
Dry Period Heat Stress Impacts Mammary Protein Metabolism in the Subsequent Lactation

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Physiological Measures and Milk Yield
2.3. Milk and Blood Profile Analysis
2.4. Mammary Biopsies
2.5. RNA Isolation and qRT-PCR
2.6. Protein Extraction and Western Blotting
2.7. Mammary Tissue AA Concentration
2.8. Statistical Analyses
3. Results
3.1. Physiological Measures and Milk Yield
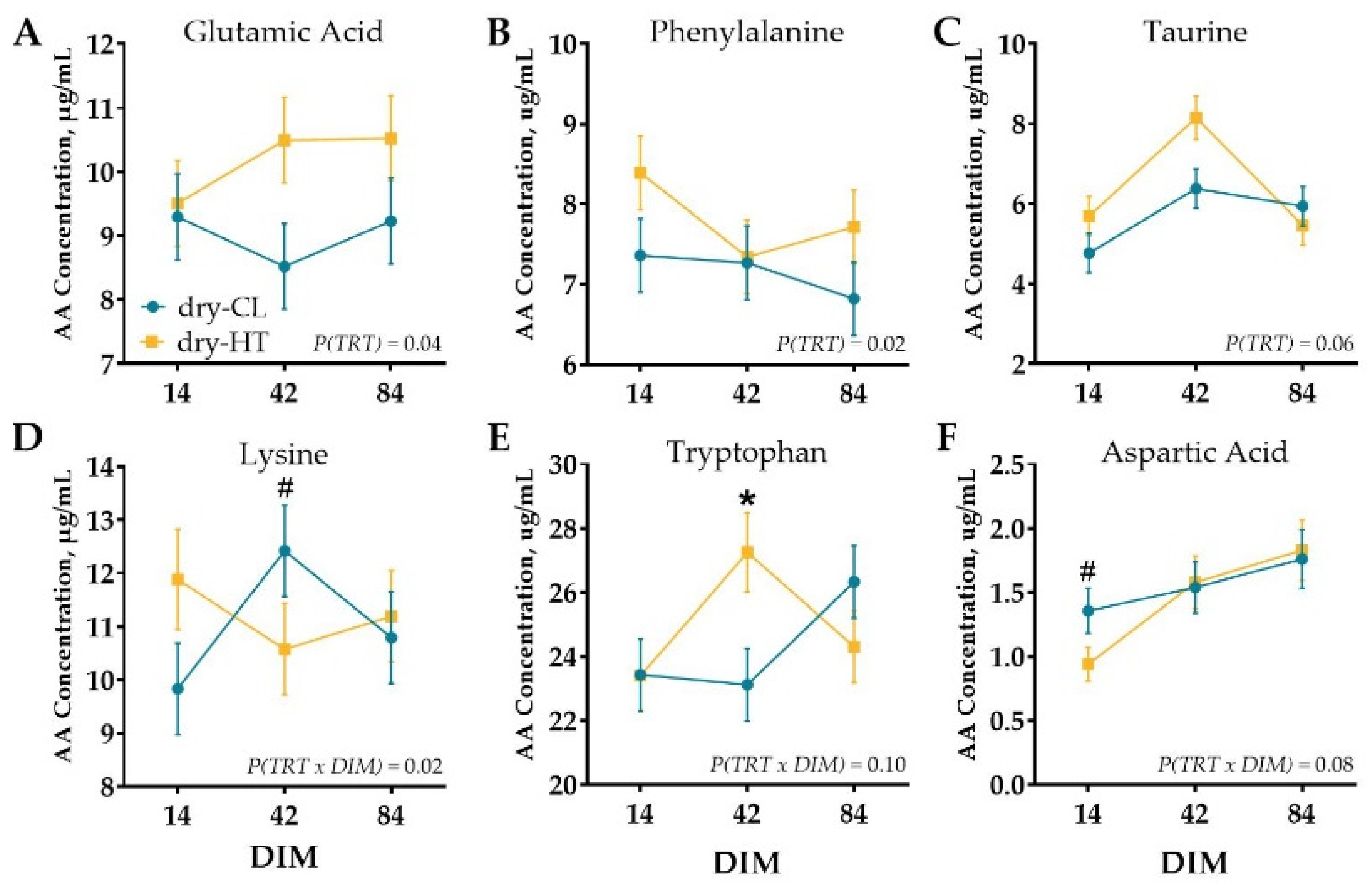
3.2. Milk Protein Profile and Plasma and Tissue AA
3.3. Mammary Tissue AA Transporter Gene Expression
3.4. Mammary Tissue mTOR Gene and Protein Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic Losses from Heat Stress by US Livestock Industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Yousef, M.K. Principle of bioclimatology and adaptation. In Bioclimatology and the Adaptation of Livestock; Johnson, H.D., Ed.; Elsevier Science Publisher: Amsterdam, The Netherlands, 1987; pp. 17–29. [Google Scholar]
- Berman, A. Estimates of Heat Stress Relief Needs for Holstein Dairy Cows. J. Anim. Sci. 2005, 83, 1377–1384. [Google Scholar] [CrossRef]
- Zimbelman, R.B.; Rhoads, R.P.; Rhoads, M.L.; Duff, G.C.; Baumgard, L.H.; Collier, R.J. A Re-Evaluation of the Impact of Temperature Humidity Index (THI) and Black Globe Humidity Index (BGHI) on Milk Production in High Producing Dairy Cows. In Proceedings of the Southwest Nutrition & Management Conference, Reno, NV, USA, 9–11 March 2009; pp. 158–169. [Google Scholar]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and Hormonal Acclimation to Heat Stress in Domesticated Ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, R.J.; Beede, D.K.; Thatcher, W.W.; Israel, L.A.; Wilcox, C.J. Influences of Environment and Its Modification on Dairy Animal Health and Production. J. Dairy Sci. 1982, 65, 2213–2227. [Google Scholar] [CrossRef]
- West, J.W.; Mullinix, B.G.; Bernard, J.K. Effects of Hot, Humid Weather on Milk Temperature, Dry Matter Intake, and Milk Yield of Lactating Dairy Cows. J. Dairy Sci. 2003, 86, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of Heat Stress and Plane of Nutrition on Lactating Holstein Cows: I. Production, Metabolism, and Aspects of Circulating Somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.T.; Guo, J.; Quan, S.Y.; Nan, X.M.; Fernandez, M.V.S.; Baumgard, L.H.; Bu, D.P. The Effects of Heat Stress on Protein Metabolism in Lactating Holstein Cows. J. Dairy Sci. 2017, 100, 5040–5049. [Google Scholar] [CrossRef] [Green Version]
- Ríus, A.G. Invited Review: Adaptations of Protein and Amino Acid Metabolism to Heat Stress in Dairy Cows and Other Livestock Species. Appl. Anim. Sci. 2019, 35, 39–48. [Google Scholar] [CrossRef]
- Cowley, F.C.; Barber, D.G.; Houlihan, A.V.; Poppi, D.P. Immediate and Residual Effect of Heat Stres and Restricted Intake on Milk Protein and Casein Composition and Energy Metabolism. J. Dairy Sci. 2015, 98, 2356–2368. [Google Scholar] [CrossRef] [Green Version]
- Hales, J.R.S. Effects of Exposure to Hot Environments on the Regional Distribution of Blood Flow and on Cardiorespiratory Function in Sheep. Pflügers Arch. 1973, 344, 133–148. [Google Scholar] [CrossRef]
- Lough, D.S.; Beede, D.L.; Wilcox, C.J. Effects of Feed Intake and Thermal Stress on Mammary Blood Flow and Other Physiological Measurements in Lactating Dairy Cows. J. Dairy Sci. 1990, 73, 325–332. [Google Scholar] [CrossRef]
- McGuire, M.A.; Beede, D.K.; DeLorenzo, M.A.; Wilcox, C.J.; Huntington, G.B.; Reynolds, C.K.; Collier, R.J. Effects of Thermal Stress and Level of Feed Intake on Portal Plasma Flow and Net Fluxes of Metabolites in Lactating Holstein Cows. J. Anim. Sci. 1989, 67, 1050–1060. [Google Scholar] [CrossRef] [Green Version]
- Morales, A.; Cota, S.E.M.; Ibarra, N.O.; Arce, N.; Htoo, J.K.; Cervantes, M. Effect of Heat Stress on the Serum Concentrations of Free Amino Acids and Some of Their Metabolites in Growing Pigs1. J. Anim. Sci. 2016, 94, 2835–2842. [Google Scholar] [CrossRef]
- Kaufman, J.D.; Kassube, K.R.; Almeida, R.A.; Ríus, A.G. Short Communication: High Incubation Temperature in Bovine Mammary Epithelial Cells Reduced the Activity of the MTOR Signaling Pathway. J. Dairy Sci. 2018, 101, 7480–7486. [Google Scholar] [CrossRef] [Green Version]
- Collier, R.J.; Stiening, C.M.; Pollard, B.C.; VanBaale, M.J.; Baumgard, L.H.; Gentry, P.C.; Coussens, P.M. Use of Gene Expression Microarrays for Evaluating Environmental Stress Tolerance at the Cellular Level in Cattle1. J. Anim. Sci. 2006, 84, E1–E13. [Google Scholar] [CrossRef] [PubMed]
- Bernabucci, U.; Basiricò, L.; Morera, P.; Dipasquale, D.; Vitali, A.; Cappelli, F.P.; Calamari, L. Effect of Summer Season on Milk Protein Fractions in Holstein Cows. J. Dairy Sci. 2002, 98, 1815–1827. [Google Scholar] [CrossRef] [Green Version]
- Berman, A.; Folman, Y.; Kaim, M.; Mamen, M.; Herz, Z.; Wolfenson, D.; Arieli, A.; Graber, Y. Upper Critical Temperatures and Forced Ventilation Effects for High-Yielding Dairy Cows in a Subtropical Climate. J. Dairy Sci. 1985, 68, 1488–1495. [Google Scholar] [CrossRef]
- Ouellet, V.; Laporta, J.; Dahl, G.E. Late Gestation Heat Stress in Dairy Cows: Effects on Dam and Daughter. Theriogenology 2020, 150, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.C.; Gennari, R.S.; Dahl, G.E.; De Vries, A. Economic Feasibility of Cooling Dry Cows across the United States. J. Dairy Sci. 2016, 99, 9931–9941. [Google Scholar] [CrossRef]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, J.; Bubolz, J.; Dahl, G.E. Heat-Stress Abatement during the Dry Period: Does Cooling Improve Transition into Lactation? J. Dairy Sci. 2009, 92, 5988–5999. [Google Scholar] [CrossRef]
- Tao, S.; Bubolz, J.W.; do Amaral, B.C.; Thompson, I.M.; Hayen, M.J.; Johnson, S.E.; Dahl, G.E. Effect of Heat Stress during the Dry Period on Mammary Gland Development. J. Dairy Sci. 2011, 94, 5976–5986. [Google Scholar] [CrossRef] [PubMed]
- Adin, G.; Gelman, A.; Solomon, R.; Flamenbaum, I.; Nikbachat, M.; Yosef, E.; Zenou, A.; Shamay, A.; Feuermann, Y.; Mabjeesh, S.J.; et al. Effects of Cooling Dry Cows under Heat Load Conditions on Mammary Gland Enzymatic Activity, Intake of Food and Water, and Performance during the Dry Period and after Parturition. Livest. Sci. 2009, 124, 189–195. [Google Scholar] [CrossRef]
- Dado-Senn, B.; Skibiel, A.L.; Fabris, T.F.; Dahl, G.E.; Laporta, J. Dry Period Heat Stress Induces Microstructural Changes in the Lactating Mammary Gland. PLoS ONE 2019, 14, e0222120. [Google Scholar] [CrossRef] [Green Version]
- NRC. A Guide to Environmental Research on Animals; National Academy of Sciences: Washington, DC, USA, 1971. [Google Scholar]
- Dikmen, S.; Hansen, P.J. Is the Temperature-Humidity Index the Best Indicator of Heat Stress in Lactating Dairy Cows in a Subtropical Environment? J. Dairy Sci. 2009, 92, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Kaniyamattam, K.; De Vries, A. Agreement between Milk Fat, Protein, and Lactose Observations Collected from the Dairy Herd Improvement Association (DHIA) and a Real-Time Milk Analyzer. J. Dairy Sci. 2014, 97, 2896–2908. [Google Scholar] [CrossRef]
- Visser, S.; Slangen, C.J.; Rollema, H.S. Phenotyping of Bovine Milk Proteins by Reversed-Phase High-Performance Liquid Chromatography. J. Chromatogr. A 1991, 548, 361–370. [Google Scholar] [CrossRef]
- Le Boucher, J.; Charret, C.; Coudray-Lucas, C.; Giboudeau, J.; Cynober, L. Amino Acid Determination in Biological Fluids by Automated Ion-Exchange Chromatography: Performance of Hitachi L-8500A. Clin. Chem. 1997, 43, 1421–1428. [Google Scholar] [CrossRef] [Green Version]
- Farr, V.C.; Stelwagen, K.; Cate, L.R.; Molenaar, A.J.; McFadden, T.B.; Davis, S.R. An Improved Method for the Routine Biopsy of Bovine Mammary Tissue. J. Dairy Sci. 1996, 79, 543–549. [Google Scholar] [CrossRef]
- Laporta, J.; Keil, K.P.; Vezina, C.M.; Hernandez, L.L. Peripheral Serotonin Regulates Maternal Calcium Trafficking in Mammary Epithelial Cells during Lactation in Mice. PLoS ONE 2014, 9, e110190. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene Networks Driving Bovine Mammary Protein Synthesis during the Lactation Cycle. Bioinform. Biol. Insights 2011, 5, 83–98. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Identification of Reference Genes for Quantitative Real-Time PCR in the Bovine Mammary Gland during the Lactation Cycle. Physiol. Genom. 2007, 29, 312–319. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pszczolkowski, V.L.; Halderson, S.J.; Meyer, E.J.; Lin, A.; Arriola Apelo, S.I. Pharmacologic Inhibition of MTORC1 Mimics Dietary Protein Restriction in a Mouse Model of Lactation. J. Anim. Sci. Biotechnol. 2020, 11, 67. [Google Scholar] [CrossRef]
- Fabris, T.F.; Laporta, J.; Skibiel, A.L.; Corra, F.N.; Senn, B.D.; Wohlgemuth, S.E.; Dahl, G.E. Effect of Heat Stress during Early, Late, and Entire Dry Period on Dairy Cattle. J. Dairy Sci. 2019, 102, 5647–5656. [Google Scholar] [CrossRef]
- Tao, S.; Dahl, G.E.; Laporta, J.; Bernard, J.K.; Orellana Rivas, R.M.; Marins, T.N. Physiology Symposium: Effects of Heat Stress during Late Gestation on the Dam and Its Calf. J. Anim. Sci. 2019, 97, 2245–2257. [Google Scholar] [CrossRef]
- Robertshaw, D. Thermal regulation and the thermal environment. In Dukes’ Physiology of Domestic Animals; Reece, W.O., Ed.; Cornell University Press: Ithaca, NY, USA, 2004; pp. 962–973. [Google Scholar]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat Stress Abatement during the Dry Period Influences Metabolic Gene Expression and Improves Immune Status in the Transition Period of Dairy Cows. J. Dairy Sci. 2011, 94, 86–96. [Google Scholar] [CrossRef]
- Bailey, K.W.; Jones, C.M.; Heinrichs, A.J. Economic Returns to Holstein and Jersey Herds under Multiple Component Pricing. J. Dairy Sci. 2005, 88, 2269–2280. [Google Scholar] [CrossRef]
- Moody, E.G.; Van Soest, P.J.; McDowell, R.E.; Ford, G.L. Effect of High Temperature and Dietary Fat on Milk Fatty Acids. J. Dairy Sci. 1971, 54, 1457–1460. [Google Scholar] [CrossRef]
- Gao, S.T.; Ma, L.; Zhou, Z.; Zhou, Z.K.; Baumgard, L.H.; Jiang, D.; Bionaz, M.; Bu, D.P. Heat Stress Negatively Affects the Transcriptome Related to Overall Metabolism and Milk Protein Synthesis in Mammary Tissue of Midlactating Dairy Cows. Physiol. Genom. 2019, 51, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Mercier, J.C.; Gaye, P. Milk protein synthesis. In Biochemistry of Lactation; Mepham, T.B., Ed.; Elsevier Science Publisher: Amsterdam, The Netherlands, 1983; pp. 177–230. [Google Scholar]
- Tao, S.; Thompson, I.M.; Monteiro, A.P.A.; Hayen, M.J.; Young, L.J.; Dahl, G.E. Effect of Cooling Heat-Stressed Dairy Cows during the Dry Period on Insulin Response. J. Dairy Sci. 2012, 95, 5035–5046. [Google Scholar] [CrossRef] [PubMed]
- Zachut, M.; Kra, G.; Livshitz, L.; Portnick, Y.; Yakoby, S.; Friedlander, G.; Levin, Y. Seasonal Heat Stress Affects Adipose Tissue Proteome toward Enrichment of the Nrf2-Mediated Oxidative Stress Response in Late-Pregnant Dairy Cows. J. Proteom. 2017, 158, 52–61. [Google Scholar] [CrossRef]
- Guo, J.; Gao, S.; Quan, S.; Zhang, Y.; Bu, D.; Wang, J. Blood Amino Acids Profile Responding to Heat Stress in Dairy Cows. AJAS 2018, 31, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Newsholme, P. Why Is L-Glutamine Metabolism Important to Cells of the Immune System in Health, Postinjury, Surgery or Infection? J. Nutr. 2001, 131, 2515S–2522S. [Google Scholar] [CrossRef]
- Morris, A.M.; Treweek, T.M.; Aquilina, J.A.; Carver, J.A.; Walker, M.J. Glutamic Acid Residues in the C-Terminal Extension of Small Heat Shock Protein 25 Are Critical for Structural and Functional Integrity. FEBS J. 2008, 275, 5885–5898. [Google Scholar] [CrossRef]
- Afolayan, A.J.; Alexander, M.; Holme, R.L.; Michalkiewicz, T.; Rana, U.; Teng, R.; Zemanovic, S.; Sahoo, D.; Pritchard, K.A.; Konduri, G.G. Domain Mapping of Heat Shock Protein 70 Reveals That Glutamic Acid 446 and Arginine 447 Are Critical for Regulating Superoxide Dismutase 2 Function * Edited by Linda Spremulli. J. Biol. Chem. 2017, 292, 2369–2378. [Google Scholar] [CrossRef] [Green Version]
- Plakidou-Dymock, S.; McGivan, J.D. Amino Acid Deprivation-Induced Stress Response in the Bovine Renal Epithelial Cell Line NBL-1: Induction of HSP 70 by Phenylalanine. Biochim. Biophys. Acta-Mol. Cell Res. 1994, 1224, 189–197. [Google Scholar] [CrossRef]
- Yang, B.; Wang, X.; Ren, X. Amino Acid Metabolism Related to Immune Tolerance by MDSCs. Int. Rev. Immunol. 2012, 31, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Belal, S.A.; Kang, D.R.; Cho, E.S.R.; Park, G.H.; Shim, K.S. Expression of Heat Shock Proteins in Broilers Exposed to Chronic Heat. Braz. J. Poult. Sci. 2018, 20, 479–486. [Google Scholar] [CrossRef]
- Lu, Z.; He, X.F.; Ma, B.B.; Zhang, L.; Li, J.L.; Jiang, Y.; Zhou, G.H.; Gao, F. The Alleviative Effects and Related Mechanisms of Taurine Supplementation on Growth Performance and Carcass Characteristics in Broilers Exposed to Chronic Heat Stress. Poult. Sci. 2019, 98, 878–886. [Google Scholar] [CrossRef]
- Dado-Senn, B.; Skibiel, A.L.; Fabris, T.F.; Zhang, Y.; Dahl, G.E.; Peñagaricano, F.; Laporta, J. RNA-Seq Reveals Novel Genes and Pathways Involved in Bovine Mammary Involution during the Dry Period and under Environmental Heat Stress. Sci. Rep. 2018, 8, 11096. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.G.; Satter, L.D.; Clay, A.B. Response of Lactating Dairy Cows to Abomasal Infusion of Amino Acids. J. Dairy Sci. 1976, 59, 1254–1270. [Google Scholar] [CrossRef]
- Lin, X.; Li, S.; Zou, Y.; Zhao, F.-Q.; Liu, J.; Liu, H. Lysine Stimulates Protein Synthesis by Promoting the Expression of ATB0,+ and Activating the MTOR Pathway in Bovine Mammary Epithelial Cells. J. Nutr. 2018, 148, 1426–1433. [Google Scholar] [CrossRef]
- Vyas, D.; Erdman, R.A. Meta-Analysis of Milk Protein Yield Responses to Lysine and Methionine Supplementation. J. Dairy Sci. 2009, 92, 5011–5018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassube, K.R.; Kaufman, J.D.; Pohler, K.G.; McFadden, J.W.; Ríus, A.G. Jugular-Infused Methionine, Lysine and Branched-Chain Amino Acids Does Not Improve Milk Production in Holstein Cows Experiencing Heat Stress. Animal 2017, 11, 2220–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, M.J.; Glynn, E.L.; Fry, C.S.; Timmerman, K.L.; Volpi, E.; Rasmussen, B.B.; Mj, D.; El, G.; Cs, F.; Kl, T.; et al. An Increase in Essential Amino Acid Availability Upregulates Amino Acid Transporter Expression in Human Skeletal Muscle. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1011–E1018. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Summers, C.M.; Pearce, S.C.; Gabler, N.K.; Valentine, R.J.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Short-Term Heat Stress Altered Metabolism and Insulin Signaling in Skeletal Muscle. J. Anim. Sci. 2018, 96, 154–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.; Reyna-Neyra, A.; Philippe, L.; Thoreen, C.C. MTORC1 Balances Cellular Amino Acid Supply with Demand for Protein Synthesis through Post-Transcriptional Control of ATF4. Cell Rep. 2017, 19, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Salama, A.A.K.; Duque, M.; Wang, L.; Shahzad, K.; Olivera, M.; Loor, J.J. Enhanced Supply of Methionine or Arginine Alters Mechanistic Target of Rapamycin Signaling Proteins, Messenger RNA, and MicroRNA Abundance in Heat-Stressed Bovine Mammary Epithelial Cells in Vitro. J. Dairy Sci. 2019, 102, 2469–2480. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.-S. The Role of Mammalian Target of Rapamycin (MTOR) in Insulin Signaling. Nutrients 2017, 9, 1176. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein (%) | Treatment | p-Value | ||||
|---|---|---|---|---|---|---|
| Dry-CL | Dry-HT | SEM | TRT | DIM | TRT × DIM | |
| Low Molecular Weight | 13.42 | 13.38 | 1.06 | 0.95 | <0.01 | 0.52 |
| Peptides | 0.51 | 0.77 | 0.12 | 0.19 | 0.02 | 0.96 |
| α-Lactalbumin | 3.84 | 3.73 | 0.23 | 0.84 | 0.77 | 0.43 |
| β-Lactoglobulin | 8.04 | 8.96 | 0.59 | 0.74 | 0.25 | 0.59 |
| α-S1 Casein | 31.76 | 32.14 | 0.61 | 0.48 | 0.08 | 0.56 |
| α-S2 Casein | 7.28 | 7.64 | 0.45 | 0.81 | 0.18 | 0.88 |
| β-Casein | 31.30 | 29.69 | 0.85 | 0.20 | 0.13 | 0.74 |
| γ-Casein | 0.57 | 0.68 | 0.10 | 0.30 | 0.13 | 0.49 |
| κ-Casein | 3.38 | 3.17 | 0.19 | 0.37 | 0.73 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dado-Senn, B.; Skibiel, A.L.; Dahl, G.E.; Arriola Apelo, S.I.; Laporta, J. Dry Period Heat Stress Impacts Mammary Protein Metabolism in the Subsequent Lactation. Animals 2021, 11, 2676. https://doi.org/10.3390/ani11092676
Dado-Senn B, Skibiel AL, Dahl GE, Arriola Apelo SI, Laporta J. Dry Period Heat Stress Impacts Mammary Protein Metabolism in the Subsequent Lactation. Animals. 2021; 11(9):2676. https://doi.org/10.3390/ani11092676
Chicago/Turabian StyleDado-Senn, Bethany, Amy L. Skibiel, Geoffrey E. Dahl, Sebastian I. Arriola Apelo, and Jimena Laporta. 2021. "Dry Period Heat Stress Impacts Mammary Protein Metabolism in the Subsequent Lactation" Animals 11, no. 9: 2676. https://doi.org/10.3390/ani11092676
APA StyleDado-Senn, B., Skibiel, A. L., Dahl, G. E., Arriola Apelo, S. I., & Laporta, J. (2021). Dry Period Heat Stress Impacts Mammary Protein Metabolism in the Subsequent Lactation. Animals, 11(9), 2676. https://doi.org/10.3390/ani11092676