine; A critical examination. Biochem. J. 1954, 58, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, M.; Osman, A.; El-Hadary, A.; Romeih, E.; Sitohy, M.; Li, L. Hepatoprotective action of papain-hydrolyzed buffalo milk protein on carbon tetrachloride oxidative stressed albino rats. J. Dairy Sci. 2020, 103, 1884–1893. [Google Scholar] [CrossRef]
- Wattanachant, S.; Benjakul, S.; Ledward, D. Composition, color, and texture of Thai indigenous and broiler chicken muscles. Poult. Sci. 2004, 83, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Wattanachant, S.; Benjakul, S.; Ledward, D. Microstructure and thermal characteristics of Thai indigenous and broiler chicken muscles. Poult. Sci. 2005, 84, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lopez, J.; Zhi, N.; Aleson-Carbonell, L.; Pérez-Alvarez, J.A.; Kuri, V. Antioxidant and antibacterial activities of natural extracts: Application in beef meatballs. Meat Sci. 2005, 69, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Botta, J.; Lauder, J.; Jewer, M. Effect of methodology on total volatile basic nitrogen (TVB-N) determination as an index of quality of fresh Atlantic cod (Gadus morhua). J. Food Sci. 1984, 49, 734–736. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2005. [Google Scholar]
- Zhao, C.; Liu, Y.; Lai, S.; Cao, H.; Guan, Y.; San Cheang, W.; Liu, B.; Zhao, K.; Miao, S.; Riviere, C. Effects of domestic cooking process on the chemical and biological properties of dietary phytochemicals. Trends Food Sci. Technol. 2019, 85, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Sow, T.; Grongnet, J.-F. Sensory characteristics and consumer preference for chicken meat in Guinea. Poult. Sci. 2010, 89, 2281–2292. [Google Scholar] [CrossRef]
- Zhuang, H.; Savage, E.M. Effect of postmortem deboning time on sensory descriptive flavor and texture profiles of cooked boneless skinless broiler thighs. LWT Food Sci. Technol. 2011, 44, 2087–2090. [Google Scholar] [CrossRef]
- Harrigan, W.F.; McCance, M.E. Laboratory Methods in Food and Dairy Microbiology; Academic Press Inc.: London, UK, 1976. [Google Scholar]
- Alagawany, M.; El-Saadony, M.; Elnesr, S.; Farahat, M.; Attia, G.; Madkour, M.; Reda, F. Use of lemongrass essential oil as a feed additive in quail’s nutrition: Its effect on growth, carcass, blood biochemistry, antioxidant and immunological indices, digestive enzymes and intestinal microbiota. Poult. Sci. 2021, 100, 101172. [Google Scholar] [CrossRef]
- Alagawany, M.; Madkour, M.; El-Saadony, M.T.; Reda, F.M. Paenibacillus polymyxa (LM31) as a new feed additive: Antioxidant and antimicrobial activity and its effects on growth, blood biochemistry, and intestinal bacterial populations of growing Japanese quail. Anim. Feed Sci. Technol. 2021, 276, 114920. [Google Scholar] [CrossRef]
- Ashour, E.A.; Abd El-Hack, M.E.; Shafi, M.E.; Alghamdi, W.Y.; Taha, A.E.; Swelum, A.A.; Tufarelli, V.; Mulla, Z.S.; El-Ghareeb, W.R.; El-Saadony, M.T. Impacts of green coffee powder supplementation on growth performance, carcass characteristics, blood indices, meat quality and gut microbial load in broilers. Agriculture 2020, 10, 457. [Google Scholar] [CrossRef]
- Sheiha, A.M.; Abdelnour, S.A.; Abd El-Hack, M.E.; Mohamed, E.; Khafaga, A.F.; Metwally, K.A.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; El-Saadony, M.T. Effects of dietary biological or chemical-synthesized nano-selenium supplementation on growing rabbits exposed to thermal stress. Animals 2020, 10, 430. [Google Scholar] [CrossRef] [Green Version]
- Szabo, R.A.; Todd, E.C.; Jean, A. Method to isolate Escherichia coli O157: H7 from food. J. Food Prot. 1986, 49, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E.A.; Hilderbrand, R.L. Method for identifying Salmonella and Shigella directly from the primary isolation plate by coagglutination of protein A-containing staphylococci sensitized with specific antibody. J. Clin. Microbiol. 1976, 3, 339–343. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Franco, C.; Vázquez, B.; Fente, C.; Barros-Velázquez, J.; Cepeda, A. Evaluation of Chromocult® enterococci agar for the isolation and selective enumeration of Enterococcus spp. in broilers. Lett. Appl. Microbiol. 2005, 41, 153–156. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Sitohy, M.Z.; Ramadan, M.F.; Saad, A.M. Green nanotechnology for preserving and enriching yogurt with biologically available iron (II). Innov. Food Sci. Emerg. Technol. 2021, 102645. [Google Scholar] [CrossRef]
- Bhawana, B.R.; Buttar, H.S.; Jain, V.; Jain, N. Curcumin nanoparticles: Preparation, characterization, and antimicrobial study. J. Agric. Food Chem. 2011, 59, 2056–2061. [Google Scholar] [CrossRef]
- Narayanan, P.; Wilson, W.S.; Abraham, A.T.; Sevanan, M. Synthesis, characterization, and antimicrobial activity of zinc oxide nanoparticles against human pathogens. BioNanoScience 2012, 2, 329–335. [Google Scholar] [CrossRef]
- Azam, A.; Ahmed, A.S.; Oves, M.; Khan, M.S.; Habib, S.S.; Memic, A. Antimicrobial activity of metal oxide nanoparticles against Gram-positive and Gram-negative bacteria: A comparative study. Int. J. Nanomed. 2012, 7, 6003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.M.; Hasan, H.; Mohamad, D. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nano Micro Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef] [Green Version]
- Hassani Sangani, M.; Nakhaei Moghaddam, M.; Forghanifard, M.M. Inhibitory effect of zinc oxide nanoparticles on pseudomonas aeruginosa biofilm formation. Nanomed. J. 2015, 2, 121–128. [Google Scholar]
- Arciniegas-Grijalba, P.; Patiño-Portela, M.; Mosquera-Sánchez, L.; Guerrero-Vargas, J.; Rodríguez-Páez, J. ZnO nanoparticles (ZnO-NPs) and their antifungal activity against coffee fungus Erythricium salmonicolor. Appl. Nanosci. 2017, 7, 225–241. [Google Scholar] [CrossRef] [Green Version]
- Alnashi, B.; Fattah, A.A. Antimicrobial activity of raw and nano turmeric powder extracts. Middle East. J. Appl. Sci. 2016, 6, 787–796. [Google Scholar]
- Rhayat, L.; Jacquier, V.; Brinch, K.; Nielsen, P.; Nelson, A.; Geraert, P.-A.; Devillard, E. Bacillus subtilis s train specificity affects performance improvement in broilers. Poult. Sci. 2017, 96, 2274–2280. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, K.; Sharma, K.; Katoch, S.; Sharma, V.; Mane, B. Probiotics in broiler poultry feeds: A review. J. Anim. Nutr. Physiol. 2015, 1, 4–16. [Google Scholar]
- Hafez, A.; Hegazi, S.; Bakr, A.; Shishtawy, H. Effect of zinc oxide nanoparticles on growth performance and absorptive capacity of the intestinal villi in broiler chickens. Life Sci. J. 2017, 14, 67–72. [Google Scholar]
- Abedini, M.; Shariatmadari, F.; Torshizi, M.K.; Ahmadi, H. Effects of a dietary supplementation with zinc oxide nanoparticles, compared to zinc oxide and zinc methionine, on performance, egg quality, and zinc status of laying hens. Live Sci. 2017, 203, 30–36. [Google Scholar] [CrossRef]
- Mahmoud, U.T.; Abdel-Mohsein, H.S.; Mahmoud, M.A.; Amen, O.A.; Hassan, R.I.; Abd-El-Malek, A.M.; Rageb, S.M.; Waly, H.S.; Othman, A.A.; Osman, M.A. Effect of zinc oxide nanoparticles on broilers’ performance and health status. Trop. Anim. Health Prod. 2020, 52, 2043–2054. [Google Scholar] [CrossRef]
- Fathi, M.; Haydari, M.; Tanha, T. Effects of zinc oxide nanoparticles on antioxidant status, serum enzymes activities, biochemical parameters and performance in broiler chickens. J. Livest. Sci. Technol. 2016, 4, 7–13. [Google Scholar]
- Sandoval, M.; Henry, P.; Luo, X.; Littell, R.; Miles, R.; Ammerman, C. Performance and tissue zinc and metallothionein accumulation in chicks fed a high dietary level of zinc. Poult. Sci. 1998, 77, 1354–1363. [Google Scholar] [CrossRef]
- Roberson, K.D.; Edwards, H.M., Jr. Effects of 1,25-dihydroxycholecalciferol and phytase on zinc utilization in broiler chicks. Poult. Sci. 1994, 73, 1312–1326. [Google Scholar] [CrossRef] [PubMed]
- Fazilati, M. Investigation toxicity properties of zinc oxide nanoparticles on liver enzymes in male rat. Eur. J. Exp. Biol. 2013, 3, 97–103. [Google Scholar]
- Hani, U.; Shivakumar, H. Solubility enhancement and delivery systems of curcumin a herbal medicine: A review. Curr. Drug Deliv. 2014, 11, 792–804. [Google Scholar] [CrossRef]
- Platel, K.; Srinivasan, K. Influence of dietary spices and their active principles on pancreatic digestive enzymes in albino rats. Food Nahr. 2000, 44, 42–46. [Google Scholar] [CrossRef]
- Chattopadhyay, I.; Biswas, K.; Bandyopadhyay, U.; Banerjee, R.K. Turmeric and curcumin: Biological actions and medicinal applications. Curr. Sci. 2004, 44–53. [Google Scholar]
- Sahoo, A.; Swain, R.; Mishra, S.K. Effect of inorganic, organic and nano zinc supplemented diets on bioavailability and immunity status of broilers. Int. J. Adv. Res. 2014, 2, 828–837. [Google Scholar]
- Lina, T.; Jianyang, J.; Fenghua, Z.; Huiying, R.; Wenli, L. Effect of nano-zinc oxide on the production and dressing performance of broiler. Chin. Agric. Sci. Bull. 2009, 2, 318. [Google Scholar]
- Abd El-Moneim, A.E.-M.E.; El-Wardany, I.; Abu-Taleb, A.M.; Wakwak, M.M.; Ebeid, T.A.; Saleh, A.A. Assessment of in ovo administration of Bifidobacterium bifidum and Bifidobacterium longum on performance, ileal histomorphometry, blood hematological, and biochemical parameters of broilers. Probiotics Antimicro Proteins 2019, 1–12. [Google Scholar] [CrossRef]
- Premavalli, K.; Sangilimadan, K.; Omprakash, A. Effect of supplementation of multi-species probiotic on production performance of Japanese quail. Inter. J. Chem. Stud. 2018, 6, 2164–2166. [Google Scholar]
- Abdel-Moneim, A.-M.E.; Selim, D.A.; Basuony, H.A.; Sabic, E.M.; Saleh, A.A.; Ebeid, T.A. Effect of dietary supplementation of Bacillus subtilis spores on growth performance, oxidative status, and digestive enzyme activities in Japanese quail birds. Rop. Anim. Health Prod. 2020, 52, 671–680. [Google Scholar] [CrossRef]
- Jin, L.; Ho, Y.; Abdullah, N.; Ali, M.; Jalaludin, S. Effects of adherent Lactobacillus cultures on growth, weight of organs and intestinal microflora and volatile fatty acids in broilers. Anim. Feed Sci. Technol. 1998, 70, 197–209. [Google Scholar] [CrossRef]
- Badawi, M.; Ali, M.; Behairy, A. Effects of zinc sources supplementation on performance of broiler chickens. J. Am. Sci. 2017, 13, 35–40. [Google Scholar]
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and anti-inflammatory effects of zinc. Zinc-dependent NF-κB signaling. Inflammopharmacology 2017, 25, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, D.; Ali, H.A.; El-Mandrawy, S.A. Effects of different zinc sources on performance, bio distribution of minerals and expression of genes related to metabolism of broiler chickens. Zagazig Vet. J. 2017, 45, 292–304. [Google Scholar] [CrossRef] [Green Version]
- El-Katcha, M.; Soltan, M.A.; El-Badry, M. Effect of dietary replacement of inorganic zinc by organic or nanoparticles sources on growth performance, immune response and intestinal histopathology of broiler chicken. Alex. J. Vet. Sci. 2017, 55. [Google Scholar] [CrossRef] [Green Version]
- Emadi, M.; Kermanshahi, H. Effect of turmeric rhizome powder on the activity of some blood enzymes in broiler chickens. Int. J. Poult. Sci 2007, 6, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Gholiyan, A.; Jafari, M.R. Effect of different levels of nanoparticle curcumin on egg quality, blood parameters, immune response and digestibility in laying hens. Res. Anim. Prod. Sci. Res. 2018, 9, 26–34. [Google Scholar]
- Unnikrishnan, M.; Rao, M. Inhibition of nitrite induced oxidation of hemoglobin by curcuminoids. Die Pharm. 1995, 50, 490–492. [Google Scholar]
- De Beer, M.; McMurtry, J.; Brocht, D.; Coon, C. An examination of the role of feeding regimens in regulating metabolism during the broiler breeder grower period. 2. Plasma hormones and metabolites. Poult. Sci. 2008, 87, 264–275. [Google Scholar] [CrossRef] [PubMed]
- AL-Kassie, G.A.; Mohseen, A.M.; Abd-AL-Jaleel, R.A. Modification of productive performance and physiological aspects of broilers on the addition of a mixture of cumin and turmeric to the diet. Res. Opin. Anim. Vet. Sci. 2011, 27, 847–854. [Google Scholar]
- Das, K.C.; Das, C.K. Curcumin (diferuloylmethane), a singlet oxygen (1O2) quencher. Biochem. Biophys. Res. Commun. 2002, 295, 62–66. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Tuzcu, Z.; Tuzcu, M.; Sahin, N. Curcumin ameloriates heat stress via inhibition of oxidative stress and modulation of Nrf2/HO-1 pathway in quail. Food Chem. Toxicol. 2012, 50, 4035–4041. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Huang, H.; Zhou, L.; Li, W.; Zhou, H.; Hou, G.; Liu, J.; Hu, L. Effects of dietary supplementation with turmeric rhizome extract on growth performance, carcass characteristics, antioxidant capability, and meat quality of Wenchang broiler chickens. Ital. J. Anim. Sci. 2015, 14, 3870. [Google Scholar] [CrossRef]
- Nawab, A.; Li, G.; Liu, W.; Lan, R.; Wu, J.; Zhao, Y.; Kang, K.; Kieser, B.; Sun, C.; Tang, S. Effect of dietary curcumin on the antioxidant status of laying hens under high-temperature condition. J. Therm. Biol. 2019, 86, 102449. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Aggarwal, B.B. “Spicing up” of the immune system by curcumin. J. Clin. Immunol. 2007, 27, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Al-Daraji, H.J.; Amen, M.H. Effect of dietary zinc on certain blood traits of broiler breeder chickens. Int. J. Poult. Sci. 2011, 10, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Alkhalf, A.; Alhaj, M.; Al-Homidan, I. Influence of probiotic supplementation on blood parameters and growth performance in broiler chickens. Saudi J. Biol. Sci. 2010, 17, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Bech-Larsen, T.; Scholderer, J. Functional foods in Europe: Consumer research, market experiences and regulatory aspects. Trends Food Sci. Technol. 2007, 18, 231–234. [Google Scholar] [CrossRef]
- Banon, S.; Díaz, P.; Rodríguez, M.; Garrido, M.D.; Price, A. Ascorbate, green tea and grape seed extracts increase the shelf life of low sulphite beef patties. Meat Sci. 2007, 77, 626–633. [Google Scholar] [CrossRef]
- Ahmed, I.; El-Rayes, T.; Ahmed, A.I. Assessment of dietary supplementation of turmeric (curcuma longa) as a phytobiotic on broiler performance and bacterial count. Egypt. J. Nutr. Feeds 2018, 21, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Wang, H.; Zhang, P.; Gao, C.; Tao, J.; Ge, Z.; Zhu, D.; Bi, Y. Modulation of gut microbiota contributes to curcumin-mediated attenuation of hepatic steatosis in rats. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Mahajan, S.; Sharma, R. Evaluation of antimicrobial activity of Curcuma longa rhizome extract against Staphylococcus aureus. Biotechnol. Rep. 2015, 6, 51–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niamsa, N.; Sittiwet, C. Antimicrobial activity of Curcuma longa aqueous extract. J. Pharmacol. Toxicol. 2009, 4, 173–177. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Items | Starter (1–3 Weeks) | Finisher (3–5 Weeks) |
|---|---|---|
| Ingredients % | ||
| Yellow corn | 55.89 | 57 |
| Soybean meal 44% | 31.5 | 29.5 |
| Gluten meal 60% | 6.5 | 4.83 |
| Dicalcium phosphate | 1.7 | 1.7 |
| Limestone | 1.24 | 1.15 |
| Vit-min Premix * | 0.3 | 0.3 |
| NaCl | 0.3 | 0.3 |
| DL-Methionine | 0.13 | 0.0 |
| L-Lysine HCl | 0.24 | 0.18 |
| Choline 50% | 0.2 | 0.2 |
| Soybean oil | 2.0 | 4.84 |
| Total | 100 | 100 |
| Calculated analysis: | ||
| Dry matter % | 91.72 | 90.43 |
| Crude protein % | 23.00 | 20.94 |
| Metabolizable energy (kcal/kg diet) | 2996.30 | 3150.70 |
| Calcium % | 1.00 | 0.96 |
| Phosphorous (Available) % | 0.44 | 0.44 |
| Lysine % | 1.3 | 1.17 |
| Methionine + Cysteine % | 0.90 | 0.70 |
| Crude fiber % | 3.52 | 3.38 |
| Microorganisms | ZnNPs (µg/mL) | Means | CurNPs (µg/mL) | Means | MIC | MBC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | 100 | 150 | 200 | 250 | 300 | 100 | 150 | 200 | 250 | 300 | ZnNPs | CurNPs | ZnNPs | CurNPs | ||
| B. cereus | 17.26 b | 18.22 b | 20.91 b | 21.65 b | 22.84 b | 20.18 AB | 18.32 b | 19.62 b | 21.41 b | 22.22 b | 23.43 b | 21.00 AB | 60.12 d | 55.23 d | 110.00 e | 100.31 e |
| L. monocytogenes | 16.55 c | 17.53 c | 19.52 c | 20.54 c | 23.42 c | 19.5 B | 17.43 c | 18.93 c | 20.95 c | 23.43 c | 24.22 c | 20.99 B | 75.21 cd | 69.24 cd | 130.01 d | 120.21 d |
| S. pyogenes | 18.36 a | 19.45 a | 21.83 a | 22.92 a | 24.53 a | 21.42 A | 19.55 a | 20.34 a | 22.53 a | 23.65 a | 25.64 a | 22.34 A | 50.00 e | 45.33 e | 90.23 f | 85.43 f |
| E. coli | 15.28 d | 16.96 d | 18.81 d | 19.57 d | 20.67 d | 18.26 BC | 16.28 d | 18.38 d | 19.24 d | 20.56 d | 21.48 d | 19.19 C | 80.13 c | 71.11 c | 140.14 c | 130.24 c |
| S. typhi | 14.38 e | 15.28 e | 17.24 e | 18.48 e | 19.88 e | 17.05 C | 15.29 e | 16.97 e | 18.56 e | 19.41 e | 20.69 e | 18.18 CD | 85.36 b | 80.10 b | 150.62 b | 140.78 b |
| p. aeruginosa | 12.79 f | 14.82 f | 16.46 f | 17.55 f | 18.52 f | 16.03 D | 13.41 f | 15.59 f | 17.44 f | 18.22 f | 19.52 f | 16.84 D | 95.61 a | 90.34 a | 170.14 a | 160.14 a |
| Means | 15.77 D | 17.04 C | 19.13 B | 20.12 AB | 21.64 A | 16.71 D | 18.31 C | 20.02 B | 21.25 AB | 22.50 A | 74.41 A | 68.56 B | 131.86 A | 122.85 B | ||
| SEM | 0.23 | 0.56 | 0.37 | 0.67 | 0.50 | 0.36 | 0.44 | 0.40 | 0.70 | 0.49 | 0.56 | 0.60 | 0.88 | 0.99 | ||
| p-value | ||||||||||||||||
| Bacteria (B) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||||||
| Concentration (C) | <0.001 | <0.001 | ||||||||||||||
| B × C | <0.001 | <0.001 | ||||||||||||||
| Fungi | Means | Means | MIC | MFC | ||||||||||||
| A. alternate | 20.91 c | 21.51 c | 22.92 c | 24.21 c | 25.25 c | 22.96 B | 21.42 c | 22.24 c | 23.54 c | 25.02 c | 26.42 c | 23.73 B | 70.12 c | 65.33 c | 120.23 d | 110.24 d |
| A. flavus | 21.81 b | 22.32 b | 24.23 b | 26.52 b | 28.22 b | 24.62 AB | 22.32 b | 23.05 b | 24.82 b | 27.33 b | 29.42 b | 25.39 AB | 65.13 cd | 60.27 cd | 110.24 e | 105.34 e |
| F. oxysporum | 19.62 d | 20.55 d | 21.94 d | 23.24 d | 24.23 d | 21.92 C | 20.13 d | 21.22 d | 22.55 d | 24.05 d | 25.43 d | 22.68 BC | 75.52 bc | 70.31 bc | 140.29 c | 130.24 c |
| A. niger | 22.43 a | 24.84 a | 25.79 a | 27.65 a | 29.14 a | 25.97 A | 22.95 a | 25.52 a | 26.38 a | 28.46 a | 30.24 a | 26.71 A | 55.15 d | 50.51 d | 100.23 f | 90.34 f |
| p. solitum | 18.52 e | 19.57 e | 20.88 e | 22.67 e | 23.42 e | 21.01 C | 19.06 e | 20.21 e | 21.42 e | 23.47 e | 24.68 e | 21.77 C | 80.33 b | 75.36 b | 150.14 b | 140.22 b |
| p. crustosum | 16.43 f | 18.22 f | 19.54 f | 21.48 f | 22.61 f | 19.66 D | 17.92 f | 18.93 f | 20.11 f | 22.22 f | 23.87 f | 20.61 D | 95.56 a | 90.25 a | 170.34 a | 155.31 a |
| Means | 19.95 D | 21.17 C | 22.55 B | 24.30 AB | 25.48 A | 20.63 D | 21.86 C | 23.14 B | 25.09 AB | 26.68 A | 73.64 A | 68.67 B | 131.91 A | 121.95 B | ||
| SEM | 0.25 | 0.60 | 0.33 | 0.74 | 0.43 | 0.32 | 0.41 | 0.43 | 0.77 | 0.40 | 0.51 | 0.62 | 0.90 | 0.83 | ||
| p-value | ||||||||||||||||
| Fungi (F) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||||||||
| Concentration (C) | <0.001 | <0.001 | ||||||||||||||
| F × C | <0.001 | <0.001 | ||||||||||||||
| Items | Treatments | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | 1 | 2 | 3 | 4 | 5 | 6 | |||
| Starter period (1–3 weeks) | |||||||||
| LBW (g) | 667.41 b | 642.36 d | 645.36 d | 652.45 c | 668.18 b | 665.95 b | 683.73 a | 4.03 | 0.038 |
| BWG (g/day) | 40.53 a | 36.40 b | 33.15 c | 35.04 bc | 38.02 b | 37.25 b | 40.12 a | 0.59 | <0.001 |
| FI (g/day) | 70.74 a | 55.42 b | 53.53 b | 54.13 b | 56.87 b | 57.52 b | 58.08 b | 1.33 | <0.001 |
| FCR | 1.74 a | 1.52 c | 1.61 b | 1.55 c | 1.50 c | 1.54 c | 1.45 d | 0.02 | 0.004 |
| Finisher period (3–5 weeks) | |||||||||
| LBW (g) | 1843.30 e | 2161.91 b | 2062.36 de | 2084.15 d | 2143.43 c | 2177.60 a | 2190.54 a | 26.26 | <0.001 |
| BWG (g) | 81.05 c | 117.61 a | 112.38 ab | 109.27 b | 112.30 ab | 118.72 a | 114.54 ab | 2.74 | <0.001 |
| FI (g) | 116.48 d | 148.56 a | 150.57 a | 131.66 c | 136.11 c | 150.47 a | 141.66 b | 3.13 | 0.007 |
| FCR | 1.44 a | 1.26 c | 1.34 b | 1.20 d | 1.21 d | 1.27 c | 1.24 c | 0.02 | 0.034 |
| Whole cycle (1–5 weeks) | |||||||||
| LBW (g) | 1843.30 e | 2161.91 b | 2062.36 de | 2084.15 d | 2143.43 c | 2177.60 a | 2190.54 a | 26.26 | <0.001 |
| BWG (g/day) | 56.08 c | 65.22 a | 61.69 b | 63.05 b | 64.87 a | 65.52 a | 66.28 a | 0.78 | <0.001 |
| FI (g/day) | 87.69 | 89.84 | 84.65 | 84.02 | 86.88 | 90.63 | 88.31 | 0.75 | 0.133 |
| FCR | 1.56 a | 1.39 b | 1.40 b | 1.35 c | 1.35 c | 1.39 b | 1.33 c | 0.02 | 0.001 |
| Mortality rate (%) | 2.30 | 2.72 | 2.72 | 0.00 | 1.82 | 2.72 | 0.00 | 0.82 | 0.542 |
| Carcass Traits (as a % of Pre-Slaughter Weight) | Items | ||||||
|---|---|---|---|---|---|---|---|
| Bursa | Spleen | Abdominal Fat | Dressing | Giblets | Carcass | Pre-Slaughter Weight | |
| 0.10 | 0.12 b | 1.11 b | 77.29 d | 3.28 e | 74.00 d | 2010.00 bc | Control |
| 0.12 | 0.08 c | 0.81 d | 78.69 b | 4.20 a | 74.49 c | 2047.50 b | T1 |
| 0.08 | 0.09 c | 1.32 a | 77.51 d | 3.88 b | 73.63 e | 1960.00 c | T2 |
| 0.14 | 0.12 b | 0.38 e | 78.85 b | 3.76 c | 75.09 b | 1995.00 bc | T3 |
| 0.13 | 0.16 a | 0.94 c | 79.19 ab | 3.60 d | 75.58 a | 2240.00 a | T4 |
| 0.15 | 0.12 b | 0.84 d | 78.08 c | 3.76 c | 74.32 c | 2020.00 bc | T5 |
| 0.17 | 0.12 b | 0.79 d | 79.55 a | 3.71 c | 75.85 a | 2075.00 b | T6 |
| 0.01 | 0.01 | 0.07 | 0.23 | 0.11 | 0.71 | 21.45 | SEM |
| 0.135 | 0.017 | 0.002 | 0.006 | <0.001 | 0.010 | 0.001 | p-value |
| Serum Parameters | Control | T1 | T2 | T3 | T4 | T5 | T6 | SEM | p Value |
|---|---|---|---|---|---|---|---|---|---|
| Hematological | |||||||||
| Hemoglobin (g/dL) | 6.90 bc | 7.00 b | 7.20 ab | 7.00 b | 7.00 b | 7.20 ab | 7.30 a | 0.051 | <0.0001 |
| MCH (P/g) | 22.90 c | 23.00 bc | 23.80 b | 23.00 bc | 22.90 c | 24.00 ab | 25.10 a | 0.279 | <0.0001 |
| MCV (Fl) | 63.00 d | 63.50 c | 64.00 bc | 63.80 c | 64.50 b | 65.00 ab | 66.50 a | 0.405 | <0.0001 |
| MCHC (g/d) | 33.22 cd | 33.90 c | 34.50 b | 34.00 bc | 33.50 c | 35.10 ab | 35.50 a | 0.285 | <0.0001 |
| PCV (%) | 22.25 c | 22.50 bc | 25.20 ab | 21.50 cd | 23.00 b | 24.50 b | 25.90 a | 0.579 | <0.0001 |
| RBC (×106/U L) | 3.30 b | 3.40 b | 3.60 ab | 2.50 c | 3.40 b | 3.50 ab | 3.80 a | 0.144 | <0.0001 |
| WBC (×103 j/L) | 4.00 a | 4.00 a | 3.25 c | 2.10 d | 3.50 b | 3.12 bc | 4.10 a | 0.251 | <0.0001 |
| Platelets (103 j/L) | 210 cd | 212 c | 215 bc | 199 e | 219 b | 205 d | 225 a | 3.027 | <0.0001 |
| Lymphocytes (%) | 72.00 bc | 67.10 d | 73.10 b | 72.10 bc | 72.12 bc | 70.31 c | 75.12 a | 0.877 | <0.0001 |
| Neutrophils (%) | 21.00 d | 21.10 d | 27.12 b | 25.60 c | 29.12 ab | 23.10 cd | 30.12 a | 1.286 | <0.0001 |
| Basophils (%) | 0.00 | 0.00 | 0.00 | 1.10 b | 1.20 b | 0.90 bc | 1.50 a | 0.228 | <0.0001 |
| Monocytes (%) | 0.30 d | 0.40 d | 1.90 bc | 0.90 c | 2.30 b | 2.50 ab | 2.80 a | 0.363 | <0.0001 |
| Eosinophils (%) | 4.30 b | 4.50 ab | 3.80 c | 4.30 b | 4.50 ab | 4.10 c | 5.13 a | 0.135 | <0.0001 |
| Biochemical | |||||||||
| Glucose (mmol/L) | 6.12 bc | 6.11 bc | 6.22 bc | 6.30 ab | 6.10 b | 6.50 a | 6.01 bc | 0.069 | <0.0001 |
| Total protein (g/dL) | 4.50 d | 4.90 d | 5.80 b | 6.50 a | 5.12 c | 5.10 c | 6.30 ab | 0.266 | <0.0001 |
| Albumin (g/dL) | 3.20 cd | 3.50 c | 4.10 b | 4.10 b | 4.00 b | 4.20 ab | 4.50 a | 0.153 | <0.0001 |
| Globulin (g/dL) | 2.10 d | 2.50 c | 2.60 c | 2.40 cd | 2.50 c | 3.05 b | 3.60 a | 0.171 | <0.0001 |
| Chloride (mmol/L) | 105 a | 99 d | 95 d | 100 c | 103 b | 100 c | 90.00 e | 1.755 | <0.0001 |
| Calcium (mmol/L) | 2.20 c | 2.40 c | 3.50 a | 3.20 ab | 3.05 b | 3.05 b | 3.50 a | 0.177 | <0.0001 |
| AST (U/I) | 23.12 c | 25.42 bc | 25.12 bc | 35.12 a | 27.05 b | 34.22 ab | 20.10 c | 1.948 | <0.0001 |
| ALT (U/I) | 20.14 c | 16.10 d | 19.01 c | 25.22 ab | 23.23 b | 26.14 a | 18.10 cd | 1.309 | <0.0001 |
| Urea (mmol/L) | 5.12 a | 4.20 b | 3.90 c | 5.01 a | 4.25 b | 3.99 c | 4.10 bc | 0.161 | <0.0001 |
| Triglycerides (mmol/L) | 1.20 a | 1.10 ab | 0.90 c | 1.05 b | 1.01 b | 0.98 bc | 0.70 c | 0.056 | <0.0001 |
| Cholesterol (mmol/L) | 4.10 ab | 4.20 a | 4.05 b | 4.20 a | 4.04 c | 4.15 bc | 4.12 c | 0.030 | <0.0001 |
| LDL (mmol/L) | 3.40 ab | 3.50 a | 1.70 c | 3.40 ab | 1.70 c | 2.40 b | 1.60 c | 0.310 | <0.0001 |
| VLDL (mmol/L) | 0.44 ab | 0.45 a | 0.30 b | 0.28 c | 0.17 | 0.32 ab | 0.25 c | 0.035 | <0.0001 |
| Oxidative enzymes | |||||||||
| GSH | 1.10 d | 1.30 c | 1.30 c | 1.20 cd | 1.50 b | 1.70 ab | 1.80 a | 0.091 | <0.0001 |
| GSR | 1.30 d | 1.50 cd | 1.60 c | 1.35 cd | 1.80 b | 1.90 ab | 2.00 a | 0.095 | <0.0001 |
| GST | 1.40 cd | 1.60 c | 1.60 c | 1.31 cd | 1.80 b | 2.00 b | 2.30 a | 0.121 | <0.0001 |
| SOD | 1.50 e | 1.70 d | 1.80 cd | 1.90 c | 2.40 b | 2.50 ab | 2.70 a | 0.160 | <0.0001 |
| MDA | 17.90 a | 17.10 ab | 16.80 b | 16.20 bc | 16.20 bc | 14.50 c | 13.10 d | 0.575 | <0.0001 |
| Duodenal Enzyme activity | |||||||||
| Amylase | 2830 g | 2935 f | 3040 e | 3145 d | 3750 c | 4022 b | 4155 a | 192.42 | <0.0001 |
| Protease | 130 fg | 140 f | 170 e | 180 d | 190 c | 225 b | 234 a | 13.72 | <0.0001 |
| Lipase | 95.00f | 100 e | 111 de | 120 d | 130 c | 140 b | 145 a | 6.75 | <0.0001 |
| Immunoglobulin level | |||||||||
| IgA | 7.50 cd | 7.80 c | 7.90 bc | 8.00 bc | 8.50 b | 9.70 ab | 10.20 a | 0.362 | <0.0001 |
| IgM | 2.50 cd | 2.90 c | 3.00 cd | 3.10 cd | 3.50 c | 3.90 b | 4.50 a | 0.237 | <0.0001 |
| IgG | 13.00 cd | 14.30 c | 15.90 bc | 16.00 bc | 16.00 bc | 17.00 b | 18.90 a | 0.658 | <0.0001 |
| Quality Parameters | Control | T1 | T2 | T3 | T4 | T5 | T6 | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Chemical | |||||||||
| Moisture | 65.9 cd | 64.30 d | 68.00 b | 68.20 b | 67.10 c | 70.00 ab | 71.00 a | 0.803 | <0.0001 |
| Protein | 19.45 d | 20.12 c | 21.40 c | 22.00 b | 21.20 c | 21.22 c | 23.00 a | 0.408 | <0.0001 |
| Lipid | 14.1 ab | 15.00 a | 10.20 c | 9.30 cd | 11.50 b | 9.00 d | 6.00 e | 1.087 | <0.0001 |
| Ash | 0.89 b | 1.10 a | 0.90 b | 1.00 ab | 1.00 ab | 0.80 c | 0.30 d | 0.092 | <0.0001 |
| pH | 5.5 de | 6.00 c | 5.80 d | 6.10 c | 6.30 b | 6.20 bc | 6.80 a | 0.143 | <0.0001 |
| TBVN | 6.4 a | 5.90 b | 5.90 b | 5.60 bc | 5.50 bc | 5.10 c | 4.80 cd | 0.187 | <0.0001 |
| TBA | 0.60 a | 0.60 a | 0.5 ab | 0.4 b | 0.3 b | 0.2 c | 0.2 c | 0.061 | <0.0001 |
| Color | |||||||||
| L* | 60.10 ab | 58.20 c | 60.20 ab | 60.00 ab | 59.60 b | 59.0 bc | 61.20 a | 0.334 | <0.0001 |
| a* | 6.00 bc | 6.50 ab | 6.00 bc | 6.00 bc | 6.70 a | 6.40 b | 5.80 c | 0.116 | <0.0001 |
| b* | 15.00 bc | 15.00 bc | 15.90 b | 14.10 cd | 14.80 c | 14.20 cd | 16.10 a | 0.268 | <0.0001 |
| Sensorial | |||||||||
| Juiciness | 4.20 bc | 4.05 c | 4.35 b | 4.30 b | 4.4 b | 4.56 ab | 4.80 a | 0.085 | <0.0001 |
| Tenderness | 4.90 b | 4.75 bc | 4.70 c | 4.95 ab | 4.70 c | 4.96 ab | 5.20 a | 0.063 | <0.0001 |
| Taste | 4.35 c | 4.20 cd | 4.44 b | 4.34 c | 4.20 cd | 4.56 ab | 4.80 a | 0.075 | <0.0001 |
| Aroma | 4.50 ab | 4.35 c | 4.50 ab | 4.55 ab | 4.56 ab | 4.46 b | 4.70 a | 0.037 | <0.0001 |
| Samples/Microbial Count | TBC | p Value | TYMC | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Feeding Period (day) | 0 | 7 | 14 | 21 | 0 | 7 | 14 | 21 | ||
| Control | 5.80 a,D | 6.01 a,C | 6.33 a,B | 6.70 a,A | <0.001 | 3.81 a,D | 4.00 a,C | 4.30 a,B | 4.82 a,A | <0.001 |
| T1 | 5.54 ab,D | 5.72 b,C | 6.12 ab,B | 6.51 ab,A | <0.001 | 3.52 ab,D | 3.72 ab,C | 4.03 ab,B | 4.55 b,A | <0.001 |
| T2 | 5.20 bD | 5.41 bcC | 5.93 bB | 6.22 b,A | <0.001 | 3.14 b,D | 3.51 b,C | 3.82 b,B | 4.12 bc,A | <0.001 |
| T3 | 4.86 c,D | 5.14 c,C | 5.50 bc,B | 5.90 bc,A | <0.001 | 2.82 bc,D | 3.14 c,C | 3.44 c,B | 3.84 c,A | <0.001 |
| T4 | 4.60 d,D | 4.95 cd,C | 5.32 c,B | 5.74 c,A | <0.001 | 2.58 c,D | 2.90 cd,C | 3.15 cd,B | 3.69 cd,A | <0.001 |
| T5 | 4.22 d,D | 4.72 d,C | 5.11 cd,B | 5.38 cd,A | <0.001 | 2.30 cd,D | 2.71 d,C | 2.82 d,B | 3.33 d,A | <0.001 |
| T6 | 3.93 e,A | 4.11 e,C | 4.85 d,B | 5.09 d,A | <0.001 | 2.10 d,D | 2.42 de,C | 2.68 de,B | 3.15 de,A | <0.001 |
| SEM | 0.25 | 0.27 | 0.61 | 0.44 | 0.46 | 0.64 | 0.09 | 0.14 | ||
| p value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||
| Samples/Microbial Count | E. coli | pValue | Coliform | pValue | ||||||
| Feeding Period (day) | 0 | 7 | 14 | 21 | 0 | 7 | 14 | 21 | ||
| Control | 2.22 a,C | 2.56 a,B | 2.92 a,AB | 3.12 a,A | <0.001 | 2.91 a,D | 3.33 a,C | 3.62 a,B | 4.02 a,A | <0.001 |
| T1 | 2.01 ab,D | 2.32 ab,C | 2.62 ab,B | 2.81 ab,A | <0.001 | 2.72 ab,D | 3.05 ab,C | 3.43 ab,B | 3.81 ab,A | <0.001 |
| T2 | 1.88 b,D | 2.11 b,C | 2.31 b,B | 2.54 b,A | <0.001 | 2.44 b,D | 2.84 b,C | 3.11 b,B | 3.51 b,A | <0.001 |
| T3 | 1.56 bc,D | 1.92 c,C | 2.15 bc,B | 2.37 bc,A | <0.001 | 2.21 bc,D | 2.58 bc,C | 2.94 bc,B | 3.23 c,A | <0.001 |
| T4 | 1.34 c,C | 1.73 c,B | 1.96 c,A | 2.08 c,A | <0.001 | 1.94 c,C | 2.11 c,C | 2.77 c,B | 3.07 cd,A | <0.001 |
| T5 | 1.12 cd,D | 1.54 cd,C | 1.64 cd,B | 1.82 cd,A | <0.001 | 1.66 cd,C | 1.84 d,C | 2.52 cd,B | 2.88 d,A | <0.001 |
| T6 | 0.85 d,C | 1.29 d,B | 1.48 d,A | 1.53 d,A | <0.001 | 1.48 d,D | 1.62 de,C | 2.29 d,B | 2.57 e,A | <0.001 |
| SEM | 0.21 | 0.23 | 0.59 | 0.40 | 0.42 | 0.60 | 0.07 | 0.10 | ||
| p value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||
| Samples | Total Bacteria | Total Yeasts and Molds | E. coli | Salmonella spp. | Enterococcus spp. | Coliform | Lactic Acid Bacteria |
|---|---|---|---|---|---|---|---|
| Control | 9.20 a | 3.92 a | 5.70 a | 1.58 | 5.82 a | 6.70 a | 4.51 d |
| T1 | 8.40 b | 3.59 ab | 5.50 ab | ND | 5.60 b | 6.51 ab | 4.81 cd |
| T2 | 7.83 bc | 3.52 b | 5.30 b | ND | 5.50 b | 6.23 b | 5.20 c |
| T3 | 7.65 c | 3.41 bc | 5.11 c | ND | 5.21 bc | 6.25 c | 5.42 c |
| T4 | 7.40 cd | 3.21 c | 4.82 cd | ND | 5.12 c | 5.84 c | 5.62 b |
| T5 | 7.25 d | 3.00 cd | 4.62 d | ND | 4.90 c | 5.55 cd | 5.81 ab |
| T6 | 6.50 e | 2.80 d | 4.44 d | ND | 4.71 d | 5.26 d | 6.18 a |
| SEM | 0.17 | 0.22 | 0.14 | 0.20 | 0.30 | 0.19 | 0.20 |
| p value | 0.001 | 0.004 | 0.001 | 0.001 | 0.012 | 0.021 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Hack, M.E.; Alaidaroos, B.A.; Farsi, R.M.; Abou-Kassem, D.E.; El-Saadony, M.T.; Saad, A.M.; Shafi, M.E.; Albaqami, N.M.; Taha, A.E.; Ashour, E.A. Impacts of Supplementing Broiler Diets with Biological Curcumin, Zinc Nanoparticles and Bacillus licheniformis on Growth, Carcass Traits, Blood Indices, Meat Quality and Cecal Microbial Load. Animals 2021, 11, 1878. https://doi.org/10.3390/ani11071878
Abd El-Hack ME, Alaidaroos BA, Farsi RM, Abou-Kassem DE, El-Saadony MT, Saad AM, Shafi ME, Albaqami NM, Taha AE, Ashour EA. Impacts of Supplementing Broiler Diets with Biological Curcumin, Zinc Nanoparticles and Bacillus licheniformis on Growth, Carcass Traits, Blood Indices, Meat Quality and Cecal Microbial Load. Animals. 2021; 11(7):1878. https://doi.org/10.3390/ani11071878
Chicago/Turabian StyleAbd El-Hack, Mohamed E., Bothaina A. Alaidaroos, Reem M. Farsi, Diaa E. Abou-Kassem, Mohamed T. El-Saadony, Ahmed M. Saad, Manal E. Shafi, Najah M. Albaqami, Ayman E. Taha, and Elwy A. Ashour. 2021. "Impacts of Supplementing Broiler Diets with Biological Curcumin, Zinc Nanoparticles and Bacillus licheniformis on Growth, Carcass Traits, Blood Indices, Meat Quality and Cecal Microbial Load" Animals 11, no. 7: 1878. https://doi.org/10.3390/ani11071878
APA StyleAbd El-Hack, M. E., Alaidaroos, B. A., Farsi, R. M., Abou-Kassem, D. E., El-Saadony, M. T., Saad, A. M., Shafi, M. E., Albaqami, N. M., Taha, A. E., & Ashour, E. A. (2021). Impacts of Supplementing Broiler Diets with Biological Curcumin, Zinc Nanoparticles and Bacillus licheniformis on Growth, Carcass Traits, Blood Indices, Meat Quality and Cecal Microbial Load. Animals, 11(7), 1878. https://doi.org/10.3390/ani11071878