miR-27a Regulates Sheep Adipocyte Differentiation by Targeting CPT1B Gene

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ovine Preadipocytes Isolation and Culture
2.2. Cloning of CPT1B
2.3. Cell Transfection
2.4. Determination of Gene Expression
2.5. Quantitative Assessment of Adipocyte Differentiation Using Lipid (Oil Red O) Staining Assay
2.6. Luciferase Reporter Assay
2.7. Western Blot Assay
2.8. Data Analysis
3. Results
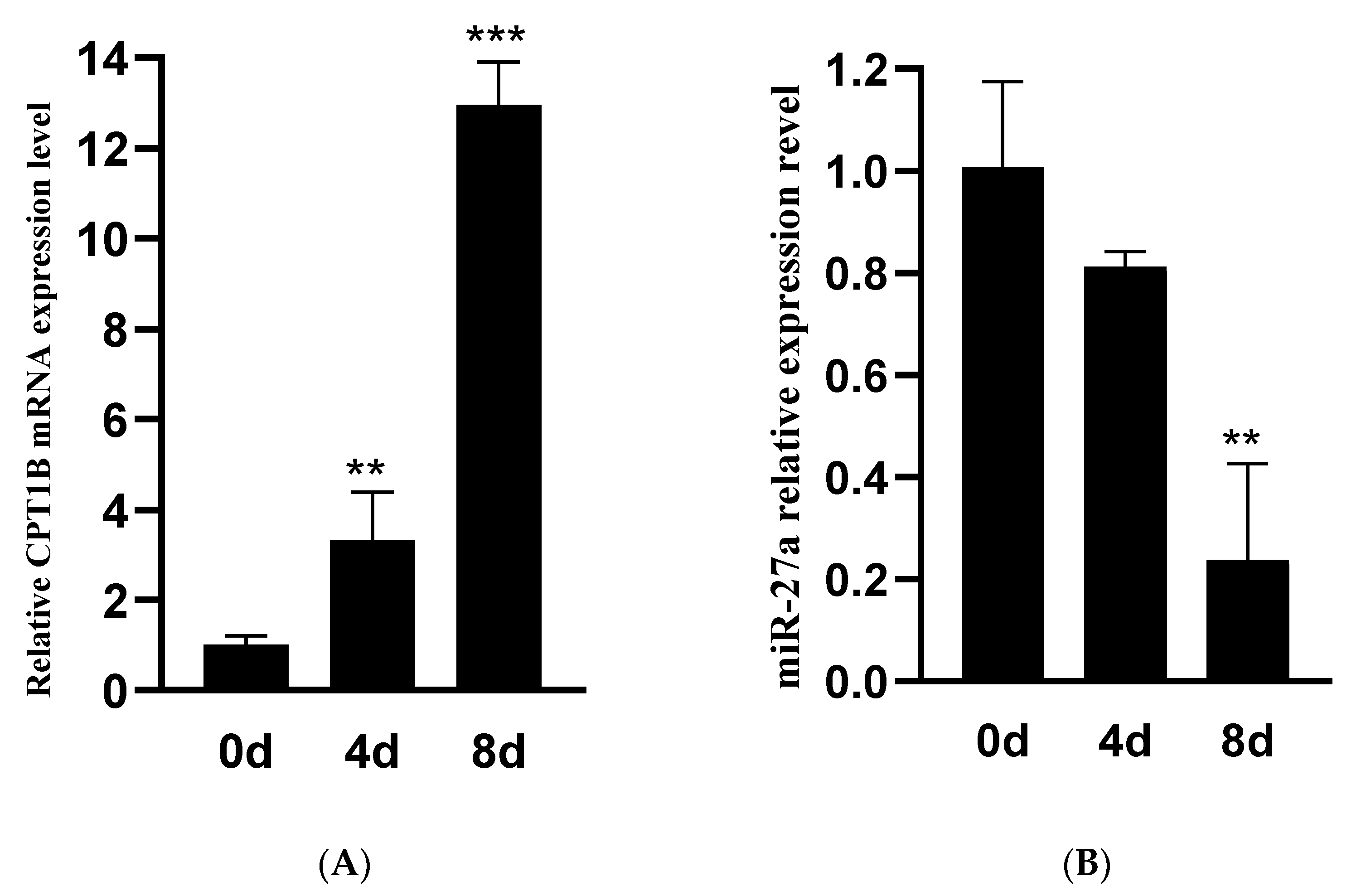
3.1. Expression of miR-27a and CPT1B under Ovine Preadipocytes Differentiation
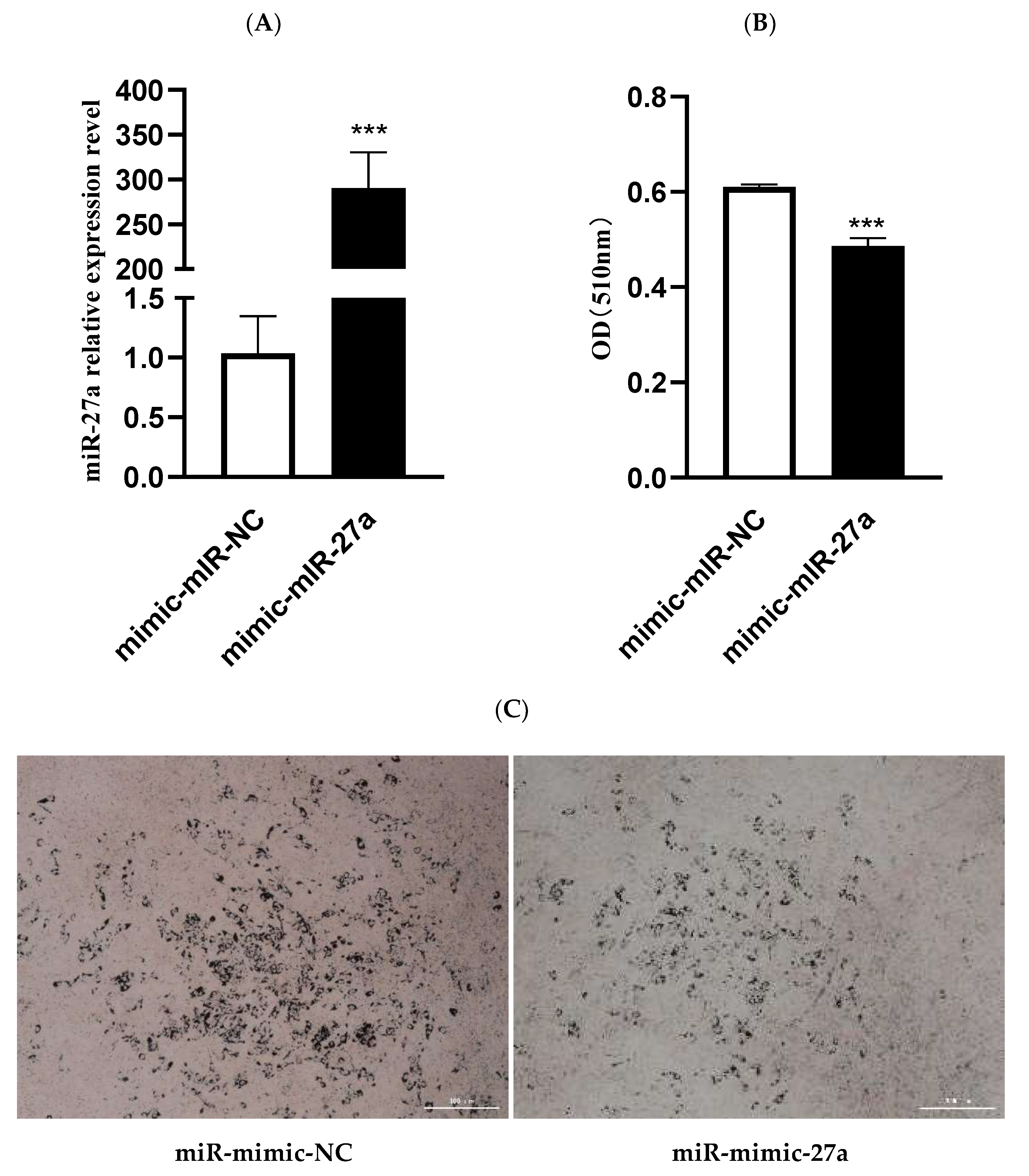
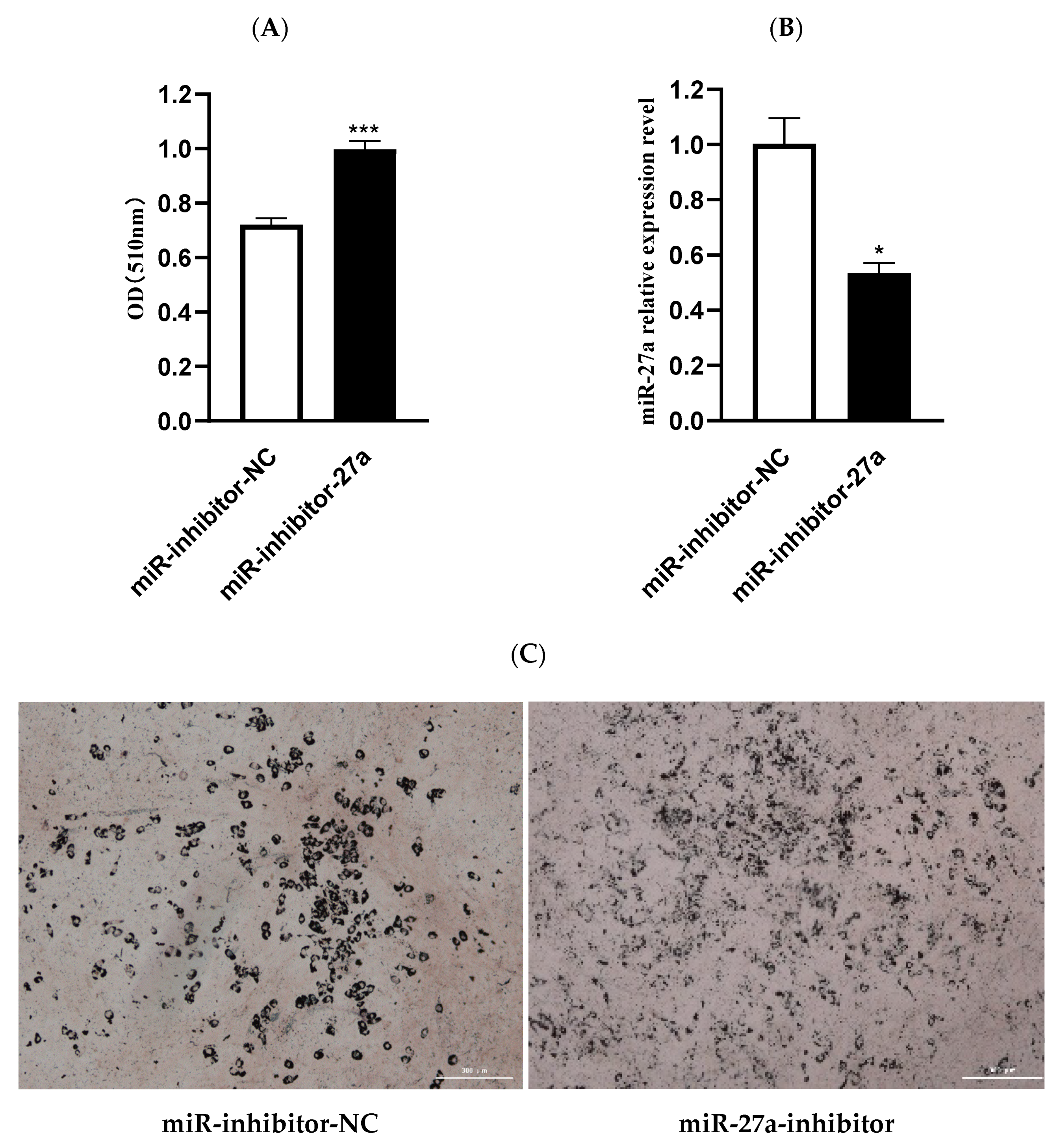
3.2. MiR-27a Inhibits Differentiation of Ovine Preadipocytes
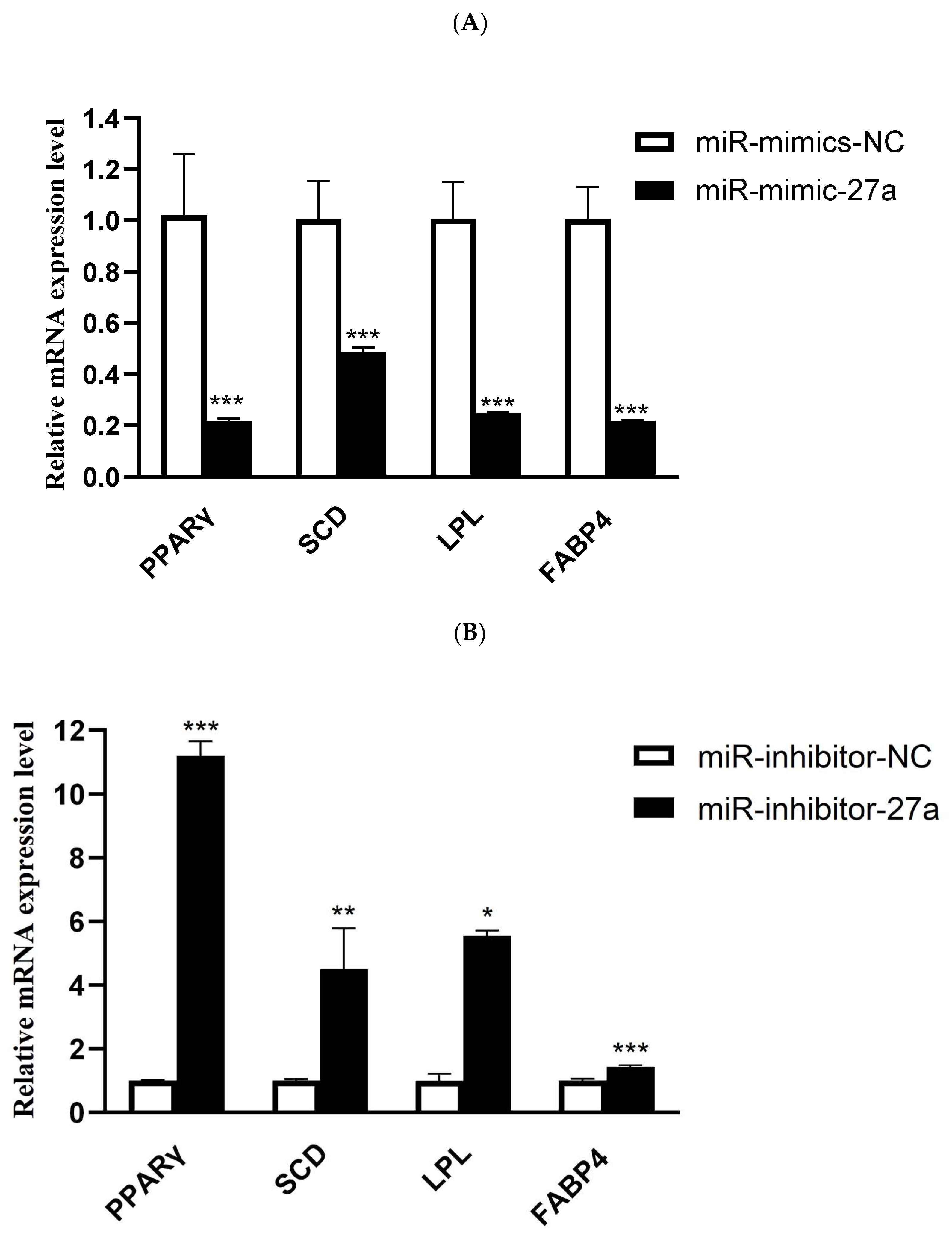
3.3. MiR-27a Regulates Expression of Genes Related to Lipid Metabolism in Ovine Preadipocytes
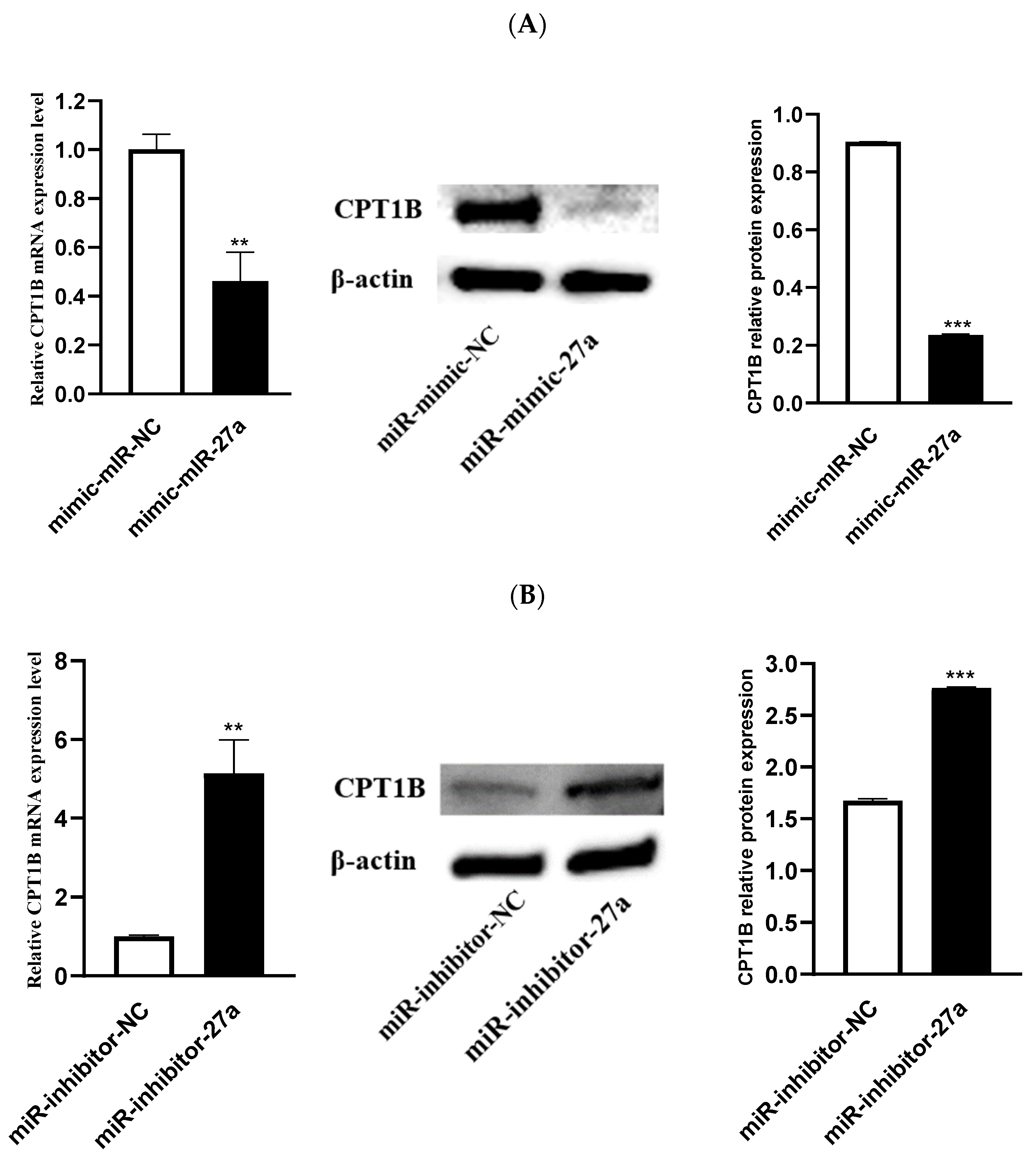
3.4. CPT1B Is a Target Gene for miR-27a
3.5. Effect of CPT1B Overexpression on Ovine Preadipocytes Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.; Watkins, P.; Ball, A.; Krishnamurthy, R.; Piyasiri, U.; Sewell, J.; Ortuño, J.; Stark, J.; Warner, R. Impact of Brassica and lucerne finishing feeds and intramuscular fat on lamb eating quality and flavor. A cross-cultural study using Chinese and Non-Chinese Australian consumers. J. Agric. Food Chem. 2016, 64, 6856–6868. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Qiao, L.; Han, Y.; Liu, J.; Zhang, J.; Liu, W. Regulatory Roles of SREBF1 and SREBF2 in lipid metabolism and deposition in two Chinese representative fat-tailed sheep breeds. Animals 2020, 10, 1317. [Google Scholar] [CrossRef] [PubMed]
- Watkins, P.J.; Rose, G.; Salvatore, L.; Allen, D.; Tucman, D.; Warner, R.D.; Dunshea, F.R.; Pethick, D.W. Age and nutrition influence the concentrations of three branched chain fatty acids in sheep fat from Australian abattoirs. Meat Sci. 2010, 86, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Zhang, Y.; Zhang, J.; Yao, Q.; Song, Y.; Shen, Q.; Lin, J.; Gao, Y.; Wang, X.; Zhang, L.; et al. Cold-inducible Klf9 regulates thermogenesis of brown and beige fat. Diabetes 2020, 69, 2603–2618. [Google Scholar] [CrossRef]
- Khanal, P.; Pandey, D.; Binti Ahmad, S.; Safayi, S.; Kadarmideen, H.N.; Olaf Nielsen, M. Differential impacts of late gestational over-and undernutrition on adipose tissue traits and associated visceral obesity risk upon exposure to a postnatal high-fat diet in adolescent sheep. Physiol. Rep. 2020, 8, e14359. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Mohr, A.M.; Mott, J.L. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, B.; Qiao, L.; Liu, J.; Ren, D.; Liu, W. miR-340–5p inhibits sheep adipocyte differentiation by targeting ATF7. Anim. Sci. J. 2020, 91, e13462. [Google Scholar] [CrossRef] [PubMed]
- Dusaulcy, R.; Handgraaf, S.; Visentin, F.; Vesin, C.; Philippe, J.; Gosmain, Y. miR-132–3p is a positive regulator of alpha-cell mass and is downregulated in obese hyperglycemic mice. Mol. Metab. 2019, 22, 84–95. [Google Scholar] [CrossRef]
- Pan, Y.; Jing, J.; Qiao, L.; Liu, J.; An, L.; Li, B.; Ren, D.; Liu, W. MiRNA-seq reveals that miR-124–3p inhibits adipogenic differentiation of the stromal vascular fraction in sheep via targeting C/EBPα. Domest. Anim. Endocrinol. 2018, 65, 17–23. [Google Scholar] [CrossRef]
- Shi, T.; Yan, X.; Qiao, L.; Li, B.; Cheng, L.; Pan, Y.; Jing, J.; Cao, N.; Liu, W. MiR-330–5p negatively regulates ovine preadipocyte differentiation by targeting branched-chain aminotransferase 2. Anim. Sci. J. 2018, 89, 858–867. [Google Scholar] [CrossRef]
- Deng, K.; Ren, C.; Fan, Y.; Liu, Z.; Zhang, G.; Zhang, Y.; You, P.; Wang, F. miR-27a is an important adipogenesis regulator associated with differential lipid accumulation between intramuscular and subcutaneous adipose tissues of sheep. Domest. Anim. Endocrinol 2020, 71, 106393. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, A.Y.; Lee, H.W.; Son, Y.H.; Lee, G.Y.; Lee, J.-W.; Lee, Y.S.; Kim, J.B. miR-27a is a negative regulator of adipocyte differentiation via suppressing PPARgamma expression. Biochem. Biophys. Res. Commun. 2010, 392, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, I.R.; Joshi, M. CPT1A-mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef]
- Dang, Y.; Xu, J.; Zhu, M.; Zhou, W.; Zhang, L.; Ji, G. Gan-Jiang-Ling-Zhu decoction alleviates hepatic steatosis in rats by the miR-138–5p/CPT1B axis. Biomed. Pharmacother. 2020, 127, 110127. [Google Scholar] [CrossRef] [PubMed]
- Deng, K.; Ren, C.; Liu, Z.; Gao, X.; Fan, Y.; Zhang, G.; Zhang, Y.; Ma, E.-S.; Wang, F.; You, P. Characterization of RUNX1T1, an Adipogenesis Regulator in Ovine Preadipocyte Differentiation. Int J. Mol. Sci. 2018, 19, 1300. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Du, J.; Wang, L.; Niu, L.; Zhao, Y.; Tang, G.; Jiang, Y.; Shuai, S.; Bai, L.; Li, X.; et al. MicroRNA-143a-3p modulates preadipocyte proliferation and differentiation by targeting MAPK7. Biomed. Pharmacother. 2018, 108, 531–539. [Google Scholar] [CrossRef]
- Cash, J.L.; Hart, R.; Russ, A.; Dixon, J.P.; Colledge, W.H.; Doran, J.; Hendrick, A.G.; Carlton, M.B.; Greaves, D.R. Synthetic chemerin-derived peptides suppress inflammation through ChemR23. J. Exp. Med. 2008, 205, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, Y.; Guo, J.; Zhong, T.; Li, L.; Zhang, H.; Wang, L. Identification and expression patterns of adipokine genes during adipocyte differentiation in the Tibetan goat (Capra hircus). Gene 2018, 643, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Narita, T.; Kobayashi, M.; Itakura, K.; Itagawa, R.; Kabaya, R.; Sudo, Y.; Okita, N.; Higami, Y. Differential response to caloric restriction of retroperitoneal, epididymal, and subcutaneous adipose tissue depots in rats. Exp. Gerontol. 2018, 104, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Richmond, S.R.; Carper, M.J.; Lei, X.; Zhang, S.; Yarasheski, K.E.; Ramanadham, S. HIV-protease inhibitors suppress skeletal muscle fatty acid oxidation by reducing CD36 and CPT1 fatty acid transporters. Biochim. Biophys. Acta 2010, 1801, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, B.V.; Yao, F.; Howenstine, A.; Takenaka, R.; Hyatt, J.A.; Sears, K.E.; Shewchuk, B.M. Emergent coordination of the CHKB and CPT1B genes in eutherian mammals: Implications for the origin of brown adipose tissue. J. Mol. Biol. 2020, 432, 6127–6145. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Gao, M.; Yang, R.; Zhao, Z.; Mi, J.; Sun, H.; Xiao, H.; Fang, X. The effect of CPT1B gene on lipid metabolism and its polymorphism analysis in Chinese Simmental cattle. Anim. Biotechnol. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vantaku, V.; Dong, J.; Ambati, C.R.; Perera, D.; Donepudi, S.R.; Amara, C.S.; Putluri, V.; Ravi, S.S.; Robertson, M.J.; Piyarathna, D.W.B.; et al. Multi-omics integration analysis robustly predicts high-grade patient survival and identifies CPT1B effect on fatty acid metabolism in bladder cancer. Clin. Cancer Res. 2019, 25, 3689–3701. [Google Scholar] [CrossRef] [Green Version]
- Van Weeghel, M.; Abdurrachim, D.; Nederlof, R.; Argmann, C.A.; Houtkooper, R.H.; Hagen, J.; Nabben, M.; Denis, S.; Ciapaite, J.; Kolwicz, S.C.; et al. Increased cardiac fatty acid oxidation in a mouse model with decreased malonyl-CoA sensitivity of CPT1B. Cardiovasc. Res. 2018, 114, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Rosqvist, F.; Parry, S.A. The influence of dietary fatty acids on liver fat content and metabolism. Proc. Nutr. Soc. 2020, 79, 30–41. [Google Scholar] [CrossRef]
- Hess, B.W.; Moss, G.E.; Rule, D.C. A decade of developments in the area of fat supplementation research with beef cattle and sheep. J. Anim. Sci. 2008, 86, E188–E204. [Google Scholar] [CrossRef]
- Chakrabarty, K.; Romans, J.R. Lipogenesis in the adipose cells of the bovine (Bos taurus) as related to their intramuscular fat content. Comp. Biochem. Physiol. B 1972, 41, 603–615. [Google Scholar] [CrossRef]
- Justin, L.; Raymond, A.; John, M.; Masatoshi, M.; Lawrence, R.; Dale, R.; Jacqueline, W. Maternal and fetal growth, body composition, endocrinology, and metabolic status in undernourished adolescent sheep. Narnia 2007, 77, 343–350. [Google Scholar]
- Sun, Y.; Wang, Q.; Zhang, Y.; Geng, M.; Wei, Y.; Liu, Y.; Liu, S.; Petersen, R.B.; Yue, J.; Huang, K.; et al. Multigenerational maternal obesity increases the incidence of HCC in offspring via miR-27a-3p. J. Hepatol. 2020, 73, 603–615. [Google Scholar] [CrossRef]
- He, C.; Zhang, Q.; Sun, H.; Cai, R.; Pang, W. Role of miRNA and lncRNA in animal fat deposition-a review. Chin. J. Biotechnol. 2020, 36, 1504–1514. [Google Scholar]
- Engin, A.B. MicroRNA and adipogenesis. Adv. Exp. Med. Biol. 2017, 960, 489–509. [Google Scholar] [PubMed]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Chen, Z.-Q.; Zhu, J.-J. Silencing of microRNA-27a facilitates autophagy and apoptosis of melanoma cells through the activation of the SYK-dependent mTOR signaling pathway. J. Cell Biochem. 2020, 121, 4694–4695. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Y.; Cao, X.; Mou, P. MiR-27a facilitates breast cancer progression via GSK-3β. Technol. Cancer Res. Treat. 2020, 19, 1533033820965576. [Google Scholar] [CrossRef]
- Choe, N.; Kwon, D.-H.; Ryu, J.; Shin, S.; Cho, H.J.; Joung, H.; Eom, G.H.; Ahn, Y.; Park, W.J.; Nam, K.-I.; et al. Targets ATF3 to reduce calcium deposition in vascular smooth muscle cells. Mol. Ther. Nucleic Acids 2020, 22, 627–639. [Google Scholar] [CrossRef]
- Lin, X.-Z.; Luo, J.; Zhang, L.-P.; Wang, W.; Shi, H.-B.; Zhu, J.-J. MiR-27a suppresses triglyceride accumulation and affects gene mRNA expression associated with fat metabolism in dairy goat mammary gland epithelial cells. Gene 2013, 521, 15–23. [Google Scholar] [CrossRef]
- Shi, C.; Huang, F.; Gu, X.; Zhang, M.; Wen, J.; Wang, X.; You, L.; Cui, X.; Ji, C.; Guo, X. Adipogenic miRNA and meta-signature miRNAs involved in human adipocyte differentiation and obesity. Oncotarget 2016, 7, 40830–40845. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Tang, K.; Wang, Y.; Zan, L. MiR-27a-5p increases steer fat deposition partly by targeting calcium-sensing receptor (CASR). Sci. Rep. 2018, 8, 3012. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Gao, Z.; Alarcon, R.M.; Ye, J.; Yun, Z. A role of miR-27 in the regulation of adipogenesis. FEBS J. 2009, 276, 2348–2358. [Google Scholar] [CrossRef]
- Teimouri, M.; Hosseini, H.; Shabani, M.; Koushki, M.; Noorbakhsh, F.; Meshkani, R. Inhibiting miR-27a and miR-142–5p attenuate nonalcoholic fatty liver disease by regulating Nrf2 signaling pathway. IUBMB Life 2020, 72, 361–372. [Google Scholar] [CrossRef]
- Yu, Y.; Du, H.; Wei, S.; Feng, L.; Li, J.; Yao, F.; Zhang, M.; Hatch, G.M.; Chen, L. Adipocyte-derived exosomal MiR-27a induces insulin resistance in skeletal muscle through repression of PPARγ. Theranostics 2018, 8, 2171–2188. [Google Scholar] [CrossRef]
- Zou, B.; Ge, Z.; Zhu, W.; Xu, Z.; Li, C. Persimmon tannin represses 3T3-L1 preadipocyte differentiation via up-regulating expression of miR-27 and down-regulating expression of peroxisome proliferator-activated receptor-γ in the early phase of adipogenesis. Eur. J. Nutr. 2015, 54, 1333–1343. [Google Scholar] [CrossRef]
- Deng, K.; Ren, C.; Fan, Y.; Pang, J.; Zhang, G.; Zhang, Y.; You, P.; Wang, F. YAP1 regulates PPARG and RXR alpha expression to affect the proliferation and differentiation of ovine preadipocyte. J. Cell Biochem. 2019, 120, 19578–19589. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Guo, T.; Yue, Y.; Lu, Z.; Liu, J.; Yuan, C.; Niu, C.; Yang, M.; Yang, B. Quantitative proteomic analysis identified differentially expressed proteins with tail/rump fat deposition in Chinese thin- and fat-tailed lambs. PLoS ONE 2021, 16, e0246279. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, M.; Gutiérrez-Gil, B.; García-Gámez, E.; Sánchez, J.P.; Arranz, J.J. Detection of quantitative trait loci affecting the milk fatty acid profile on sheep chromosome 22: Role of the stearoyl-CoA desaturase gene in Spanish Churra sheep. J. Dairy Sci. 2010, 93, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.-S.; Hu, S.-L.; Yu, K.; Wang, H.; Wang, W.; Loor, J.; Luo, J. Lipoprotein lipase, tissue expression and effects on genes related to fatty acid synthesis in goat mammary epithelial cells. Int. J. Mol. Sci. 2014, 15, 22757–22771. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Target | Primer Sequence (5′-3′) | Gene ID |
|---|---|---|
| ꞵ-actin(internal reference of mRNA) | F: ACCGTGAGAAGATGACCCAGA | 443052 |
| R: AGAGGCGTACAGGGACAGCA | ||
| PPAR γ | F: TGGATGACCACTCCCATGCC | 443513 |
| R: TTGGGAACGGAATGTCCTC | ||
| FABP4 | F: GGATGATAAGCTGGTGCTGG | 100137067 |
| R: CTCTGGTAGCAGTGACACCG | ||
| SCD | F: TTCATCCTGCCCACACTCG | 443185 |
| R: TAGTTGTGGAAGCCCTCACC | ||
| LPL | F: CCCAGCAGCATTATCCAGTGT | 443408 |
| R: ATTCATCCGCCATCCAGTTC | ||
| miR-27a | F: TCGGCAGGTTCACAGTGGCTA | 102465824 |
| R: CTCAACTGGTGTCGTGGAGTC | ||
| U6 (internal reference of mRNA) | F: CAAGGGCCACATAGATCCG | 101121962 |
| R: AACGCTTCACGAATTTGCGT | ||
| CPT1B | F: AGATCCGTATGTTCGACCCAA | 443193 |
| R: CTGCGATCATGTAGGAAACACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Huang, X.; Yang, C.; Ge, T.; Zhao, L.; Zhang, X.; Tian, L.; Zhang, E. miR-27a Regulates Sheep Adipocyte Differentiation by Targeting CPT1B Gene. Animals 2022, 12, 28. https://doi.org/10.3390/ani12010028
Li B, Huang X, Yang C, Ge T, Zhao L, Zhang X, Tian L, Zhang E. miR-27a Regulates Sheep Adipocyte Differentiation by Targeting CPT1B Gene. Animals. 2022; 12(1):28. https://doi.org/10.3390/ani12010028
Chicago/Turabian StyleLi, Bo, Xiaoyu Huang, Chen Yang, Ting Ge, Leiyun Zhao, Xiaoqiang Zhang, Lintao Tian, and Enping Zhang. 2022. "miR-27a Regulates Sheep Adipocyte Differentiation by Targeting CPT1B Gene" Animals 12, no. 1: 28. https://doi.org/10.3390/ani12010028