
The First Report of Genetic Polymorphisms of the Equine SPRN Gene in Outbred Horses, Jeju and Halla Horses



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals
2.3. Genetic Analysis
2.4. mRNA Structural Analysis
2.5. Literature Search
2.6. Statistical Analysis
3. Results
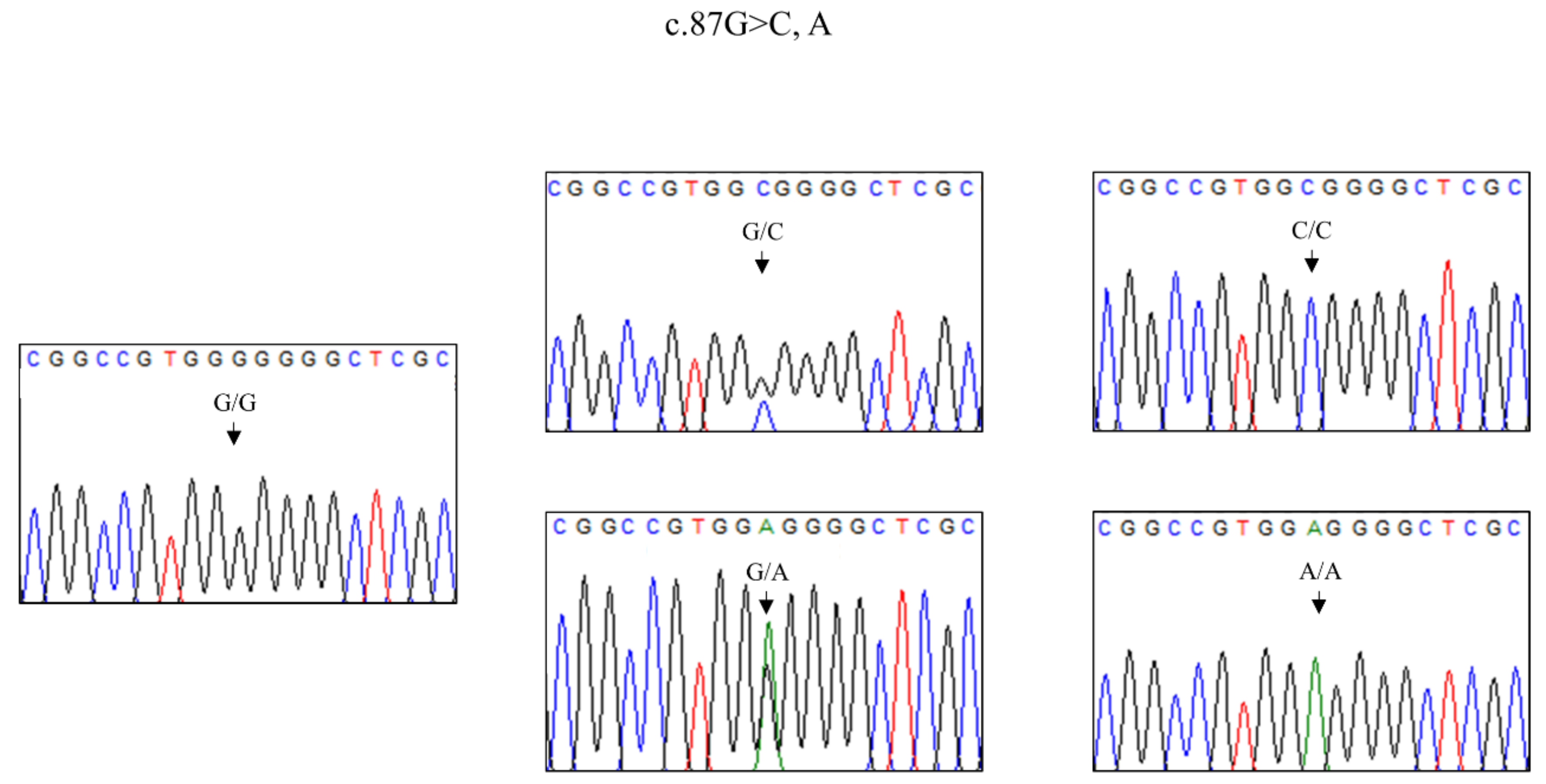
3.1. Comparative Analysis of Genotype and Allele Frequencies of the SPRN Gene among Three Horse Species
3.2. Haplotype Analysis of the SPRN SNPs in Three Horses
3.3. LD Analysis of SPRN Polymorphisms in Jeju and Halla Horses
3.4. mRNA Secondary Structure Analysis of the Equine SPRN Gene According to the Polymorphisms
3.5. The Number of Polymorphisms of the SPRN Gene in Prion Disease-Resistant Animals and Prion Disease-Susceptible Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PrPC | Endogenous prion protein |
| PrPSc | Pathogenic prion protein |
| PRNP | Prion protein gene |
| PRND | Prion-like protein gene |
| PRNT | Prion-related protein gene |
| SPRN | Shadow of prion protein gene |
| LD | Linkage disequilibrium |
| GPI | Glycosylphosphatidylinositol |
| vCJD | Variant Creutzfeldt-Jakob disease |
| UTR | Untranslated region |
| BSE | Bovine spongiform encephalopathy |
| SNPs | Single nucleotide polymorphisms |
| ORF | Open reading frame |
References
- Prusiner, S.B. The prion diseases. Brain Pathol. 1998, 8, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, S.B. Prions. Proc. Natl. Acad. Sci. USA 1998, 95, 13363–13383. [Google Scholar] [CrossRef] [Green Version]
- Gambetti, P.; Cali, I.; Notari, S.; Kong, Q.; Zou, W.Q.; Surewicz, W.K. Molecular biology and pathology of prion strains in sporadic human prion diseases. Acta Neuropathol. 2011, 121, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Cassmann, E.D.; Greenlee, J.J. Pathogenesis, detection, and control of scrapie in sheep. Am. J. Vet. Res. 2020, 81, 600–614. [Google Scholar] [CrossRef]
- Won, S.Y.; Kim, Y.C.; Jeong, B.H. First Report of the Potential Bovine Spongiform Encephalopathy (BSE)-Related Somatic Mutation E211K of the Prion Protein Gene (PRNP) in Cattle. Int. J. Mol. Sci. 2020, 21, 4246. [Google Scholar] [CrossRef]
- Scheckel, C.; Aguzzi, A. Prions, prionoids and protein misfolding disorders. Nat. Rev. Genet. 2018, 19, 405–418. [Google Scholar] [CrossRef] [Green Version]
- Baylis, M.; Goldmann, W. The genetics of scrapie in sheep and goats. Curr. Mol. Med. 2004, 4, 385–396. [Google Scholar] [CrossRef]
- Lloyd, S.; Mead, S.; Collinge, J. Genetics of prion disease. Top. Curr. Chem. 2011, 305, 1–22. [Google Scholar] [CrossRef]
- Murdoch, B.M.; Murdoch, G.K. Genetics of Prion Disease in Cattle. Bioinform. Biol. Insights 2015, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Jeong, B.H. First report of prion-related protein gene (PRNT) polymorphisms in cattle. Vet. Rec. 2018, 182, 717. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, B.H. The first report of prion-related protein gene (PRNT) polymorphisms in goat. Acta Vet. Hung. 2017, 65, 291–300. [Google Scholar] [CrossRef]
- Roh, I.S.; Kim, Y.C.; Kim, H.J.; Won, S.Y.; Jeong, M.J.; Kang, H.E.; Sohn, H.J.; Jeong, B.H. Identification of the prion-related protein gene (PRNT) sequences in various species of the Cervidae family. Mol. Biol. Rep. 2020, 47, 6155–6164. [Google Scholar] [CrossRef]
- Won, S.Y.; Kim, Y.C.; Do, K.; Jeong, B.H. Absence of Strong Genetic Linkage Disequilibrium between Single Nucleotide Polymorphisms (SNPs) in the Prion Protein Gene (PRNP) and the Prion-Like Protein Gene (PRND) in the Horse, a Prion-Resistant Species. Genes 2020, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Kim, S.K.; Won, S.Y.; Jeong, B.H. Polymorphisms of shadow of prion protein gene (SPRN) in Korean native cattle (Hanwoo) and Holstein cattle. Sci. Rep. 2020, 10, 15272. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kim, S.K.; Jeong, B.H. Scrapie susceptibility-associated indel polymorphism of shadow of prion protein gene (SPRN) in Korean native black goats. Sci. Rep. 2019, 9, 15261. [Google Scholar] [CrossRef] [Green Version]
- Onodera, T.; Nishimura, T.; Sugiura, K.; Sakudo, A. Function of Prion Protein and the Family Member, Shadoo. Curr. Issues Mol. Biol. 2020, 36, 67–88. [Google Scholar] [CrossRef]
- Westaway, D.; Daude, N.; Wohlgemuth, S.; Harrison, P. The PrP-like proteins Shadoo and Doppel. Top. Curr. Chem. 2011, 305, 225–256. [Google Scholar] [CrossRef]
- Ciric, D.; Richard, C.A.; Moudjou, M.; Chapuis, J.; Sibille, P.; Daude, N.; Westaway, D.; Adrover, M.; Beringue, V.; Martin, D.; et al. Interaction between Shadoo and PrP Affects the PrP-Folding Pathway. J. Virol. 2015, 89, 6287–6293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, J.A.; Campbell, T.A.; Adamson, G.; Poulter, M.; Uphill, J.B.; Molou, E.; Collinge, J.; Mead, S. Association of a null allele of SPRN with variant Creutzfeldt-Jakob disease. J. Med. Genet. 2008, 45, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Peletto, S.; Bertolini, S.; Maniaci, M.G.; Colussi, S.; Modesto, P.; Biolatti, C.; Bertuzzi, S.; Caramelli, M.; Maurella, C.; Acutis, P.L. Association of an indel polymorphism in the 3’UTR of the caprine SPRN gene with scrapie positivity in the central nervous system. J. Gen. Virol. 2012, 93, 1620–1623. [Google Scholar] [CrossRef]
- Gurgul, A.; Polak, M.P.; Larska, M.; Slota, E. PRNP and SPRN genes polymorphism in atypical bovine spongiform encephalopathy cases diagnosed in Polish cattle. J. Appl. Genet. 2012, 53, 337–342. [Google Scholar] [CrossRef]
- Won, S.Y.; Kim, Y.C.; Kim, S.K.; Jeong, B.H. The First Report of Genetic and Structural Diversities in the SPRN Gene in the Horse, an Animal Resistant to Prion Disease. Genes 2019, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- McGivney, B.A.; Han, H.; Corduff, L.R.; Katz, L.M.; Tozaki, T.; MacHugh, D.E.; Hill, E.W. Genomic inbreeding trends, influential sire lines and selection in the global Thoroughbred horse population. Sci. Rep. 2020, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Daude, N.; Westaway, D. Biological properties of the PrP-like Shadoo protein. Front. Biosci. 2011, 16, 1505–1516. [Google Scholar] [CrossRef]
- Lampo, E.; Duchateau, L.; Schepens, B.; Van Poucke, M.; Saelens, X.; Erkens, T.; Van Zeveren, A.; Peelman, L.J. Identification of polymorphisms in the ovine Shadow of prion protein (SPRN) gene and assessment of their effect on promoter activity and susceptibility for classical scrapie. Anim. Genet. 2010, 41, 169–178. [Google Scholar] [CrossRef]
- Stewart, P.; Shen, C.; Zhao, D.; Goldmann, W. Genetic analysis of the SPRN gene in ruminants reveals polymorphisms in the alanine-rich segment of shadoo protein. J. Gen. Virol. 2009, 90, 2575–2580. [Google Scholar] [CrossRef]
- Allais-Bonnet, A.; Pailhoux, E. Role of the prion protein family in the gonads. Front. Cell Dev. Biol. 2014, 2, 56. [Google Scholar] [CrossRef] [Green Version]
- Makzhami, S.; Passet, B.; Halliez, S.; Castille, J.; Moazami-Goudarzi, K.; Duchesne, A.; Vilotte, M.; Laude, H.; Mouillet-Richard, S.; Beringue, V.; et al. The prion protein family: A view from the placenta. Front. Cell Dev. Biol. 2014, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Won, S.Y.; Jeong, B.H. Absence of single nucleotide polymorphisms (SNPs) in the open reading frame (ORF) of the prion protein gene (PRNP) in a large sampling of various chicken breeds. BMC Genom. 2019, 20, 922. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, B.H. The first report of polymorphisms and genetic characteristics of the prion protein gene (PRNP) in horses. Prion 2018, 12, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, S.Y.; Kim, Y.C.; Kim, K.; Kim, A.D.; Jeong, B.H. The First Report of Polymorphisms and Genetic Features of the prion-like Protein Gene (PRND) in a Prion Disease-Resistant Animal, Dog. Int. J. Mol. Sci. 2019, 20, 1404. [Google Scholar] [CrossRef] [Green Version]
- Im, E.H.; Choi, S.S. Synonymous Codon Usage Controls Various Molecular Aspects. Genom. Inform. 2017, 15, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Didiano, D.; Hobert, O. Molecular architecture of a miRNA-regulated 3′ UTR. RNA 2008, 14, 1297–1317. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymorphisms | Breeds | Total, n | Allele Frequency, n (%) | p-Value | References | ||
|---|---|---|---|---|---|---|---|
| c.87G > C | G | C | A | ||||
| Thoroughbred | 194 | 382 (98.5) | 6 (1.5) | 0 (0) | <0.0001 | [22] | |
| Halla | 88 | 163 (92.6) | 4 (2.3) | 9 (5.1) | <0.0001 | Current study | |
| Jeju | 141 | 150 (53.2) | 95 (33.7) | 37 (13.1) | - | Current study | |
| c.502C > T | C | T | |||||
| Thoroughbred | 194 | 361 (93.1) | 27 (6.9) | <0.0001 | [22] | ||
| Halla | 88 | 143 (81.3) | 33 (18.7) | <0.0001 | Current study | ||
| Jeju | 141 | 137 (48.6) | 145 (51.4) | - | Current study | ||
| c.696C > T | C | T | |||||
| Thoroughbred | 194 | 359 (92.5) | 29 (7.5) | <0.0001 | [22] | ||
| Halla | 88 | 143 (81.3) | 33 (18.7) | <0.0001 | Current study | ||
| Jeju | 141 | 137 (48.6) | 145 (51.4) | - | Current study | ||
| c.728C > T | C | T | |||||
| Thoroughbred | 194 | 270 (69.6) | 118 (30.4) | <0.0001 | [22] | ||
| Halla | 88 | 94 (53.4) | 82 (46.6) | <0.0001 | Current study | ||
| Jeju | 141 | 72 (25.5) | 210 (74.5) | - | Current study | ||
| Haplotype | Frequency | p-Value 1 | p-Value 2 | ||
|---|---|---|---|---|---|
| Jeju (n = 282) | Halla (n = 176) | Thoroughbred (n = 388) | |||
| CTTT | 85 (0.301) | 9 (0.051) | 6 (0.015) | <0.0001 | <0.0001 |
| GCCT | 60 (0.213) | 53 (0.301) | 105 (0.271) | 0.0997 | 0.0862 |
| GCCC | 38 (0.135) | 76 (0.432) | 251 (0.647) | <0.0001 | <0.0001 |
| GTTT | 24 (0.085) | 15 (0.085) | 5 (0.013) | 0.9967 | <0.0001 |
| ACCT | 23 (0.082) | 9 (0.051) | 0 (0) | 0.2451 | <0.0001 |
| ATTT | 14 (0.050) | 0 (0) | 0 (0) | 0.0015 | <0.0001 |
| CTTC | 13 (0.046) | 0 (0) | 0 (0) | 0.0026 | <0.0001 |
| GTTC | 13 (0.046) | 4 (0.023) | 14 (0.036) | 0.3092 | 0.5151 |
| Others | 12 (0.042) | 10 (0.057) | 7 (0.018) | - | |
| Jeju | Halla | |||||||
|---|---|---|---|---|---|---|---|---|
| r2 | c.87G > C (Codon 29) | c.502C > T (Codon 168) | c.696C > T (Codon 232) | c.728C > T (Codon 243) | c.87G > C (Codon 29) | c.502C > T (Codon 168) | c.696C > T (Codon 232) | c.728C > T (Codon 243) |
| c.87G > C (codon 29) | - | - | - | - | - | - | - | - |
| c.502C > T (codon 168) | 0.543 | - | - | - | 0.098 | - | - | - |
| c.696C > T (codon 232) | 0.543 | 1.0 | - | - | 0.098 | 1.0 | - | - |
| c.728C > T (codon 243) | 0.072 | 0.052 | 0.052 | - | 0.033 | 0.01 | 0.01 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, S.-Y.; Kim, Y.-C.; Do, K.; Jeong, B.-H. The First Report of Genetic Polymorphisms of the Equine SPRN Gene in Outbred Horses, Jeju and Halla Horses. Animals 2021, 11, 2574. https://doi.org/10.3390/ani11092574
Won S-Y, Kim Y-C, Do K, Jeong B-H. The First Report of Genetic Polymorphisms of the Equine SPRN Gene in Outbred Horses, Jeju and Halla Horses. Animals. 2021; 11(9):2574. https://doi.org/10.3390/ani11092574
Chicago/Turabian StyleWon, Sae-Young, Yong-Chan Kim, Kyoungtag Do, and Byung-Hoon Jeong. 2021. "The First Report of Genetic Polymorphisms of the Equine SPRN Gene in Outbred Horses, Jeju and Halla Horses" Animals 11, no. 9: 2574. https://doi.org/10.3390/ani11092574