
Scorpion Venom Peptide Effects on Inhibiting Proliferation and Inducing Apoptosis in Canine Mammary Gland Tumor Cell Lines

 , , ,
, , ,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Antimicrobial Peptides (AMPs)
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Annexin V/Propidium Iodide (PI) Apoptosis Analysis by Flow Cytometry
2.5. RNA Isolation and Real-Time PCR
2.6. Statistical Analysis
3. Results
3.1. Effect of BmKn-2 on Cell Viability of CMGT Cell Lines
3.1.1. Effect of BmKn-2 on Percentage Viability of CHMP-5b and CHMP-13a Cell Lines
3.1.2. Effect of BmKn-2 on Viability and Morphology of CHMp-5b and CHMp-13a Cell Lines
3.2. Effect of BmKn-2 on Apoptosis Induction in CMGT Cell Lines
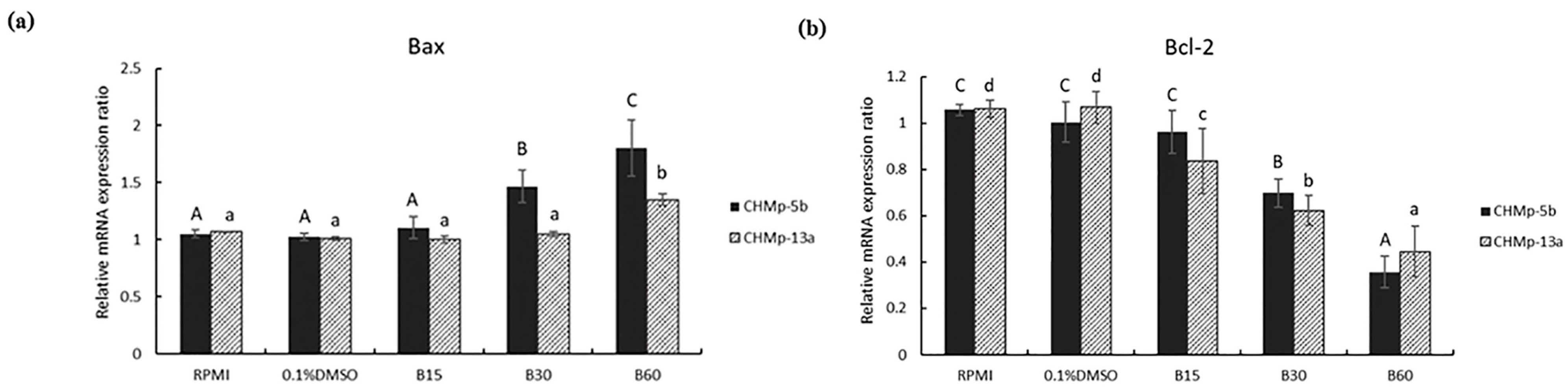
3.3. Effect of BmKn-2 on Apoptosis Induction and Identify Apoptosis Pathways in CMGT Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salas, Y.; Márquez, A.; Diaz, D.; Romero, L. Epidemiological study of mammary tumors in female dogs diagnosed during the period 2002–2012: A growing animal health problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef] [Green Version]
- Thumser-Henner, P.; Nytko, K.J.; Bley, C.R. Mutations of BRCA2 in canine mammary tumors and their targeting potential in clinical therapy. BMC Vet. Res. 2020, 16, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas, C.; Peña, L.; Granados-Soler, J.; Pérez-Alenza, M. Adjuvant therapy for highly malignant canine mammary tumours: Cox-2 inhibitor versus chemotherapy: A case–control prospective study. Vet. Rec. 2016, 30, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, C.G.; Pino, A.M.; Sierralta, W.D. A cyclized peptide derived from α fetoprotein inhibits the proliferation of ER-positive canine mammary cancer cells. Oncol. Rep. 2009, 21, 1397–1404. [Google Scholar] [PubMed]
- Karayannopoulou, M.; Lafioniatis, S. Recent advances on canine mammary cancer chemotherapy: A review of studies from 2000 to date. Breast Cancer Res. 2016, 29, 43. [Google Scholar]
- Ettinger, S.; Feldman, E. Chemotherapy. In Textbook of Veterinary Internal Medicine, 4th ed.; W. B. Saunders Company, Elsevier, Health Sciences Division 11830 Westline Industrial Drive: St. Louis, MO, USA, 1995; pp. 473–484. [Google Scholar]
- Karayannopoulou, M.; Kaldrymidou, E.; Constantinidis, T.; Dessiris, A. Adjuvant post-operative chemotherapy in bitches with mammary cancer. J. Vet. Med. Ser. A 2001, 48, 85–96. [Google Scholar] [CrossRef]
- Yigit, N.; Benli, M. The venom gland of the scorpion species Euscorpius mingrelicus (Scorpiones: Euscorpiidae): Morphological and ultrastructural characterization. J. Venom. Anim. Toxins Incl. Trop. Dis. 2008, 14, 466–480. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-R.; Ji, Y.-H. Scorpion Venom Research Around the World: Chinese Scorpion Mesobuthus martensii Karsch. Scorpion Venoms Toxinology 2015, 383–385. [Google Scholar] [CrossRef]
- Liu, G.; Yang, F.; Li, F.; Li, Z.; Lang, Y.; Shen, B.; Wu, Y.; Li, W.; Harrison, P.L.; Strong, P.N. Therapeutic potential of a scorpion venom-derived antimicrobial peptide and its homologs against antibiotic-resistant gram-positive bacteria. Front. Microbiol. 2018, 9, 1159. [Google Scholar] [CrossRef]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [Green Version]
- Seyfi, R.; Kahaki, F.A.; Ebrahimi, T.; Montazersaheb, S.; Eyvazi, S.; Babaeipour, V.; Tarhriz, V. Antimicrobial peptides (AMPs): Roles, functions and mechanism of action. Int. J. Pept. Res. Ther. 2020, 26, 1451–1463. [Google Scholar] [CrossRef]
- Teerapo, K.; Roytrakul, S.; Sistayanarain, A.; Kunthalert, D. A scorpion venom peptide derivative BmKn‒22 with potent antibiofilm activity against Pseudomonas aeruginosa. PLoS ONE 2019, 14, e0218479. [Google Scholar] [CrossRef] [Green Version]
- Tong-ngam, P.; Roytrakul, S.; Sritanaudomchai, H. BmKn-2 scorpion venom peptide for killing oral cancer cells by apoptosis. Asian Pac. J. Cancer Prev. 2015, 16, 2807–2811. [Google Scholar] [CrossRef] [Green Version]
- Satitmanwiwat, S.; Changsangfa, C.; Khanuengthong, A.; Promthep, K.; Roytrakul, S.; Arpornsuwan, T.; Saikhun, K.; Sritanaudomchai, H. The scorpion venom peptide BmKn2 induces apoptosis in cancerous but not in normal human oral cells. Biomed. Pharmacother. 2016, 84, 1042–1050. [Google Scholar] [CrossRef]
- Arpornsuwan, T.; Sriwai, W.; Jaresitthikunchai, J.; Phaonakrop, N.; Sritanaudomchai, H.; Roytrakul, S. Anticancer activities of antimicrobial BmKn2 peptides against oral and colon cancer cells. Int. J. Pept. Res. Ther. 2014, 20, 501–509. [Google Scholar] [CrossRef]
- Giribaldi, J.; Smith, J.J.; Schroeder, C.I. Recent developments in animal venom peptide nanotherapeutics with improved selectivity for cancer cells. Biotechnol. Adv. 2021, 50, 14. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaliotas, G.I.; Mavridis, K.; Scorilas, A.; Babis, G.C. Quantitative analysis of the mRNA expression levels of BCL2 and BAX genes in human osteoarthritis and normal articular cartilage: An investigation into their differential expression. Mol. Med. Rep. 2015, 12, 4514–4521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Flórez, M.; Fêo, H.B.; Da Silva, G.; Yamatogi, R.S.; Aguiar, A.; Araújo Jr, J.; Rocha, N.S. Cell cycle kinetics, apoptosis rates and gene expressions of MDR-1, TP53, BCL-2 and BAX in transmissible venereal tumour cells and their association with therapy response. Vet. Comp. Oncol. 2017, 15, 793–807. [Google Scholar] [CrossRef]
- Li, H.; Tong, J.; Bao, J.; Tang, D.; Tian, W.; Liu, Y. Hematoporphyrin monomethyl ether combined with He-Ne laser irradiation-induced apoptosis in canine breast cancer cells through the mitochondrial pathway. J. Vet. Sci. 2016, 17, 235. [Google Scholar] [CrossRef]
- Paoloni, M.; Khanna, C. Translation of new cancer treatments from pet dogs to humans. Nat. Rev. Cancer 2008, 8, 147–156. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine mammary tumours as a model to study human breast cancer: Most recent findings. In Vivo 2011, 25, 455–465. [Google Scholar] [PubMed]
- Murai, K.; Nakagawa, T.; Endo, Y.; Kamida, A.; Yoshida, K.; Mochizuki, M.; Nishimura, R.; Sasaki, N. Establishment of a pair of novel cloned tumour cell lines with or without metastatic potential from canine mammary adenocarcinoma. Res. Vet. Sci. 2012, 93, 468–472. [Google Scholar] [CrossRef]
- Foong, J.N.; Selvarajah, G.T.; Rasedee, A.; Rahman, H.S.; How, C.W.; Beh, C.Y.; Teo, G.Y.; Ku, C.L. Zerumbone-loaded nanostructured lipid carrier induces apoptosis of canine mammary adenocarcinoma cells. BioMed Res. Int. 2018, 2018, 8691569. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Li, H.; Song, X.; Wu, Y.; Liu, Y. 5-Azacytidine treatment induces demethylation of DAPK1 and MGMT genes and inhibits growth in canine mammary gland tumor cells. OncoTargets Ther. 2018, 11, 2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Le, W.; Wang, Y.; Li, Z.; Wang, D.; Ren, L.; Lin, L.; Cui, S.; Hu, J.J.; Hu, Y. Targeting negative surface charges of cancer cells by multifunctional nanoprobes. Theranostics 2016, 6, 1887. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wideranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.C.; Soriano, B.J.; Lush, R.M.; Borner, M.M.; Figg, W.D. Apoptosis: Its role in the development of malignancies and its potential as a novel therapeutic target. Ann. Pharmacother. 1997, 31, 76–82. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Savill, J.; Fadok, V. Corpse clearance defines the meaning of cell death. Nature 2000, 407, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.K.; Yazan, L.S.; Ismail, M. Thymoquinone from Nigella sativa was more potent than cisplatin in eliminating of SiHa cells via apoptosis with down-regulation of Bcl-2 protein. Toxicol. Vitr. 2011, 25, 1392–1398. [Google Scholar] [CrossRef]
- Liang, Y.; Arnold, T.; Michlmayr, A.; Rainprecht, D.; Perticevic, B.; Spittler, A.; Oehler, R. Serum-dependent processing of late apoptotic cells for enhanced efferocytosis. Cell Death Dis. 2014, 5, e1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, N.; Verma, R.; Pawar, H.N.; Deka, D.; Agrawal, R.; Singh, A. Cloning, sequence analysis and structure prediction of B cell lymphoma-2 of Canis familiaris. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 370–382. [Google Scholar]
- Mitra, N.; Verma, R.; Deka, D.; Pawar, H.N.; Sood, N.K.; Gupta, K.; Mahajan, S.K.; Mohindroo, J. Differential expression of apoptosis-associated genes in canine mammary tumors. Biologia 2015, 70, 846–852. [Google Scholar] [CrossRef]
- Makin, G.W.; Corfe, B.M.; Griffiths, G.J.; Thistlethwaite, A.; Hickman, J.A.; Dive, C. Damage-induced Bax N-terminal change, translocation to mitochondria and formation of Bax dimers/complexes occur regardless of cell fate. EMBO J. 2001, 20, 6306–6315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, A.; Jockel, J.; Wei, M.C.; Korsmeyer, S.J. Enforced dimerization of BAX results in its translocation, mitochondrial dysfunction and apoptosis. EMBO J. 1998, 17, 3878–3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korsmeyer, S.J.; Shutter, J.R.; Veis, D.J.; Merry, D.E.; Oltvai, Z.N. Bcl-2/Bax: A rheostat that regulates an anti-oxidant pathway and cell death. Semin. Cancer Biol. 1993, 4, 327–332. [Google Scholar] [PubMed]
- Tudor, G.; Aguilera, A.; Halverson, D.; Laing, N.; Sausville, E. Susceptibility to drug-induced apoptosis correlates with differential modulation of Bad, Bcl-2 and Bcl-x L protein levels. Cell Death Differ. 2000, 7, 574–586. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′to3′) | Annealing | Product Size (bp 1) | Accession Number |
|---|---|---|---|---|
| Bcl-2 | Forward: TGGATGACTGAGTACCTGAA Reverse: GGCCTACTGACTTCACTTAT | 59 | 206 | AB116145 |
| Bax | Forward: GGTTGTTGCCCTCCTCTACT Reverse: GTAAGCACTCCAGCCACAAA | 59 | 219 | AB080230 |
| GAPDH | Forward: TGTCCCCACCCCCAATGTATC Reverse: CTCCGATGCCTGCTTCACTACCTT | 59 | 100 | NM 001003142 |
| RPS-19 | Forward: CCTTCCTCAAAAAGTCTGGG Reverse: GTTCTCATCGTAGGGAGCAAG | 59 | 95 | XM 533657.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panja, K.; Buranapraditkun, S.; Roytrakul, S.; Kovitvadhi, A.; Lertwatcharasarakul, P.; Nakagawa, T.; Limmanont, C.; Jaroensong, T. Scorpion Venom Peptide Effects on Inhibiting Proliferation and Inducing Apoptosis in Canine Mammary Gland Tumor Cell Lines. Animals 2021, 11, 2119. https://doi.org/10.3390/ani11072119
Panja K, Buranapraditkun S, Roytrakul S, Kovitvadhi A, Lertwatcharasarakul P, Nakagawa T, Limmanont C, Jaroensong T. Scorpion Venom Peptide Effects on Inhibiting Proliferation and Inducing Apoptosis in Canine Mammary Gland Tumor Cell Lines. Animals. 2021; 11(7):2119. https://doi.org/10.3390/ani11072119
Chicago/Turabian StylePanja, Kamonporn, Supranee Buranapraditkun, Sittiruk Roytrakul, Attawit Kovitvadhi, Preeda Lertwatcharasarakul, Takayuki Nakagawa, Chunsumon Limmanont, and Tassanee Jaroensong. 2021. "Scorpion Venom Peptide Effects on Inhibiting Proliferation and Inducing Apoptosis in Canine Mammary Gland Tumor Cell Lines" Animals 11, no. 7: 2119. https://doi.org/10.3390/ani11072119
APA StylePanja, K., Buranapraditkun, S., Roytrakul, S., Kovitvadhi, A., Lertwatcharasarakul, P., Nakagawa, T., Limmanont, C., & Jaroensong, T. (2021). Scorpion Venom Peptide Effects on Inhibiting Proliferation and Inducing Apoptosis in Canine Mammary Gland Tumor Cell Lines. Animals, 11(7), 2119. https://doi.org/10.3390/ani11072119