
Associative Effects between Forages and Concentrates on In Vitro Fermentation of Working Equine Diets


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Treatments and Experimental Diets
2.2. Ingredients and Chemical Composition
2.3. In Vitro Incubations
2.4. In Vitro GP Kinetics
2.5. Experimental Design and Statistical Analysis
3. Results
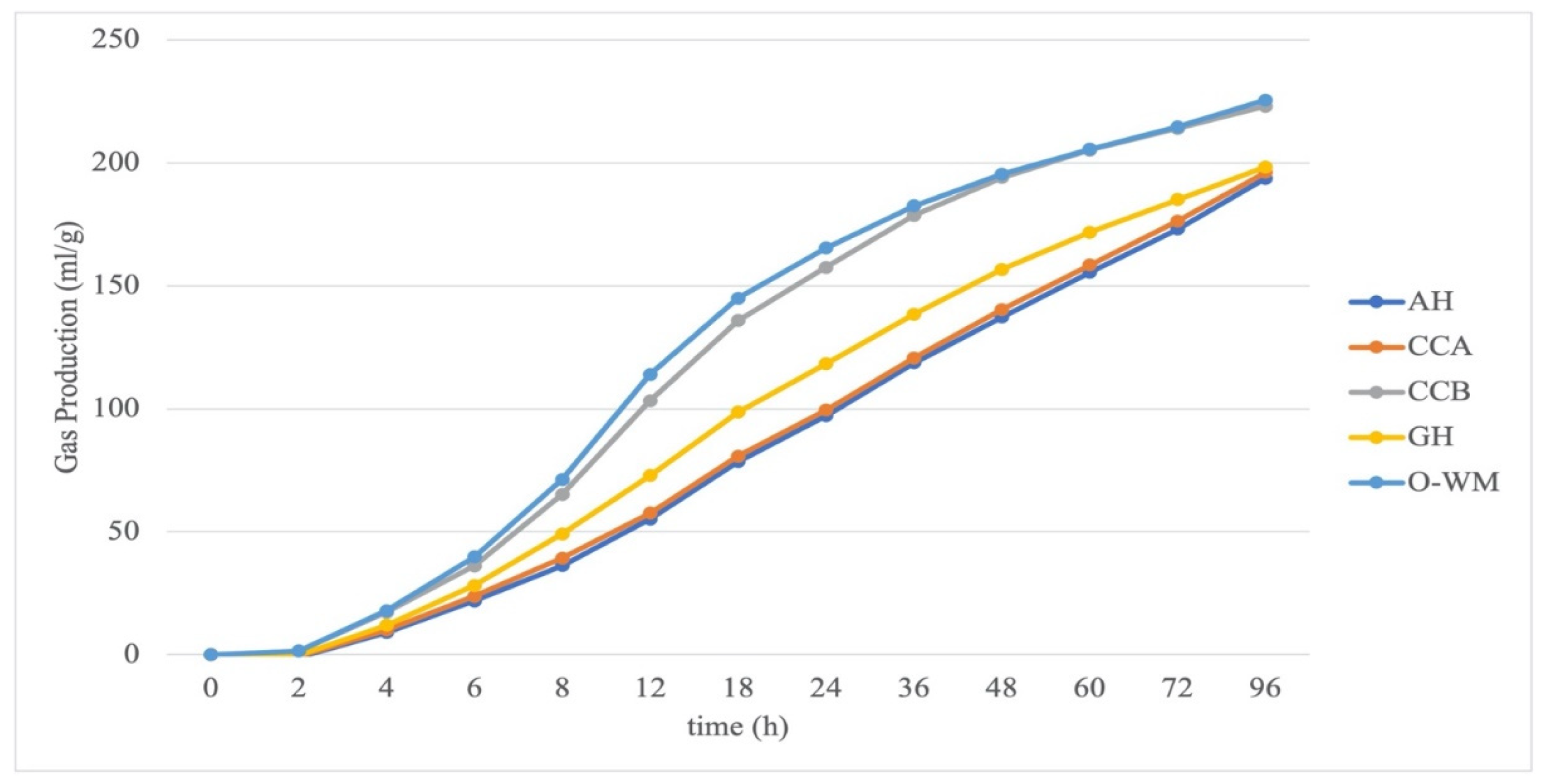
3.1. In Vitro GP and Volatile Fatty Acids from Fermentation of Pure Ingredients
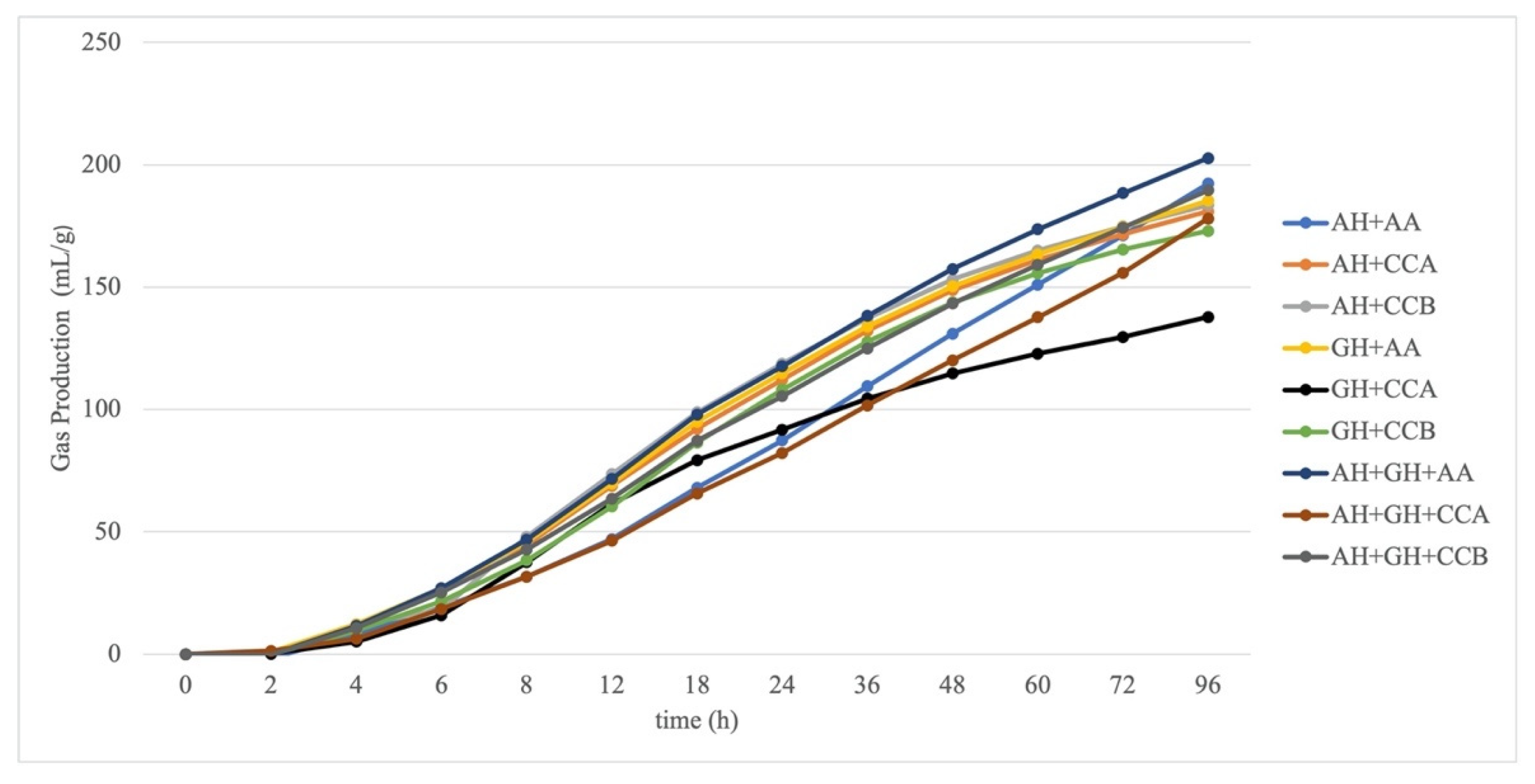
3.2. In Vitro GP Kinetics and Volatile Fatty Acids of Fermentation from the Experimental Diets
4. Discussion
In Vitro Fermentation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leahy, E.R.; Burk, A.O.; Greene, E.A.; Williams, C.A. Nutrition-associated problems facing elite level three-day eventing horses. Equine Vet. J. 2010, 42, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.; Dunnett, C. Nutritional tips for veterinarians. Equine Vet. Educ. 2018, 30, 486–496. [Google Scholar] [CrossRef]
- Bull, J.; Bas, F.; Silva-Guzman, M.; Wentzel, H.H.; Keim, J.P.; Gandarillas, M. Characterization of Feeding, Sport Management, and Routine Care of the Chilean Corralero Horse during Rodeo Season. Animals 2019, 9, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council. Nutrient Requeriments of Horses, 6th revised ed.; The National Academies Press: Washington, DC, USA, 2007; p. 341. [Google Scholar]
- Brunner, J.; Wichert, B.; Burger, D.; von Peinen, K.; Liesegang, A. A survey on the feeding of eventing horses during competition. J. Anim. Physiol. Anim. Nutr. 2012, 96, 878–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.A.; Burk, A. Feeding Management of the Three-Day Event Horse. In Proceedings of the Kentucky Equine Research Nutrition Conference, Lexington, KY, USA, 26–27 April 2010; Kentucky Equine Research: Lexington, KY, USA, 2010. [Google Scholar]
- Brunner, J.; Liesegang, A.; Weiss, S.; Wichert, B. Feeding practice and influence on selected blood parameters in show jumping horses competing in Switzerland. J. Anim. Physiol. Anim. Nutr. 2014, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.; Ellis, A.; Fradinho, M.; Jansson, A.; Julliand, V.; Luthersson, N.; Santos, A.S.; Vervuert, I. Review: Feeding conserved forage to horses: Recent advances and recommendations. Animal 2016, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, B.H.; Flatt, W.P. The Evaluation of Feeds through Digestibility Experiments; University of Georgia Press: Athens, Greece, 1975; p. 423. [Google Scholar]
- Sweeney, C.R. Completion of an In Vivo Digestibility Trials in Horses and In Vitro Digestibility Assay Development; CAL POLY: San Luis Obispo, CA, USA, 2012. [Google Scholar]
- Coles, L.T.; Moughan, P.J.; Darragh, A.J. In vitro digestion and fermentation methods, including gas production techniques, as applied to nutritive evaluation of foods in the hindgut of humans and other simple-stomached animals. Anim. Feed Sci. Technol. 2005, 123–124, 421–444. [Google Scholar] [CrossRef]
- Bush, J.A.; Freeman, D.E.; Kline, K.H.; Merchen, N.R.; Fahey, G.C. Dietary fat supplementation effects on in vitro nutrient disappearance and in vivo nutrient intake and total tract digestibility by horses. J. Anim. Sci. 2001, 79, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Tassone, S.; Fortina, R.; Valle, E.; Cavallarin, L.; Raspa, F.; Boggero, S.; Bergero, D.; Giammarino, M.; Renna, M. Comparison of in vivo and in vitro digestibility in donkeys. Animals 2020, 10, 2100. [Google Scholar] [CrossRef]
- Tassone, S.; Renna, M.; Barbera, S.; Valle, E.; Fortina, R. In vitro digestibility measurement of feedstuffs in donkeys using the daisy(II) incubator. J. Equine Vet. Sci. 2019, 75, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed Sci. Technol. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Earing, J.; Cassill, B.; Hayes, S.; Vanzant, E.; Lawrence, L. Comparison of in vitro digestibility estimates using the Daisy II incubator to in vivo digestibility estimates. J. Anim. Sci. 2010, 88, 3954–3963. [Google Scholar] [CrossRef] [PubMed]
- Abdouli, H.; Attia, S.B. Evaluation of a two-stage in vitro technique for estimating digestibility of equine feeds using horse faeces as the source of microbial inoculum. Anim. Feed Sci. Technol. 2007, 132, 155–162. [Google Scholar] [CrossRef]
- Murray, J.-A.; Scott, B.; Hastie, P. Fermentative capacity of equine faecal inocula obtained from clinically normal horses and those predisposed to laminitis. Anim. Feed Sci. Technol. 2009, 151. [Google Scholar] [CrossRef]
- Lattimer, J.; Cooper, S.; Freeman, D.; Lalman, D. Effect of yeast culture on in vitro fermentation of a high-concentrate or high-fiber diet using equine fecal inoculum in a Daisy II incubator. J. Anim. Sci. 2007, 85, 2484–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ersahince, A.; Kara, K. Nutrient composition and in vitro digestion parameters of Jerusalem artichoke (Helianthus tuberosus L.) herbage at different maturity stages in horse and ruminant. J. Anim. Feed Sci. 2017. [Google Scholar] [CrossRef]
- Elghandour, M.M.Y.; Mellado, M.; Kholif, A.E.; Salem, A.Z.M.; Barbabosa, A.; Ballinas, S.; Esquivel, A.; Odongo, N.E. Fecal Gas Production of Ten Common Horse Feeds Supplemented with Saccharomyces cerevisiae. J. Equine Vet. Sci. 2016, 47, 1–8. [Google Scholar] [CrossRef]
- Elghandour, M.M.Y.; Vázquez Chagoyán, J.C.; Salem, A.Z.M.; Kholif, A.E.; Martínez Castañeda, J.S.; Camacho, L.M.; Buendía, G. In Vitro Fermentative Capacity of Equine Fecal Inocula of 9 fibrous Forages in the Presence of Different Doses of Saccharomyces cerevisiae. J. Equine Vet. Sci. 2014, 34, 619–625. [Google Scholar] [CrossRef]
- Niderkorn, V.; Baumont, R. Associative effects between forages on feed intake and digestion in ruminants. Animal 2009, 3, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.-A.M.D.; Longland, A.; Moore-Colyer, M. In vitro fermentation of different ratios of high-temperature dried lucerne and sugar beet pulp incubated with an equine faecal inoculum. Anim. Feed Sci. Technol. 2006, 129, 89–98. [Google Scholar] [CrossRef]
- Berry, D.P.; Coyne, J.; Coughlan, B.; Burke, M.; McCarthy, J.; Enright, B.; Cromie, A.R.; McParland, S. Genetics of milking characteristics in dairy cows. Animal 2013, 7, 1750–1758. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1996. [Google Scholar]
- Strauch, S.; Wichert, B.; Greef, J.M.; Hillegeist, D.; Zeyner, A.; Liesegang, A. Evaluation of an in vitro system to simulate equine foregut digestion and the influence of acidity on protein and fructan degradation in the horse’s stomach. J. Anim. Physiol. Anim. Nutr. 2017, 101 (Suppl. 1), 51–58. [Google Scholar] [CrossRef] [Green Version]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Groot, J.C.J.; Cone, J.W.; Williams, B.A.; Debersaques, F.M.A.; Lantinga, E.A. Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Oliver, S. SAS for Mixed Models, 2nd ed.; SAS Publishing: Cary, NC, USA, 2006. [Google Scholar]
- Niderkorn, V.; Baumont, R.; Le Morvan, A.; Macheboeuf, D. Occurrence of associative effects between grasses and legumes in binary mixtures on in vitro rumen fermentation characteristics. J. Anim. Sci. 2011, 89, 1138–1145. [Google Scholar] [CrossRef]
- Lowman, R.; Theodorou, M.; Hyslop, J.; Dhanoa, M.; Cuddeford, D. Evulation of an in vitro batch culture technique for estimating the in vivo digestibility and digestible energy content of equine feeds using equine faeces as the source of microbial inoculum. Anim. Feed Sci. Technol. 1999, 80, 11–27. [Google Scholar] [CrossRef]
- Mauricio, R.M.; Owen, E.; Mould, F.L.; Givens, I.; Theodorou, M.K.; France, J.; Davies, D.R.; Dhanoa, M.S. Comparison of bovine rumen liquor and bovine faeces as inoculum for an in vitro gas production technique for evaluating forages. Anim. Feed Sci. Technol. 2001, 89, 33–48. [Google Scholar] [CrossRef]
- Julliand, S.; Martin, A.; Julliand, V. Effect of dehydrated alfalfa on equine gastric and faecal microbial ecosystems. Livest. Sci. 2018, 215, 16–20. [Google Scholar] [CrossRef]
- Moore-Colyer, M.J.S.; Hyslop, J.J.; Longland, A.C.; Cuddeford, D. The mobile bag technique as a method for determining the degradation of four botanically diverse fibrous feedstuffs in the small intestine and total digestive tract of ponies. Br. J. Nutr. 2002, 88, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, J.; Merediz, E.F.-C.; Salmon, K.S.H.; Proudman, C.J.; Edwards, G.B.; Shirazi-Beechey, S.P. Molecular characterisation of carbohydrate digestion and absorption in equine small intestine. Equine Vet. J. 2002, 34, 349–358. [Google Scholar] [CrossRef]
- Murray, J.-A.M.D.; Bice, R.K.T.; Moore-Colyer, M.J.S. The effect of particle size on the in vitro fermentation of different ratios of high-temperature dried lucerne and sugar beet pulp incubated with equine faecal inocula. Anim. Feed Sci. Technol. 2010, 162, 47–57. [Google Scholar] [CrossRef]
- Murray, J.M.D.; Longland, A.; Moore-Colyer, M.; Dunnett, C. The effect of feeding a low- or a high-starch diet on the in vitro fermentative capacity of equine faecal inocula. Anim. Sci. 2006, 82, 627–635. [Google Scholar] [CrossRef]
- Longland, A.C.; Carruthers, J.; Low, A.G. The Ability of Piglets 4 to 8 Weeks Old to Digest and Perform on Diets Containing 2 Contrasting Sources of Nonstarch Polysaccharide. Animal Prod. 1994, 58, 405–410. [Google Scholar] [CrossRef]
- McLean, B.M.L.; Hyslop, J.J.; Longland, A.C.; Cuddeford, D.; Hollands, T. Physical processing of barley and its effects on intra-caecal fermentation parameters in ponies. Anim. Feed Sci. Technol. 2000, 85, 79–87. [Google Scholar] [CrossRef]
- Milinovich, G.J.; Trott, D.J.; Burrell, P.C.; Van Eps, A.W.; Thoefner, M.B.; Blackall, L.L.; Al Jassim, R.A.M.; Morton, J.M.; Pollitt, C.C. Changes in equine hindgut bacterial populations during oligofructose-induced laminitis. Environ. Microbiol. 2006, 8, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Garber, A.; Hastie, P.M.; Handel, I.; Murray, J.M.D. In vitro fermentation of different ratios of alfalfa and starch or inulin incubated with an equine faecal inoculum. Livest. Sci. 2018, 215, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Fehlberg, L.K.; Lattimer, J.M.; Drouillard, J.S.; Douthit, T.L. Effect of Cobalt Chloride on Fermentation of Alfalfa and Smooth Bromegrass Hays by Horse Cecal Microorganisms. J. Equine Vet. Sci. 2019, 77, 75–79. [Google Scholar] [CrossRef]
- Brøkner, C.; Austbø, D.; Næsset, J.A.; Blache, D.; Bach Knudsen, K.E.; Tauson, A.H. Metabolic response to dietary fibre composition in horses. Animal 2016, 10, 1155–1163. [Google Scholar] [CrossRef] [Green Version]
- Philippeau, C.; Sadet-Bourgeteau, S.; Varloud, M.; Julliand, V. Impact of barley form on equine total tract fibre digestibility and colonic microbiota. Animal 2015, 9, 1943–1948. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.H.P.; de Rezende, A.S.C.; da Silva Inácio, D.F. Pectin-rich by-products in feeding horses—A review. Cogent Food Agric. 2016, 2, 1193925. [Google Scholar] [CrossRef]
- Jansson, A.; Lindberg, J.E. A forage-only diet alters the metabolic response of horses in training. Animal 2012, 6, 1939–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollitt, C.C.; Van Eps, A.W. Equine laminitis: A new induction model based on alimentary overload with fructan. In Proceedings of the Bain Fallon Memorial Lectures, Melbourne, Australia, 24–28 July 2002; Australian Equine Veterinary Association: Sydney, Australia, 2002; Volume 24, pp. 96–97. [Google Scholar]
- Wallace, J.; Wallace, S.J.A.; McKain, N.; Nsereko, V.; Hartnell, G. Influence of supplementary fibrolytic enzymes on the fermentation of corn and grass silages by mixed ruminal microorganisms in vitro. J. Anim. Sci. 2001, 79, 1905–1916. [Google Scholar] [CrossRef] [Green Version]
- Hudson, J.M.; Cohen, N.D.; Gibbs, P.G.; Thompson, J.A. Feeding practices associated with colic in horses. J. Am. Vet. Med. Assoc. 2001, 219, 1419–1425. [Google Scholar] [CrossRef]
- Bailey, S.R.; Marr, C.M.; Elliott, J. Current research and theories on the pathogenesis of acute laminitis in the horse. Vet. J. 2004, 167, 129–142. [Google Scholar] [CrossRef]
- Haaland, G.L.; Tyrrell, H.F.; Moe, P.W.; Wheeler, W.E. Effect of Crude Protein Level and Limestone Buffer in Diets Fed at 2 Levels of Intake on Rumen Ph, Ammonia-Nitrogen, Buffering Capacity and Volatile Fatty-Acid Concentration of Cattle. J. Anim. Sci. 1982, 55, 943–950. [Google Scholar] [CrossRef]
- Christensen, H.N. Proteins as buffers. Ann. N. Y. Acad. Sci. 1966, 133, 34–40. [Google Scholar] [CrossRef]
- Marry, R.J.; Jones, R.; Theodorou, M. Grass: Its Production and Utilization, 3rd ed.; Department for Environment, Food and Rural Affairs: Carlisle, UK, 2000; Volume 55.
- Thompson, K.; Jackson, S.; Baker, J. Apparent digestion coefficients and associative effects of varying hay: Grain ratios fed to horses. Nutr. Rep. Int. 1984, 30, 189–197. [Google Scholar]
- Murray, J.M.D.; McMullin, P.; Handel, I.; Hastie, P.M. Comparison of intestinal contents from different regions of the equine gastrointestinal tract as inocula for use in an in vitro gas production technique. Anim. Feed Sci. Technol. 2014, 187, 98–103. [Google Scholar] [CrossRef]
- de Fombelle, A.; Julliand, V.; Drogoul, C.; Jacotot, E. Feeding and microbial disorders in horses: 1-effects of an abrupt incorporation of two levels of barley in a hay diet on microbial profile and activities. J. Equine Vet. Sci. 2001, 21, 439–445. [Google Scholar] [CrossRef]
- Drogoul, C.; de Fombelle, A.; Julliand, V. Feeding and microbial disorders in horses: 2: Effect of three hay:grain ratios on digesta passage rate and digestibility in ponies. J. Equine Vet. Sci. 2001, 21, 487–491. [Google Scholar] [CrossRef]
- Julliand, V.; de Fombelle, A.; Drogoul, C.; Jacotot, E. Feeding and microbial disorders in horses: Part 3—Effects of three hay:grain ratios on microbial profile and activities. J. Equine Vet. Sci. 2001, 21, 543–546. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Ingredient | Dry Matter | Crude Protein | NDF | ADF | Ether Extract | Ash | Ezymatic Digestibility |
|---|---|---|---|---|---|---|---|
| Grass hay | 87.50 | 11.86 | 53.66 | 34.50 | 0.91 | 3.43 | 33.90 |
| Alfalfa hay | 88.50 | 17.65 | 45.81 | 37.02 | 1.32 | 4.32 | 31.20 |
| Comercial concentrate A | 88.50 | 14.61 | 24.61 | 11.21 | 3.82 | 5.35 | 55.50 |
| Comercial concentrate B | 86.50 | 11.31 | 23.88 | 10.73 | 2.07 | 2.69 | 58.90 |
| Oats/wheat middlings | 87.50 | 12.83 | 35.66 | 13.11 | 5.41 | 2.87 | 54.80 |
| Forages | Concentrate | p Value | |||||||
|---|---|---|---|---|---|---|---|---|---|
| AH | GH | AH-GH | CCA | CCB | O-WM | Concentrates v/s Forages | Within Forages | Within Concentrates | |
| A | 220.2 | 252.9 | 236.6 | 257.9 | 224.7 | 220.6 | 0.838 | 0.061 | 0.007 |
| GP96 | 198.3 | 193.7 | 196.0 | 196.1 | 223.0 | 225.5 | 0.023 | 0.641 | 0.005 |
| K | 22.3 | 39.0 | 30.7 | 38.5 | 14.3 | 12.8 | 0.204 | 0.068 | 0.001 |
| C | 0.031 | 0.016 | 0.0235 | 0.017 | 0.06 | 0.07 | 0.046 | 0.0395 | <0.001 |
| MDR | 0.035 | 0.021 | 0.028 | 0.02 | 0.06 | 0.07 | 0.047 | 0.0291 | <0.001 |
| pH | 6.52 | 6.55 | 6.54 | 6.40 | 6.38 | 6.47 | <0.001 | 0.008 | 0.009 |
| VFA | 67.6 | 69.7 | 68.7 | 104.7 | 109.5 | 94.2 | <0.001 | 0.4271 | 0.538 |
| C2 | 58.8 | 59.7 | 59.3 | 55.1 | 57.3 | 57.9 | 0.258 | 0.475 | 0.194 |
| C3 | 23.2 | 23.1 | 23.2 | 26.4 | 26.7 | 23.2 | 0.649 | 0.931 | 0.338 |
| C4 | 18 | 17.2 | 17.6 | 18.4 | 16.2 | 18.4 | 0.423 | 0.39 | 0.194 |
| C2/C3 | 2.54 | 2.58 | 2.56 | 2.08 | 2.14 | 2.43 | 0.022 | 0.805 | 0.339 |
| Alfalfa Hay | Grass Hay | 50% Alfalfa Hay–50% Grass Hay | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CCA | CCB | O-WM | CCA | CCB | O-WM | CCA | CCB | O-WM | p Value | ||||||||||
| A | 196.8 | −29% | 190.9 | −28% | 283.9 | 14% | 136.5 | −70% | 184.9 | −20% | 200.2 | −10% | 268.9 | 10% | 223.6 | −4% | 228.6 | −1% | <0.001 |
| GP96 | 192.3 | −1% | 183.6 | −10% | 192.3 | −6% | 137.8 | −43% | 173 | −19% | 185.33 | −11% | 178 | −10% | 189.6 | −8% | 202.7 | −1% | <0.001 |
| K | 20.6 | −88% | 18.3 | −73% | 51.8 | 40% | 15.6 | −74% | 20.35 | 2% | 20.43 | 5% | 55 | 40% | 27.6 | 7% | 24.1 | −5% | <0.001 |
| C | 0.037 | 56% | 0.047 | 38% | 0.011 | −193% | 0.055 | 51% | 0.04 | 1% | 0.036 | −19% | 0.01 | −116% | 0.022 | −57% | 0.028 | −34% | <0.001 |
| MDR | 0.04 | 48% | 0.048 | 32% | 0.015 | −138% | 0.057 | 46% | 0.042 | −1% | 0.039 | −17% | 0.015 | −71% | 0.028 | −34% | 0.032 | −27% | <0.001 |
| pH | 6.48 | 0% | 6.49 | 0% | 6.49 | −1% | 6.43 | −1% | 6.43 | −1% | 6.41 | −1% | 6.45 | −1% | 6.44 | −1% | 6.46 | −1% | 0.099 |
| VFA | 81.9 | 2% | 87.3 | 6% | 77.3 | 0% | 81.5 | 3% | 85.6 | 6% | 83.1 | 9% | 77.1 | −3% | 77.3 | −5% | 85.4 | 11% | 0.058 |
| C2 | 60.7 | 4% | 62.1 | 5% | 61.3 | 3% | 60.1 | 4% | 62.3 | 6% | 61.6 | 5% | 60 | 3% | 60 | 2% | 63 | 7% | 0.003 |
| C3 | 23.4 | −3% | 22.4 | −8% | 22.7 | −3% | 22.5 | −7% | 22 | −10% | 21.9 | −7% | 23.4 | −3% | 23.4 | −3% | 21.7 | −8% | 0.021 |
| C4 | 15.8 | −11% | 15.5 | −9% | 16 | −10% | 17.4 | −4% | 15.6 | −12% | 16.5 | −10% | 16.6 | −7% | 16.7 | −3% | 15.4 | −16% | 0.001 |
| C2/C3 | 2.6 | 7% | 2.78 | 12% | 2.7 | 6% | 2.68 | 10% | 2.84 | 15% | 2.81 | 11% | 2.56 | 6% | 2.56 | 5% | 2.9 | 13% | 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandarillas, M.; Keim, J.P.; Gapp, E.M. Associative Effects between Forages and Concentrates on In Vitro Fermentation of Working Equine Diets. Animals 2021, 11, 2212. https://doi.org/10.3390/ani11082212
Gandarillas M, Keim JP, Gapp EM. Associative Effects between Forages and Concentrates on In Vitro Fermentation of Working Equine Diets. Animals. 2021; 11(8):2212. https://doi.org/10.3390/ani11082212
Chicago/Turabian StyleGandarillas, Mónica, Juan Pablo Keim, and Elisa María Gapp. 2021. "Associative Effects between Forages and Concentrates on In Vitro Fermentation of Working Equine Diets" Animals 11, no. 8: 2212. https://doi.org/10.3390/ani11082212
APA StyleGandarillas, M., Keim, J. P., & Gapp, E. M. (2021). Associative Effects between Forages and Concentrates on In Vitro Fermentation of Working Equine Diets. Animals, 11(8), 2212. https://doi.org/10.3390/ani11082212