
Individual Feed Efficiency Monitoring of Charolaise Candidate Young Bulls in Relation to Feeding Behavior and Self-Performance Test Results

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arthur, R.K. Genetic and phenotypic relationships among different measures of growth and feed efficiency in young Charolais bulls. Livest. Prod. Sci. 2001, 68, 131–139. [Google Scholar] [CrossRef]
- Nkrumah, J.D.; Basarab, J.A.; Wang, Z.; Li, C.; Price, M.A.; Okine, E.K.; Moore, S.S. Genetic and phenotypic relationships of feed intake and measures of efficiency with growth and carcass merit of beef cattle. J. Anim. Sci. 2007, 85, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Grion, A.L.; Mercadante, M.E.Z.; Cyrillo, J.N.S.G.; Bonilha, S.F.M.; Magnani, E.; Branco, R.H. Selection for feed efficiency traits and correlated genetic responses in feed intake and weight gain of Nellore cattle. J. Anim. Sci. 2014, 92, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Mehrban, H.; Naserkheil, M.; Lee, D.; Ibáñez-Escriche, N. Genetic parameters and correlations of related feed efficiency, growth, and carcass traits in Hanwoo beef cattle. Anim. Biosci. 2021, 34, 824–832. [Google Scholar] [CrossRef]
- Herd, R.M.; Oddy, V.H.; Richardson, E.C. Biological basis for variation in residual feed intake in beef cattle. 1. Review of potential mechanisms. Austral. J. Experiment. Agric. 2004, 44, 423–430. [Google Scholar] [CrossRef]
- Lancaster, P.A.; Carstens, G.E.; Crews, D.H.; Welsh, T.H.; Forbes, T.D.A.; Forrest, D.W.; Rouquette, F.M. Phenotypic and genetic relationships of residual feed intake with performance and ultrasound carcass traits in Brangus heifers. J. Anim. Sci. 2009, 87, 3887–3896. [Google Scholar] [CrossRef] [PubMed]
- Koch, R.M.; Swiger, L.A.; Chambers, D.; Gregory, K.E. Efficiency of feed use in beef cattle. J. Anim. Sci. 1963, 22, 486–494. [Google Scholar] [CrossRef]
- Elolimy, A.A.; Abdelmegeid, M.K.; McCann, J.C.; Shike, D.W.; Loor, J.J. Residual feed intake in beef cattle and its association with carcass traits, ruminal solid-fraction bacteria, and epithelium gene expression. J. Anim. Sci. Biotechnol. 2018, 9, 67. [Google Scholar] [CrossRef]
- Basarab, J.A.; Colazo, M.G.; Ambrose, D.J.; Novak, S.; McCartney, D.; Baron, V.S. Residual feed intake adjusted for backfat thickness and feeding frequency is independent of fertility in beef heifers. Can. J. Anim. Sci. 2011, 91, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Mao, F.; Chen, L.; Vinsky, M.; Okine, E.; Wang, Z.; Basarab, J.; Crews, D.; Li, C. Phenotypic and genetic relationships of feed efficiency with growth performance, ultrasound, and carcass merit traits in angus and charolais steers. J. Anim. Sci. 2013, 91, 2067–2076. [Google Scholar] [CrossRef]
- Kenny, D.A.; Fitzsimons, C.; Waters, S.M.; McGee, M. Invited review: Improving feed efficiency of beef cattle: Current state of the art and future challenges. Animal 2018, 12, 1815–1826. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.; Ducrocq, V.; Faverdin, P.; Friggens, N. Invited review: Disentangling residual feed intake-Insights and approaches to make it more fit for purpose in the modern context. J. Dairy Sci. 2021, 104, 6329–6342. [Google Scholar] [CrossRef] [PubMed]
- Esfandyari, H.; Jensen, J. Simultaneous Bayesian estimation of genetic parameters for curves of weight, feed intake, and residual feed intake in beef cattle. J. Anim. Sci. 2021, 99, skab231. [Google Scholar] [CrossRef]
- Manca, E.; Cesarani, A.; Falchi, L.; Atzori, A.S.; Gaspa, G.; Rossoni, A.; Macciotta, N.P.P.; Dimauro, C. Genome-wide association study for residual concentrate intake using different approaches in Italian Brown Swiss, Italian. J. Anim. Sci. 2021, 20, 957–1967. [Google Scholar] [CrossRef]
- Schwartzkopf-Genswein, K.; Huisma, C.; McAllister, T.A. Validation of a radio frequency identification system for monitoring the feeding patterns of feedlot cattle. Livest. Prod. Sci. 1999, 60, 27–31. [Google Scholar] [CrossRef]
- Krawczel, P.; Klaiber, L.; Thibeau, S.; Dann, H. Technical note: Data loggers are a valid method for assessing the feeding behavior of dairy cows using the Calan Broadbent Feeding System. J. Dairy Sci. 2012, 95, 4452–4456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancaster, P.A.; Carstens, G.E.; Ribeiro, F.R.; Tedeschi, L.O.; Crews, D.H. Characterization of feed efficiency traits and relationships with feeding behaviour and ultrasound carcass traits in growing bulls. J. Anim. Sci. 2009, 87, 1528–1539. [Google Scholar] [CrossRef]
- Mendes, E.D.M.; Carstens, G.E.; Tedeschi, L.O.; Pinchak, W.E.; Friend, T.H. Validation of a system for monitoring feeding behavior in beef cattle. J. Anim. Sci. 2011, 89, 2904–2910. [Google Scholar] [CrossRef] [PubMed]
- Puzio, N.; Purwin, C.; Nogalski, Z.; Białobrzewski, I.; Tomczyk, L.; Michalski, J.P. The effects of age and gender (bull vs steer) on the feeding behavior of young beef cattle fed grass silage. AsianAustralas. J. Anim. Sci. 2019, 32, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, D.N.; Sleator, R.D.; Murphy, C.P.; Conroy, S.B.; Judge, M.M.; Berry, D.P. Large variability in feeding behavior among crossbred growing cattle. J. Anim. Sci. 2020, 98, skaa216. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.L. On the interpretation of feeding behaviour measures and the use of feeding rate as an indicator of social constraint. Appl. Anim. Behav. Sci. 1999, 63, 79–91. [Google Scholar] [CrossRef]
- Capper, J.L. The environmental impact of beef production in the United States: 1977 compared with 2007. J. Anim. Sci. 2011, 89, 4249–4261. [Google Scholar] [CrossRef] [Green Version]
- Meale, S.J.; Morgavi, D.P.; Cassar-Malek, I.; Andueza, D.; Ortigues-Marty, I.; Robins, R.J.; Schiphorst, A.-M.; Laverroux Graulet, S.B.; Boudra, H.; Cantalapiedra-Hijar, G. Exploration of biological markers of feed efficiency in young bulls. J. Agric. Food Chem. 2017, 65, 9817–9827. [Google Scholar] [CrossRef]
- Imaz, J.A.; García, S.; González, L.A. Real-time monitoring of self-fed supplement intake, feeding behaviour, and growth rate as affected by forage quantity and quality of rotationally grazed beef cattle. Animals 2019, 9, 1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzanilla-Pech, C.I.V.; Veerkamp, R.F.; Tempelman, R.J.; van Pelt, M.L.; Weigel, K.A.; VandeHaar, M.; Lawlor, T.J.; Spurlock, D.M.; Armentano, L.E.; Staples, C.R.; et al. Genetic parameters between feed-intake-related traits and conformation in 2 separate dairy populations: The Netherlands and United States. J. Dairy Sci. 2016, 99, 443–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Török, M. In Vivo Ultrasonic Measurements in Beef Cattle Breeding towards Developing Breeding Value Estimation. Ph.D. Thesis, Pannon University Georgikon Faculty, Keszthely, Hungary, 2009. [Google Scholar]
- Bullock, D. Improving herd efficiency. Charolais J. 2000, 34, 70–71. [Google Scholar]
- Fitzsimons, C.; Kenny, D.A.; McGee, M. Visceral organ weights, digestion and carcass characteristics of beef bulls differing in residual feed intake offered a high concentrate diet. Animal 2014, 8, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Trevizan, N.; Canesin, R.C.; Branco, R.H.; Batalha, C.D.A.; Cyrillo, J.N.S.G.; Bonilha, S.F.M. Growth, ruminal and metabolic parameters and feeding behavior of Nellore cattle with different residual feed intake phenotypes. Livest. Sci. 2021, 244, 1043931. [Google Scholar] [CrossRef]
- Hyslop, J.J.; Fuller, R.; Taylor, U.; Thirlwell, D.; Wareing, S. Feed intake, animal performance and net feed efficiency (NFE) in finishing Stabiliser steers. In Proceedings of the British Society of Animal Science (BSAS), Nottingham, UK, 29–30 April 2014; p. 127. [Google Scholar]
- Consolo, N.R.B.; Buarque, V.L.M.; Silva, J.; Poleti, M.D.; Barbosa, L.C.G.S.; Higuera-Padilla, A.; Gomez, J.F.M.; Colnago, L.A.; Gerrard, D.E.; Saran Netto, A.; et al. Muscle and liver metabolomic signatures associated with residual feed intake in Nellore cattle. Anim. Feed Sci. Technol. 2021, 271, 114757. [Google Scholar] [CrossRef]
- Lancaster, P.A.; Carstens, G.E.; Michal, J.J.; Brennan, K.M.; Johnson, K.A.; Davis, M.E. Relationships between residual feed intake and hepatic mitochondrial function in growing beef cattle. J. Anim. Sci. 2014, 92, 3134–3141. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.; Oltjen, J.W.; Sainz, R.D. Mitochondrial abundance and function in muscle from beef steers with divergent residual feed intakes. Animal 2020, 14, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Jiu, Z.; Roy, B.C.; Das, C.W.; Wendy, V.J.; Juarez, M.; Fitzsimmons, C.; Li, C.; Plastow, G.; Aalhus, J.L.; Bruce, H.L. Meat and sensory quality of major muscles from Angus, Charolais, and Angus crossbred steers with high and low residual feed intake. Can. J. Anim. Sci. 2020, 100, 140–153. [Google Scholar] [CrossRef]
- McKenna, C.; Porter, R.K.; Fitzsimons, C.; Waters, S.M.; McGee, M.; Kenny, D.A. Mitochondrial abundance and function in skeletal muscle and liver from Simmental beef cattle divergent for residual feed intake. Animal 2020, 14, 1710–1717. [Google Scholar] [CrossRef]
- Basarab, J.A.; Price, M.A.; Aalhus, J.L.; Okine, E.K.; Snelling, W.M.; Lyle, K.L. Residual feed intake and body composition in young growing cattle. Can. J. Anim. Sci. 2003, 83, 189–204. [Google Scholar] [CrossRef]
- Baker, S.D.; Szasz, J.I.; Klein, T.A.; Kuber, P.S.; Hunt, C.W.; Glaze, J.B.; Hill, R.A. Residual feed intake of purebred Angus steers: Effects on meat quality and palatability. J. Anim. Sci. 2006, 84, 938–945. [Google Scholar] [CrossRef] [PubMed]
- McDonagh, M.B.; Herd, R.M.; Richardson, E.C.; Oddy, V.H.; Archer, J.A.; Arthur, P.F. Meat quality and the calpain system of feedlot steers following a single generation of divergent selection for residual feed intake. Aust. J. Expt. Agric. 2001, 41, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Richardson, E.C.; Herd, R.M.; Oddy, V.H.; Thompson, J.M.; Archer, J.A.; Arthur, P.F. Body composition and implications for heat production of Angus steer progeny of parents selected for and against residual feed intake. Aust. J. Exp. Agric. 2001, 41, 1065–1072. [Google Scholar] [CrossRef] [Green Version]
- Perkins, S.D.; Key, C.N.; Garrett, C.F.; Foradori, C.D.; Bratcher, C.L.; Kriese-Anderson, L.A.; Brandebourg, T.D. Residual feed intake studies in Angus-sired cattle reveal a potential role for hypothalamic gene expression in regulating feed efficiency. J. Anim. Sci. 2014, 92, 549–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Colazo, M.G.; Basarab, J.A.; Goonewardene, L.A.; Ambrose, D.J.; Marques, E.; Plastow, G.; Miller, S.P.; Moore, S.S. Impact of selection for residual feed intake on breeding soundness and reproductive performance of bulls on pasture-based multisire mating. J. Anim. Sci. 2012, 90, 2963–2969. [Google Scholar] [CrossRef] [Green Version]
- Awda, B.J.; Miller, S.P.; Montanholi, Y.R.; Voort, G.V.; Caldwell, T.; Buhr, M.M.; Swanson, K.C. The relationship between feed efficiency traits and fertility in young beef bulls. Can. J. Anim. Sci. 2013, 93, 185–192. [Google Scholar] [CrossRef]
- Haskell, M.J.; Rooke, J.A.; Roehe, R.; Turner, S.P.; Hyslop, J.J.; Waterhouse, A.; Duthie, C.-A. Relationships between feeding behaviour, activity, dominance and feed efficiency in finishing beef steers. Appl. Anim. Behav. Sci. 2019, 210, 9–15. [Google Scholar] [CrossRef]
- Romanzin, A.; Degano, L.; Vicario, D.; Spanghero, M. Feeding efficiency and behavior of young Simmental bulls selected for high growth capacity: Comparison of bulls with high vs. low residual feed intake. Livest. Sci. 2021, 249, 104525. [Google Scholar] [CrossRef]
- Montanholi, Y.R.; Swanson, K.C.; Palme, R.; Schenkel, F.S.; McBride, B.W.; Lu, D.; Miller, S.P. Assessing feed efficiency in beef steers through feeding behavior, infrared thermography and glucocorticoids. Animal 2010, 4, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra-Hijar, G.; Abo-Ismail, M.; Carstens, G.E.; Guan, L.L.; Hegarty, R.; Kenny, D.A.; McGee, M.; Plastow, G.; Relling, A.; Ortigues-Marty, I. Review: Biological determinants of between-animal variation in feed efficiency of growing beef cattle. Animal 2018, 12, s321–s335. [Google Scholar] [CrossRef] [Green Version]
- Pereira, I.C.; Costa, C.F.; Martins, C.L.; Pereira, M.C.S.; Squizatti, M.M.; Owens, F.N.; Cruz, G.D.; Millen, D.D.; Arrigoni, M.D.B. Voluntary daily fluctuation in dry matter intake is associated to feedlot performance, feeding behavior and rumen morphometrics in beef cattle. Livest. Sci. 2021, 250, 104565. [Google Scholar] [CrossRef]
- Schneider, L.; Volkmann, N.; Kemper, N.; Spindler, B. Feeding behavior of fattening bulls fed six times per day using an automatic feeding system. Front. Vet. Sci. 2020, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Philipps, C. Cattle Behaviour and Welfare, 2nd ed.; Blackwell Science Ltd.: Oxford, UK, 2002. [Google Scholar]
- Kelly, A.M.; McGee, D.; Crews, D.H.; Sweeney, T.; Boland, T.M.; Kenny, D. Repeatability of feed efficiency, carcass ultrasound, feeding behavior, and blood metabolic variables in finishing heifers divergently selected for residual feed intake. J. Anim. Sci. 2010, 88, 3214–3225. [Google Scholar] [CrossRef]
- DelCurto-Wyffels, H.M.; Dafoe, J.M.; Parsons, C.T.; Boss, D.L.; DelCurto, T.; Wyffels, S.A.; Van Emon, M.L.; Bowman, J.G.P. Corn versus barley in finishing diets: Effect on steer performance and feeding behavior. Animals 2021, 11, 935. [Google Scholar] [CrossRef]
- Castro Bulle, F.C.P.; Paulino, P.V.; Sanches, A.C.; Sainz, R.D. Growth, carcass quality, and protein and energy metabolism in beef cattle with different growth potentials and residual feed intakes. J. Anim. Sci. 2007, 85, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Hümmel, J. Physiological adaptations of ruminants and their potential relevance for production systems. R. Bras. Zootec. 2017, 46, 606–613. [Google Scholar] [CrossRef] [Green Version]


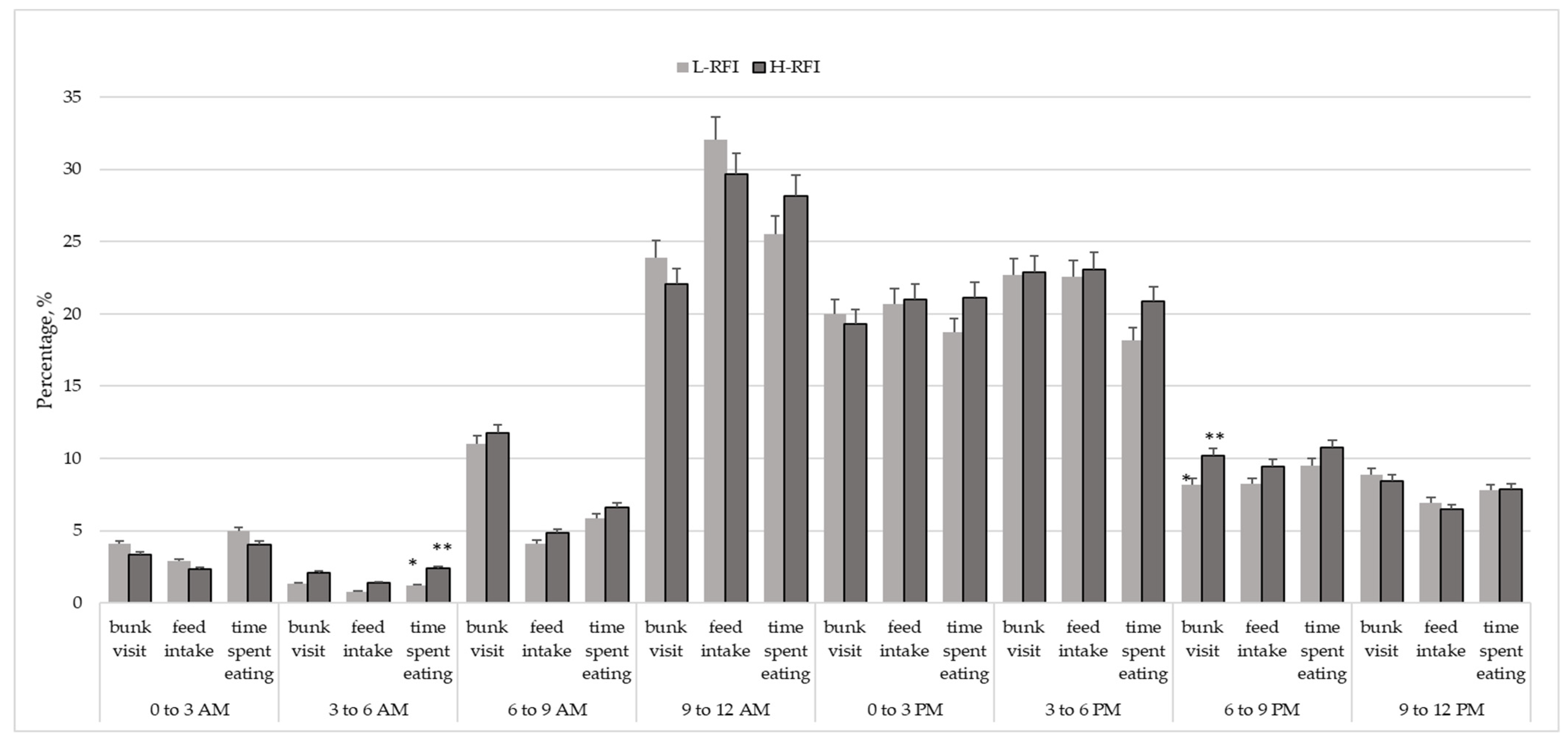{kind=link}
| Ingredient [%] | I. Period | II. Period |
|---|---|---|
| Vitalbull®concentrate feedstuff | 32 | 35 |
| alfalfa hay | 26 | 25 |
| alfalfa haylage | 21 | 20 |
| triticale haylage | 21 | 20 |
| Chemical composition | ||
| Dry matter [%] | 69.52 | 70.52 |
| Crude ash | 9.22 | 9.16 |
| Energy dependent metabolizable protein | 9.55 | 9.64 |
| Crude fat | 3.03 | 3.13 |
| Crude fiber | 19.98 | 19.06 |
| Starch | 18.82 | 20.57 |
| Acid detergent fiber | 23.57 | 22.56 |
| Neutral detergent fiber | 33.37 | 33.30 |
| NEm [MJ/kg] | 6.07 | 6.17 |
| NEg [MJ/kg] | 3.43 | 3.54 |
| Minerals, vitamins | ||
| Ca [g] | 0.92 | 0.92 |
| P [g] | 0.35 | 0.37 |
| Na [g] | 0.19 | 0.21 |
| Vitamin A [IU] | 9642.32 | 10,686.90 |
| Vitamin D [IU] | 1902.95 | 2109.10 |
| Vitamin E [mg] | 48.00 | 53.20 |
| Traits | L-RFI | H-RFI | SEM | p-Value (RFI) |
|---|---|---|---|---|
| Residual feed intake (kg/day) | −0.64 | 1.13 | 0.244 | 0.000 |
| Dry matter intake (kg/day) | 13.24 | 15.02 | 0.331 | 0.004 |
| Body weight, kg | ||||
| 0 day | 453.67 | 464.90 | 10.62 | 0.612 |
| 29 day | 513.22 | 522.18 | 11.13 | 0.700 |
| 57 day | 566.67 | 575.09 | 11.44 | 0.725 |
| Final body weight | 615.11 | 622.36 | 11.53 | 0.764 |
| Average daily gain (kg/day) | 1.95 | 1.90 | 0.06 | 0.693 |
| Total gain (kg) | 161.44 | 157.46 | 4.84 | 0.693 |
| Live weight gain | 1.53 | 1.50 | 0.024 | 0.808 |
| Gain to feed | 0.14 | 0.12 | 0.004 | 0.005 |
| Ultrasonography measurements | ||||
| Longissimus muscle area (cm2) | 86.83 | 93.61 | 2.33 | 0.152 |
| Back fat (mm) | 4.9 | 4.4 | 0.32 | 0.974 |
| Rump fat (mm) | 6.0 | 5.5 | 0.28 | 0.316 |
| Intramuscular fat level (%) | 5.13 | 5.06 | 0.26 | 0.265 |
| Traits | L-RFI | H-RFI | SEM | p-Value (RFI) |
|---|---|---|---|---|
| Muscularity (score) | ||||
| shoulder width | 6.11 | 6.55 | 0.233 | 0.367 |
| back width | 5.89 | 6.36 | 0.233 | 0.323 |
| roundness of thigh | 5.33 | 6.18 | 0.268 | 0.117 |
| width of thigh | 5.44 | 6.36 | 0.312 | 0.147 |
| loin thickness | 5.89 | 6.36 | 0.221 | 0.297 |
| Skeletal measurements (score) | ||||
| canon girth | 4.56 | 4.91 | 0.123 | 0.158 |
| back-loin length | 6.55 | 6.09 | 0.391 | 0.569 |
| rump length | 5.89 | 5.82 | 0.357 | 0.925 |
| width of hip bones | 5.22 | 6.09 | 0.272 | 0.115 |
| frame | 5.89 | 5.27 | 0.366 | 0.417 |
| Functional traits (score) | ||||
| muzzle width | 5.56 | 5.82 | 0.300 | 0.675 |
| forelegs | 4 | 5.45 | 0.329 | 0.023 |
| hind legs | 4.33 | 5.5 | 0.284 | 0.09 |
| top line straightness | 6.11 | 6.27 | 0.213 | 0.716 |
| chest depth | 5.33 | 5.82 | 0.197 | 0.231 |
| chest width | 5.22 | 5.73 | 0.199 | 0.215 |
| rump width | 5.22 | 6.09 | 0.272 | 0.115 |
| thigh length | 5.0 | 5.36 | 0.258 | 0.497 |
| Body condition score | 6.33 | 6.7 | 0.193 | 0.881 |
| Traits | L-RFI | H-RFI | SEM | p-Value (RFI) |
|---|---|---|---|---|
| Time spent eating, per day, min | 136.83 | 129.17 | 6.04 | 0.543 |
| Visits per day | 32.30 | 54.19 | 4.82 | 0.019 |
| Time/visit min | 4.46 | 2.83 | 0.353 | 0.018 |
| Eating rate g/min | 146.36 | 174.59 | 10.20 | 0.175 |
| Intake g/body weight/day | 21.60 | 24.17 | 0.418 | 0.000 |
| Intake g/visit | 324.24 | 236.97 | 23.06 | 0.060 |
| SD of intake per day/kg DM | 2.24 | 2.78 | 0.150 | 0.072 |
| CV of intake per day/kg DM | 17.12 | 18.41 | 0.907 | 0.494 |
| RFI | G:F | ADG | LMA | BFT | P8 | IMF | |
|---|---|---|---|---|---|---|---|
| G:F | −0.61 ** | 1 | 0.67 ** | −0.42 | - | 0.29 | −0.42 |
| Shoulder width | - | −0.33 | - | 0.61 ** | - | - | - |
| Back width | - | −0.27 | - | 0.53 * | - | −0.26 | - |
| Roundness of thigh | 0.29 | −0.42 | - | 0.79 ** | - | −0.26 | - |
| Width of thigh | 0.21 | −0.39 | - | 0.66 ** | −0.2 | - | - |
| Loin thickness | - | −0.21 | - | 0.57 ** | −0.2 | - | - |
| Canon girth | 0.28 | - | 0.24 | 0.33 | −0.2 | - | - |
| Back loin length | −0.45 * | - | - | 0.31 | −0.32 | −0.33 | - |
| Rump length | −0.39 | - | - | 0.61 ** | −0.36 | ||
| Width of hip bones | 0.26 | −0.37 | - | 0.61 ** | - | - | - |
| Frame | −0.47 * | 0.29 | 0.21 | 0.25 | −0.36 | −0.29 | - |
| Muzzle width | −0.50 * | −0.26 | - | 0.43 | −0.22 | - | - |
| Forelegs | 0.56 ** | 0.23 | - | 0.30 | −0.33 | −0.23 | - |
| Hind legs | 0.34 | - | 0.32 | - | −0.35 | - | - |
| Top line straightness | - | - | - | - | −0.66 ** | −0.44 * | −0.46 * |
| Chest depth | 0.29 | −0.44 * | - | 0.52 * | - | - | - |
| Chest width | 0.30 | −0.43 | - | 0.63 ** | - | - | 0.21 |
| Rump width | 0.26 | −0.37 | - | 0.61 ** | - | - | - |
| Thigh length | - | −0.36 | −0.33 | 0.50 * | - | - | - |
| BCS | 0.20 | - | 0.26 | 0.29 | 0.56 * | 0.30 | 0.27 |
| Time spent eating, per day, min | - | 0.21 | 0.46 * | −0.34 | - | 0.20 | 0.35 |
| Visits per day | 0.63 ** | - | - | - | - | - | - |
| Time/visit min | −0.56 ** | - | −0.22 | - | - | - | - |
| Eating rate g/min | - | −0.23 | - | 0.44 * | - | - | 0.44 * |
| Intake per day/kg DM | 0.75 ** | −0.31 | 0.46 * | 0.57 ** | - | - | 0.38 |
| Intake g/body weight/day | 0.93 ** | −0.32 | - | - | - | 0.23 | - |
| Intake g/visit | −0.51 * | −0.22 | 0.26 | - | - | 0.22 | |
| SD of intake per day/kg DM | 0.41 | - | 0.35 | - | - | - | - |
| Predictor Trait | Factor | Adjusted R2 | p-Value | SEE | Equations |
|---|---|---|---|---|---|
| Intake g/BW/day(A) | 1 | 0.86 | 0.000 | 0.416 | −12.156 + 0.543A |
| Intake g/BW/day; G:F(B) | 2 | 0.96 | 0.000 | 0.220 | −7.812 + 0.476A − 21.437B |
| Intake g/BW/day; G:F; intake per day(C); | 3 | 0.98 | 0.000 | 0.139 | −8.601 + 0.408A − 19.491B + 0.001C |
| Intake g/BW/day; G:F; intake per day; P8(D); | 4 | 0.99 | 0.000 | 0.064 | −9.005 + 0.433A − 16.370B + 0.001C − 1.036D |
| Intake g/BW/day; G:F; intake per day; P8; chest depth(F) | 5 | 0.99 | 0.000 | 0.052 | −9.409 + 0.442A − 15.327B + 0.001C − 1.053D + 0.049F |
| Intake g/BW/day; G:F; intake per day; P8; chest depth; thigh length(G) | 6 | 0.99 | 0.000 | 0.037 | −9.042 + 0.435A − 16.152B + 0.001C − 0.990D + 0.087F − 0.044G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holló, G.; Nagy-Kiszlinger, H.; Tossenberger, J.; Török, M.; Húth, B. Individual Feed Efficiency Monitoring of Charolaise Candidate Young Bulls in Relation to Feeding Behavior and Self-Performance Test Results. Animals 2022, 12, 35. https://doi.org/10.3390/ani12010035
Holló G, Nagy-Kiszlinger H, Tossenberger J, Török M, Húth B. Individual Feed Efficiency Monitoring of Charolaise Candidate Young Bulls in Relation to Feeding Behavior and Self-Performance Test Results. Animals. 2022; 12(1):35. https://doi.org/10.3390/ani12010035
Chicago/Turabian StyleHolló, Gabriella, Henrietta Nagy-Kiszlinger, János Tossenberger, Márton Török, and Balázs Húth. 2022. "Individual Feed Efficiency Monitoring of Charolaise Candidate Young Bulls in Relation to Feeding Behavior and Self-Performance Test Results" Animals 12, no. 1: 35. https://doi.org/10.3390/ani12010035
APA StyleHolló, G., Nagy-Kiszlinger, H., Tossenberger, J., Török, M., & Húth, B. (2022). Individual Feed Efficiency Monitoring of Charolaise Candidate Young Bulls in Relation to Feeding Behavior and Self-Performance Test Results. Animals, 12(1), 35. https://doi.org/10.3390/ani12010035