
Using the Relationship between Concentrations of Selected Whey Proteins and BHBA to Characterize the Metabolism of Dairy Cows in Early Lactation

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Chemical Analyses
2.3. Statistical Analysis
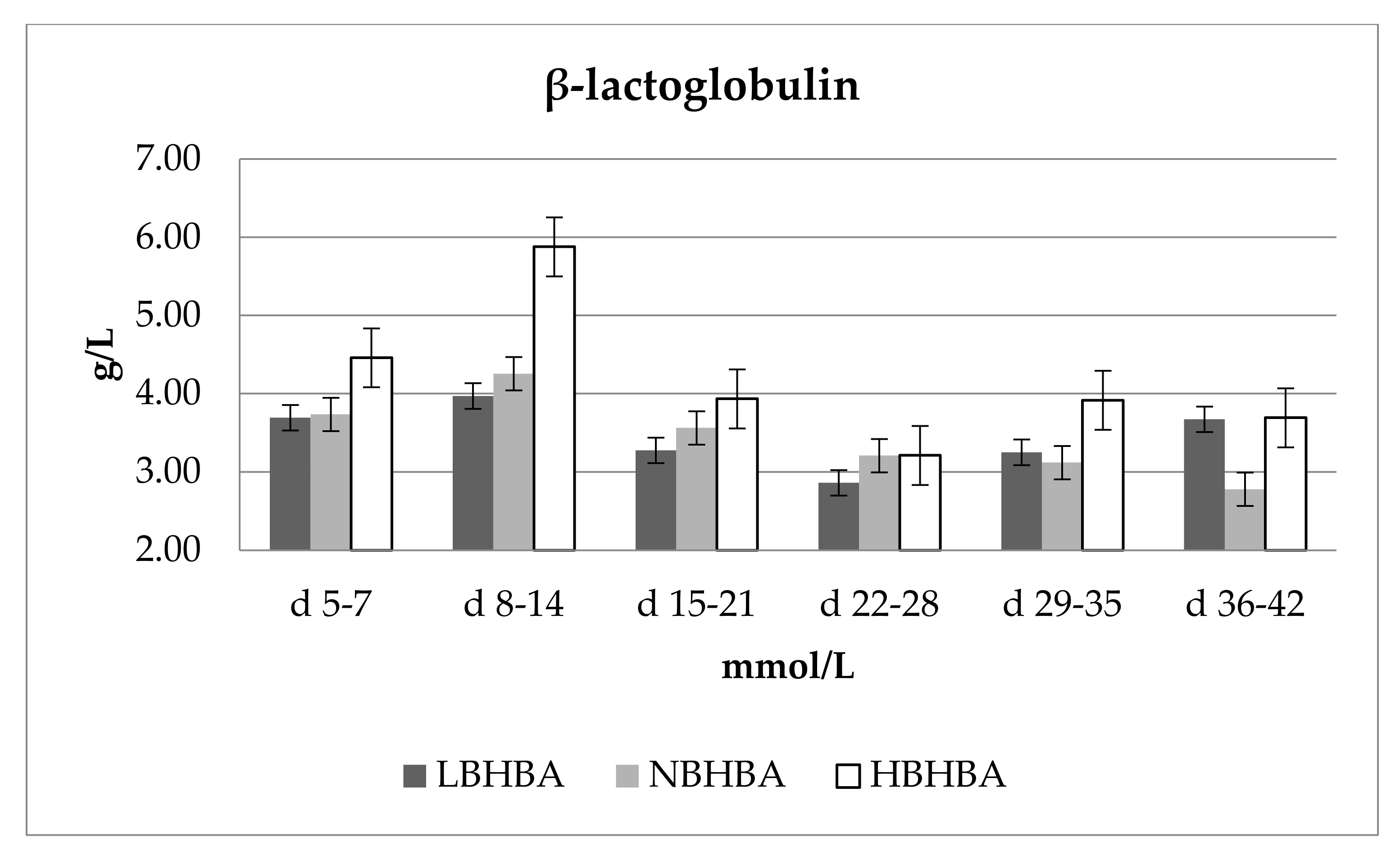
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Puppel, K.; Kuczyńska, B. Metabolic profiles of cow’s blood; a review. J. Sci. Food Agric. 2016, 96, 4321–4328. [Google Scholar] [CrossRef]
- Gołębiewski, M. Badanie Przydatności Zmodyfikowanej Oceny Kondycji Krów Mlecznych w Zarządzaniu Ich Stadem ze Szczególnym Uwzględnieniem Parametrów Produkcyjnych, Funkcjonowania Układu Rozrodczego Oraz Zdrowia Zwierząt; Wydawnictwo SGGW: Warsaw, Poland, 2017. [Google Scholar]
- Oetzel, G.R. Herd-level ketosis–diagnosis and risk factors. In Proceedings of the 40th Annual Conference of Bovine Practitioners, Vancouver, BC, Canada, 19 September 2007; Citeseer: Princeton, NJ, USA, 2021. Available online: https://www.vetmed.wisc.edu/dms/fapm/fapmtools/2nutr/ketosis.pdf (accessed on 11 February 2021).
- van der Drift, S.G.A.; Houweling, M.; Schonewille, J.T.; Tielens, A.G.M.; Jorritsma, R. Protein and fat mobilization and associations with serum beta-hydroxybutyrate concentrations in dairy cows. J. Dairy Sci. 2012, 95, 4911–4920. [Google Scholar] [CrossRef] [PubMed]
- Kanikarla-Marie, P.; Jain, S.K. Hyperketonemia and ketosis increase the risk of complications in type 1 diabetes. Free Radic. Biol. Med. 2016, 95, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Puppel, K.; Gołębiewski, M.; Solarczyk, P.; Grodkowski, G.; Slósarz, J.; Kunowska-Slósarz, M.; Balcerak, M.; Przysucha, T.; Kalińska, A.; Kuczyńska, B. The relationship between plasma β-hydroxybutyric acid and conjugated linoleic acid in milk as a biomarker for early diagnosis of ketosis in postpartum Polish Holstein-Friesian cows. BMC Vet. Res. 2019, 367. [Google Scholar] [CrossRef] [Green Version]
- Ospina, P.A.; Nydam, D.V.; Stokol, T.; Overton, T.R. Associations of elevated nonesterified fatty acids and beta-hydroxybutyrate concentrations with early lactation reproductive performance and milk production in transition dairy cattle in the northeastern United States. J. Dairy Sci. 2010, 93, 1596–1603. [Google Scholar] [CrossRef] [Green Version]
- Krause, K.; Oetzel, G. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Tech. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Penner, G.; Oba, M.; Gäbel, G.; Aschenbach, J. A single mild episode of subacute ruminal acidosis does not affect ruminal barrier function in short term. J. Dairy Sci. 2010, 93, 4838–4845. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Lopez, E.; Wiese, B.I.; Hendrick, S.; McKinnon, J.J.; McAllister, T.A.; Beauchemin, K.A.; Penner, G.B. Incidence, prevalence, severity and risk factors for ruminal acidosis in feedlot steers during back grounding diet transition, and finishing. J. Anim. Sci. 2014, 92, 3053–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leighton, B.; Nicholas, A.R.; Pogson, C.I. The pathway of ketogenesis in rumen epithelium of the sheep. Biochem. J. 1983, 216, 769–772. [Google Scholar] [CrossRef] [Green Version]
- Meertens, L.M.; Miyata, K.S.; Cechetto, J.D.; Rachubinski, R.A.; Capone, J.P. A mitochondrial ketogenic enzyme regulates its gene expression by association with the nuclear hormone receptor PPARα. EMBO J. 1998, 17, 6972–6978. [Google Scholar] [CrossRef] [Green Version]
- Owens, F.N.; Secrist, D.S.; Hill, W.J.; Gill, D. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Enemark, J.M. The monitoring, prevention and treatment of sub-acute ruminal acidosis (SARA): A review. Vet. J. 2008, 176, 32–43. [Google Scholar] [CrossRef] [PubMed]
- IBM Crop: Released IBM SPSS for Windows, Version 23.0; IBM Crop: Armonk, NY, USA, 2021.
- Bionaz, M.; Periasamy, K.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Hurley, W.L.; Loor, J.J. Old and new stories: Revelations from functional analysis of the bovine mammary transcriptome during the lactation cycle. PLoS ONE 2012, 7, e33268. [Google Scholar] [CrossRef]
- Griinari, J.M.; McGuire, M.A.; Dwyer, D.A.; Bauman, D.E.; Barbano, D.M.; House, W.A. The role of insulin in the regulation of milk protein synthesis in dairy cows. J. Dairy Sci. 1997, 80, 2361–2371. [Google Scholar] [CrossRef]
- Underwood, W.J.; Blauwiekel, R.; Delano, M.L.; Gillesby, R.; Mischler, S.A.; Schoell, A. Chapter 15—Biology and Diseases of Ruminants (Sheep, Goats, and Cattle). In American College of Laboratory Animal Medicine, Laboratory Animal Medicine, 3rd ed.; James, G.F., Lynn, C.A., Glen, M.O., Kathleen, R., Pritchett-Corning, M.T.W., Eds.; Academic Press: Cambridge, MA, USA, 2015; ISBN 9780124095274. [Google Scholar]
- Duffield, T.F.; Lissemore, K.D.; McBride, B.W.; Leslie, K.E. Impact of hyperketonemia in early lactation dairy cows on health and production. J. Dairy Sci. 2009, 92, 571–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieve, D.G.; Korver, S.; Rijpkema, Y.S.; Hof, G. Relationship between milk composition and some nutritional parameters in early lactation. Livest. Prod. 1986, 14, 239–254. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, S.; Hui, Q.; Lei, L.; Du, X.; Gao, W.; Zhang, R.; Liu, G.; Li, X. β-Hydroxybutyrate Facilitates Fatty Acids Synthesis Mediated by Sterol Regulatory Element-Binding Protein1 in Bovine Mammary Epithelial Cells. Cell Physiol. Biochem. 2015, 37, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Månsson, H.L. Fatty acids in bovine milk fat. Food Nutr. Res. 2008, 52, 1821. [Google Scholar] [CrossRef] [Green Version]
- Schimmel, P.; Alexander, R.W. Protein Synthesis. Encyclopedia of Physical Science and Technology, 3rd ed.; Robert, A.M., Ed.; Academic Press: Cambridge, MA, USA, 2003; ISBN 9780122274107. [Google Scholar] [CrossRef]
- Cozzi, G.; Ravarotto, L.; Gottardo, F.; Stefani, A.L.; Contiero, B.; Moro, L.; Brscic, M.; Dalvit, P. Short communication: Reference values for blood parameters in Holstein dairy cows: Effects of parity, stage of lactation, and season of production. J. Dairy Sci. 2011, 94, 3895–3901. [Google Scholar] [CrossRef]
- Gellrich, K.; Meyer, H.H.D.; Wiedemann, S. Composition of major proteins in cow milk differing in mean protein concentration during the first 155 days of lactation and the influence of season as well as short-term restricted feeding in early and mid-lactation. Czech J. Anim. Sci. 2014, 59, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Barbana, C.; Sanchez, L.; Perez, M.D. Bioactivity of alpha-lactalbumin related to its interaction with fatty acids: A review. Crit. Rev. Food Sci. Nutr. 2011, 51, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Nasrollahi, S.M.; Zali, A.; Ghorbani, G.R.; Kahyani, A.; Beauchemin, K.A. Short communication: Blood metabolites, body reserves, and feed efficiency of high-producing dairy cows that varied in ruminal pH when fed a high-concentrate diet. J. Dairy Sci. 2019, 102, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.D.; Calvo, M. Interaction of beta-lactoglobulin with retinol and fatty acids and its role as a possible biological function for this protein: A review. J. Dairy Sci. 1995, 78, 978–988. [Google Scholar] [CrossRef]
- Considine, T.; Patel, H.A.; Singh, H.; Creamer, L.K. Influence of binding conjugated linoleic acid and myristic acid on the heat-and high-pressure-induced unfolding and aggregation of beta-lactoglobulin B. Food Chem. 2007, 102, 1270–1280. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Acidosis (n = 30) | Healthy (n = 37) | Ketosis (n = 28) | ||||
|---|---|---|---|---|---|---|
| LSM | SEM | LSM | SEM | LSM | SEM | |
| Milk Yield (kg/d) | 31.221 | 1.005 | 29.927 | 1.024 | 27.115 | 0.902 |
| Glucose (mg/dL) | 58.223 | 0.945 | 63.025 | 0.887 | 39.011 | 0.862 |
| GGTP (U/L) | 27.255 | 0.521 | 25.027 | 0.668 | 72.036 | 0.754 |
| AspAT (U/L) | 127.215 | 1.114 | 63.061 | 0.847 | 64.052 | 0.825 |
| BHBA (mmol/L) | 0.200–0.500 | 0.012 | 0.510–1.200 | 0.014 | > 1.200 | 0.012 |
| F/P | 1.058 | 0.071 | 1.212 | 0.065 | 1.625 | 0.045 |
| Days in Milk | BHBA Group | Casein (%) | Protein (%) | Fat (%) | Lactose (%) | Interaction DIM × BHBA | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| LSM | SE | LSM | SE | LSM | SE | LSM | SE | |||
| d 5–7 | LBHBA | 3.54 | 0.152 | 4.51 | 0.196 | 4.26 | 0.687 | 3.93 | 0.186 | |
| NBHBA | 3.07 | 0.058 | 3.83 | 0.074 | 4.84 | 0.26 | 4.48 | 0.07 | p ≤ 0.01 | |
| HBHBA | 2.69 | 0.076 | 3.18 | 0.098 | 6.6 | 0.344 | 4.35 | 0.093 | ||
| d 8–14 | LBHBA | 2.85 | 0.068 | 3.52 | 0.088 | 3.13 | 0.307 | 4.64 | 0.083 | |
| NBHBA | 2.96 | 0.037 | 3.61 | 0.047 | 4.64 | 0.167 | 4.74 | 0.045 | p ≤ 0.01 | |
| HBHBA | 2.79 | 0.068 | 3.42 | 0.088 | 5.39 | 0.307 | 4.6 | 0.083 | ||
| d 15–21 | LBHBA | 2.72 | 0.051 | 3.32 | 0.065 | 3.25 | 0.229 | 4.79 | 0.062 | p ≤ 0.01 |
| NBHBA | 2.75 | 0.037 | 3.28 | 0.047 | 4.34 | 0.167 | 4.9 | 0.045 | ||
| HBHBA | 2.57 | 0.076 | 3.02 | 0.098 | 5.59 | 0.344 | 4.78 | 0.093 | ||
| d 22–28 | LBHBA | 2.67 | 0.068 | 3.11 | 0.088 | 3.22 | 0.307 | 5.01 | 0.083 | |
| NBHBA | 2.55 | 0.048 | 2.98 | 0.062 | 4.7 | 0.217 | 4.93 | 0.059 | p ≤ 0.01 | |
| HBHBA | 2.58 | 0.108 | 2.94 | 0.138 | 5.48 | 0.486 | 4.99 | 0.131 | ||
| d 29–35 | LBHBA | 2.48 | 0.088 | 2.95 | 0.113 | 3.76 | 0.397 | 4.92 | 0.107 | |
| NBHBA | 2.55 | 0.031 | 3.02 | 0.04 | 3.74 | 0.14 | 4.96 | 0.038 | p ≤ 0.01 | |
| HBHBA | 2.3 | 0.051 | 2.73 | 0.065 | 4.87 | 0.229 | 4.73 | 0.062 | ||
| d 36–42 | LBHBA | 2.65 | 0.108 | 3.15 | 0.138 | 4.23 | 0.486 | 4.87 | 0.131 | |
| NBHBA | 2.56 | 0.032 | 3.07 | 0.041 | 3.8 | 0.143 | 4.83 | 0.039 | p ≤ 0.01 | |
| HBHBA | 2.44 | 0.108 | 2.9 | 0.138 | 4.23 | 0.486 | 4.85 | 0.131 | ||
| Fat | Protein | Lactose | BHBA | α-LA | β-LG | |
|---|---|---|---|---|---|---|
| Fat | 1 | 0.220 ** | −0.198 ** | 0.300 ** | −0,136 | 0.241 ** |
| Protein | 0.220 ** | 1 | −0.495 ** | −0.310 ** | 0.178 * | −0.211 ** |
| Lactose | −0.198 ** | −0.495 ** | 1 | 0.219 ** | −0.233 ** | 0.178 * |
| BHBA | 0.300 ** | −0.310 ** | 0.219 ** | 1 | −0.695 ** | 0.757 ** |
| α-LA | −0,136 | 0.178 * | −0.233 ** | −0.695 ** | 1 | −0.615 ** |
| β-LG | 0.241 ** | −0.211 ** | 0.178 * | 0.757 ** | −0.615 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puppel, K.; Staniszewska, P.; Gołębiewski, M.; Slósarz, J.; Grodkowski, G.; Solarczyk, P.; Kunowska-Slósarz, M.; Kostusiak, P.; Kuczyńska, B.; Przysucha, T. Using the Relationship between Concentrations of Selected Whey Proteins and BHBA to Characterize the Metabolism of Dairy Cows in Early Lactation. Animals 2021, 11, 2298. https://doi.org/10.3390/ani11082298
Puppel K, Staniszewska P, Gołębiewski M, Slósarz J, Grodkowski G, Solarczyk P, Kunowska-Slósarz M, Kostusiak P, Kuczyńska B, Przysucha T. Using the Relationship between Concentrations of Selected Whey Proteins and BHBA to Characterize the Metabolism of Dairy Cows in Early Lactation. Animals. 2021; 11(8):2298. https://doi.org/10.3390/ani11082298
Chicago/Turabian StylePuppel, Kamila, Patrycja Staniszewska, Marcin Gołębiewski, Jan Slósarz, Grzegorz Grodkowski, Paweł Solarczyk, Małgorzata Kunowska-Slósarz, Piotr Kostusiak, Beata Kuczyńska, and Tomasz Przysucha. 2021. "Using the Relationship between Concentrations of Selected Whey Proteins and BHBA to Characterize the Metabolism of Dairy Cows in Early Lactation" Animals 11, no. 8: 2298. https://doi.org/10.3390/ani11082298