
Gastrointestinal Parasite Community and Phenotypic Plasticity in Native and Introduced Alien Lagomorpha

 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
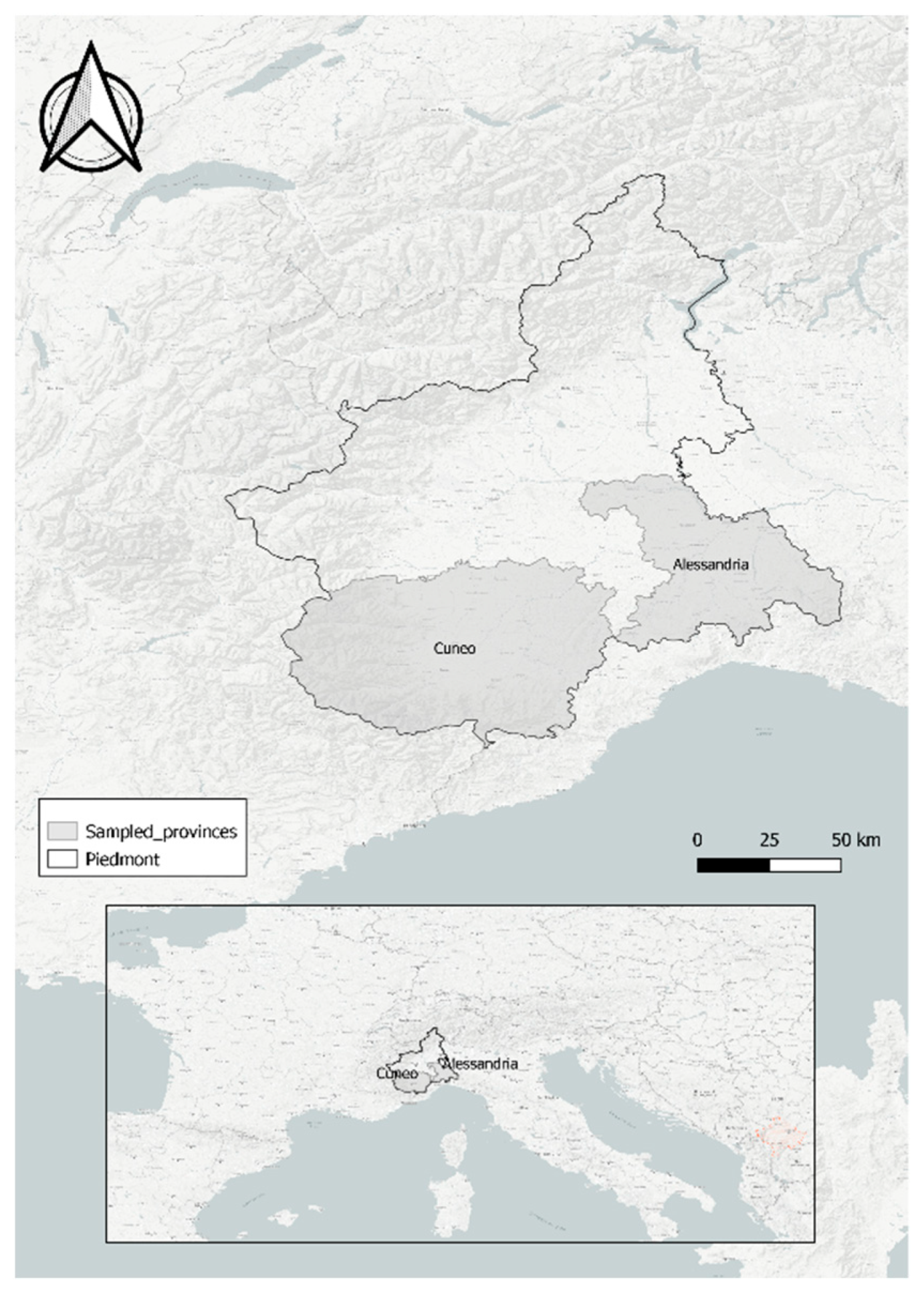
2.1. Study Area
2.2. Collection of Specimens and Parasitological Analysis
2.3. Biometric Measurements of Parasites
2.4. Software for Image Analysis
2.5. Statistical Analysis
3. Results
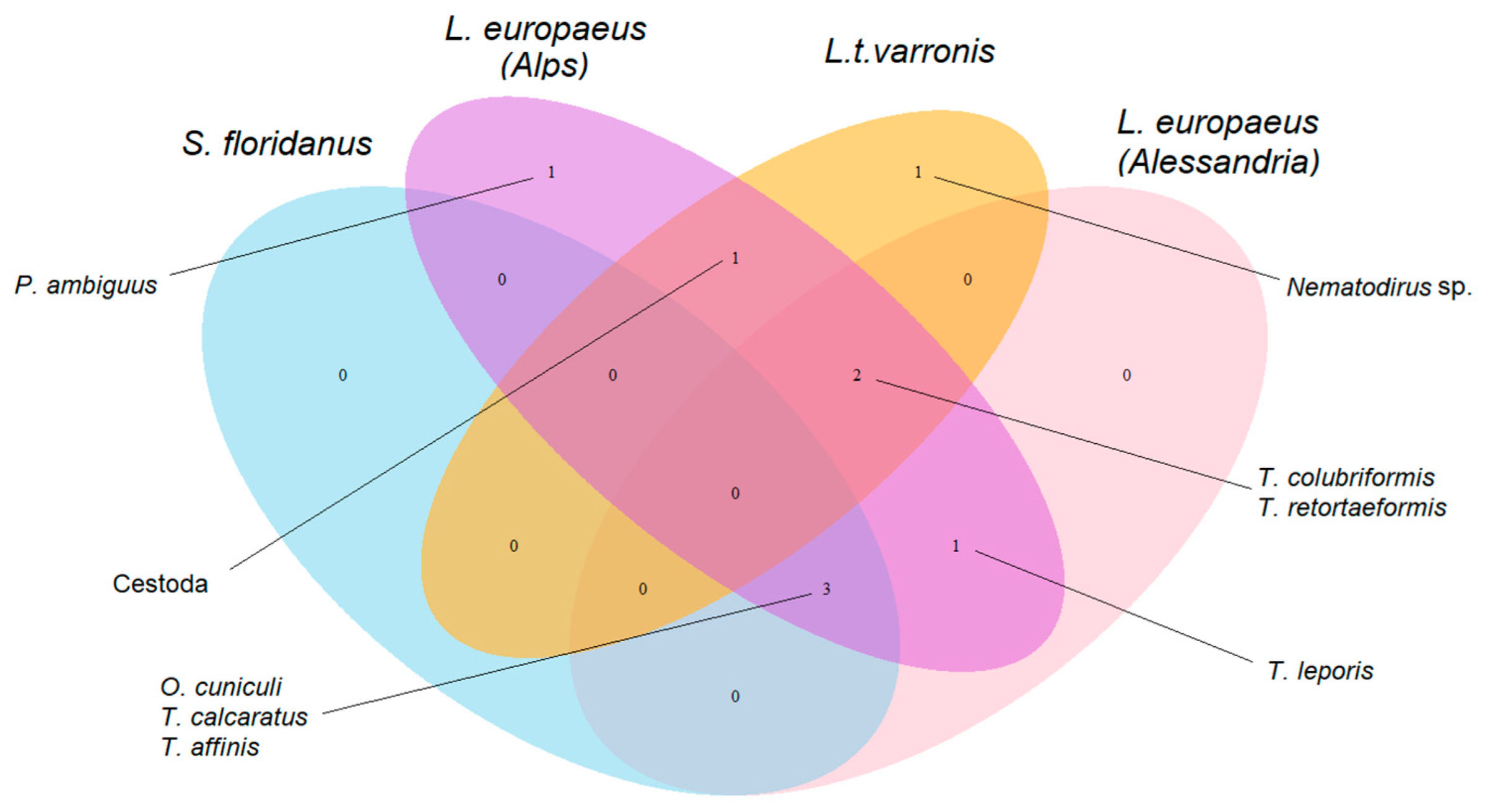
3.1. Description of the Parasite Community
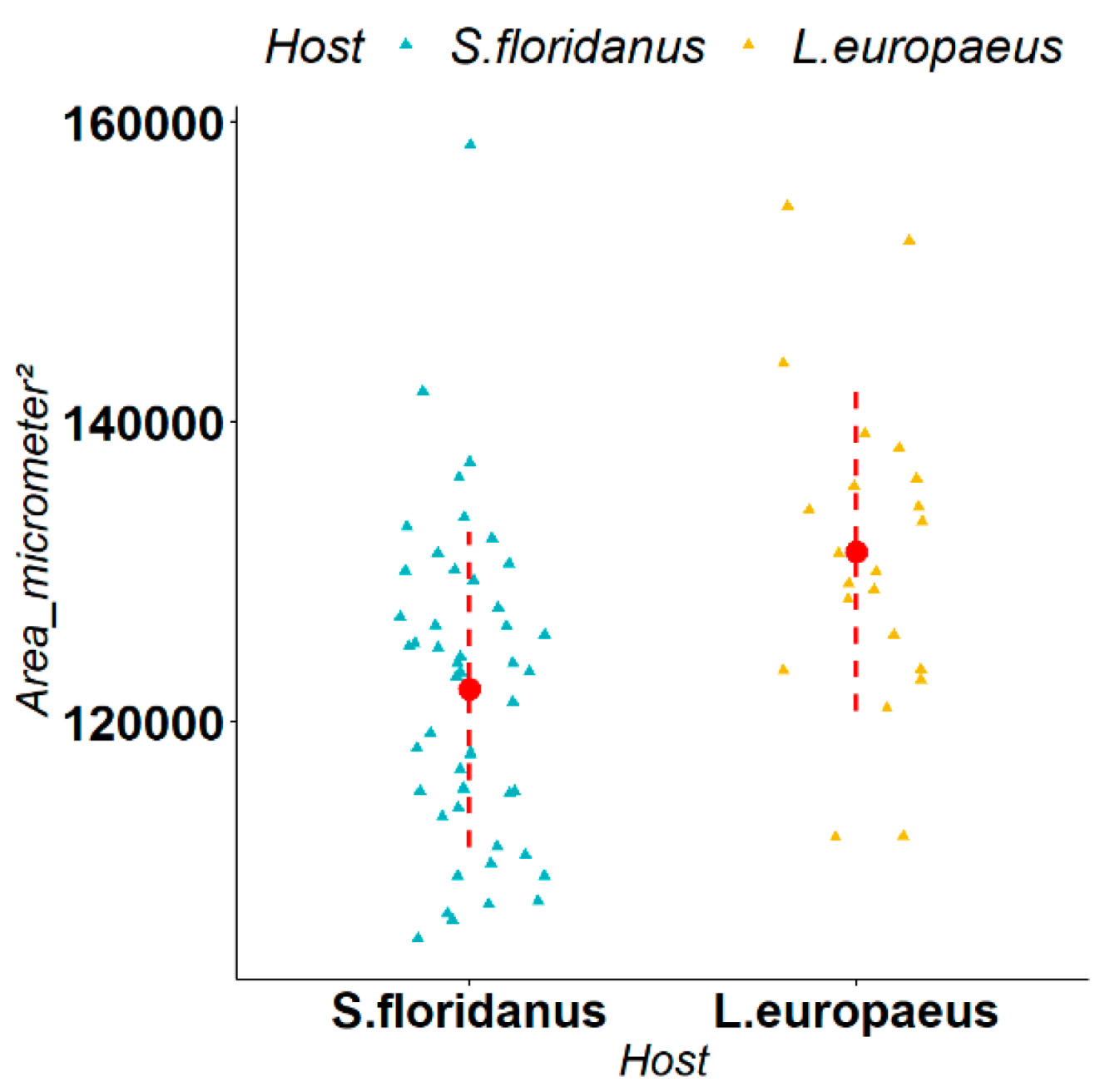
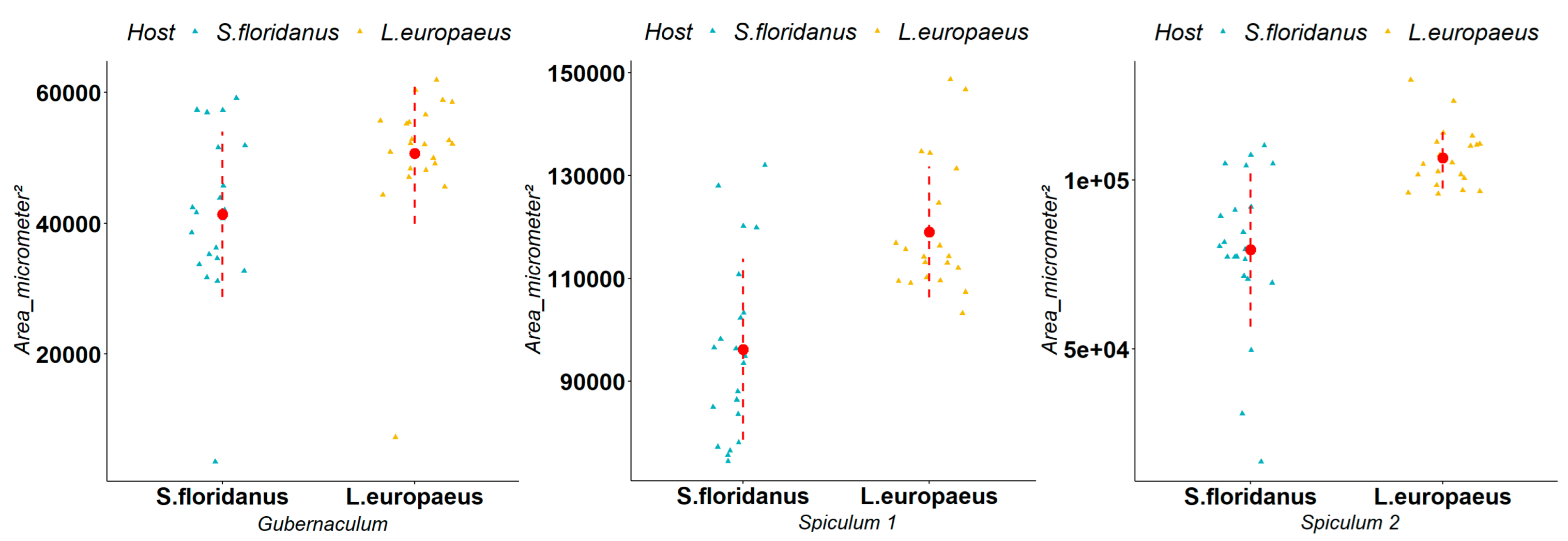
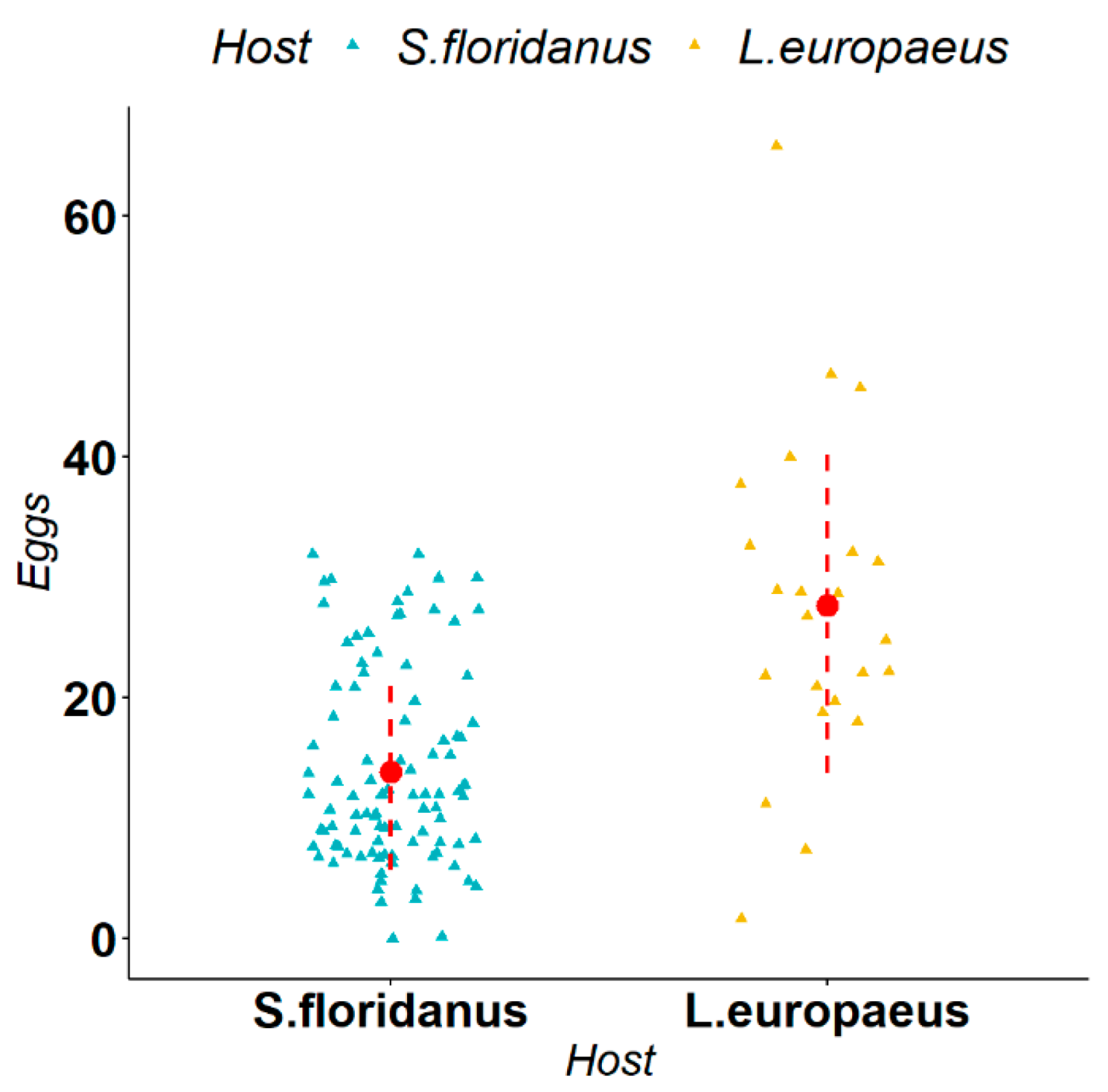
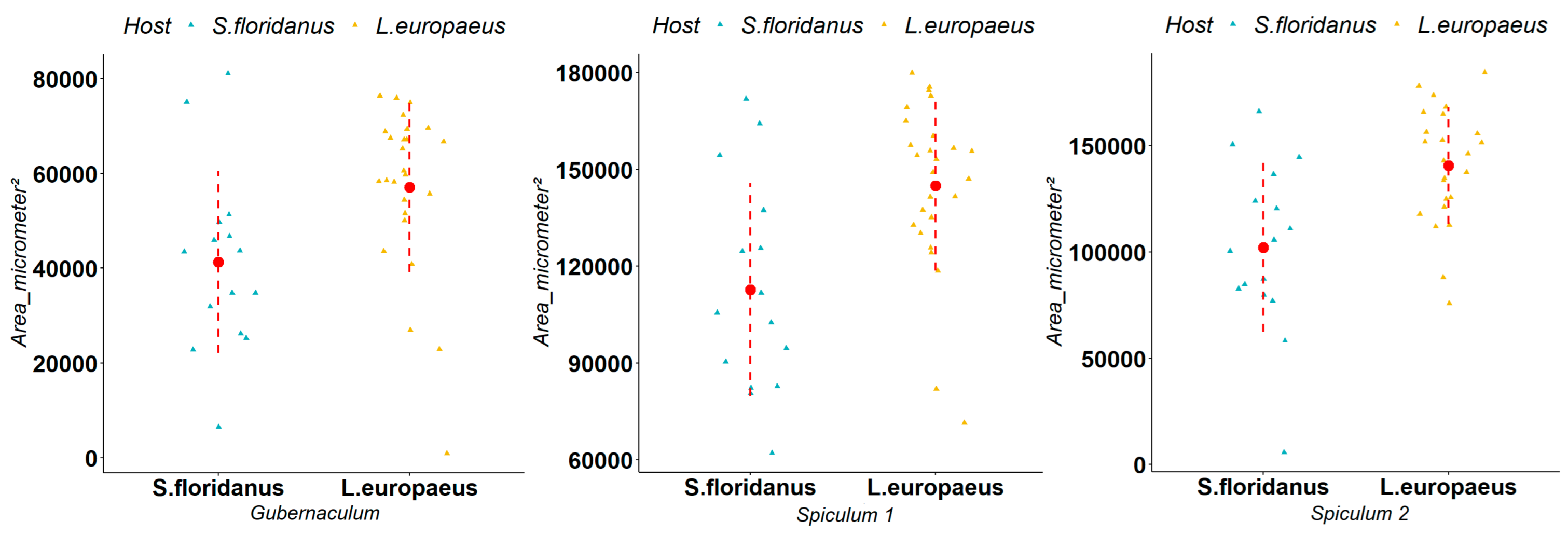
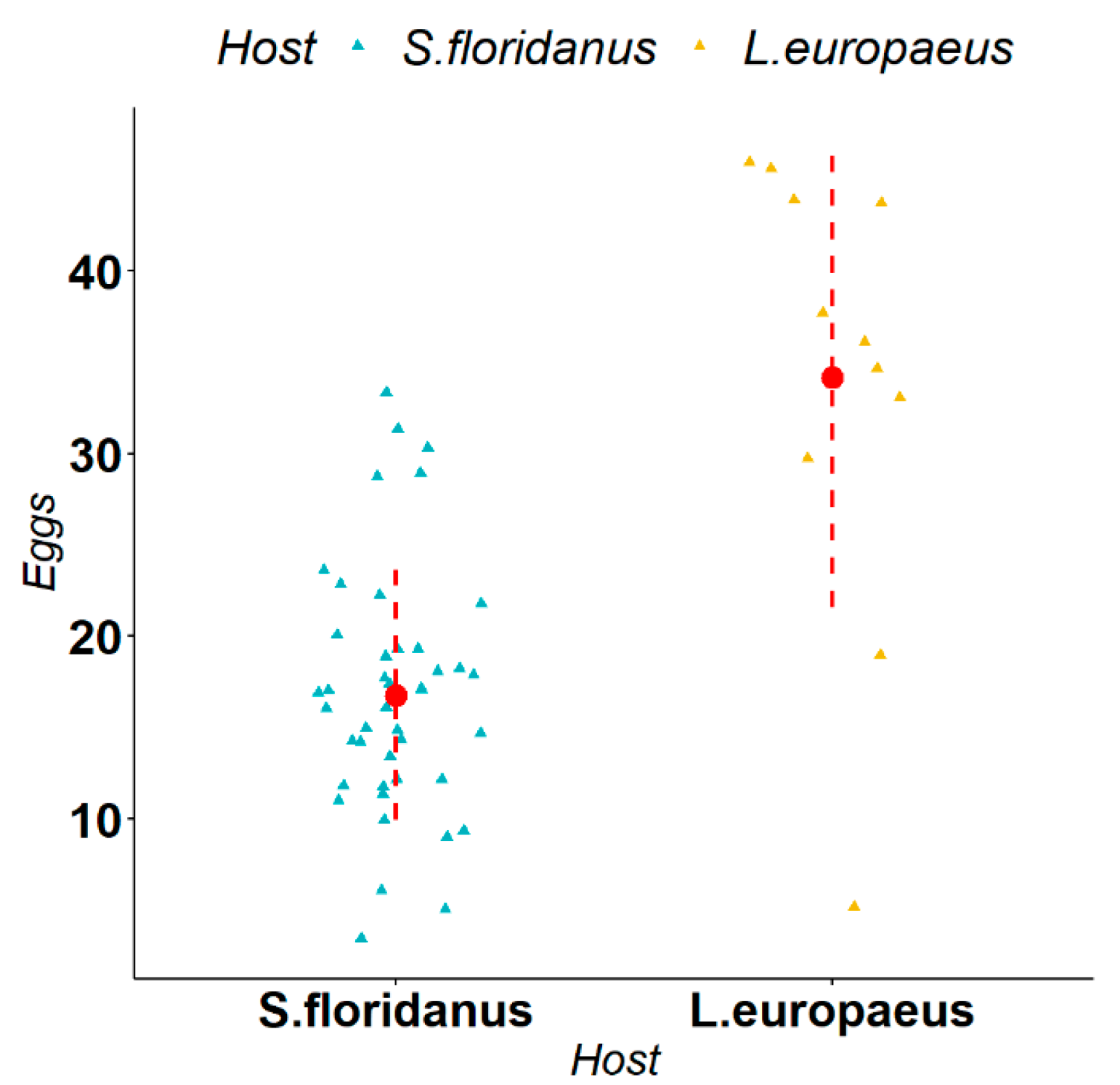
3.2. Biometric Measurements of Parasites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Convention on Biological Diversity (CBD), Glossary of Terms. Available online: https://www.cbd.int/invasive/terms.shtml (accessed on 17 March 2022).
- McNeely, J.A. The Great Reshuffling: Human Dimensions of Invasive Alien Species, 3rd ed.; McNeely, J.A., Ed.; World Conservation Union: Gland, Switzerland, 2001; ISBN 978-2831706023. [Google Scholar]
- Russell, J.C.; Towns, D.R.; Clout, M.N. Review of rat invasion biology: Implications for island biosecurity. Sci. Conserv. 2008, 286, 53. [Google Scholar]
- Massetti, M. Uomini e (Non Solo) Topi. Gli Animali Domestici e la Fauna Antropocora; Firenze University Press: Firenze, Italy, 2008; ISBN 978-8884538161. [Google Scholar]
- Cocchi, R.; Volponi, S.; Baccetti, N. Piano di Gestione Nazionale dell’Ibis Sacro Threskiornis aethiopicus (Latham, 1790); MATTM, ISPRA, SNPA: Rome, Italy, 2020. Available online: https://www.mite.gov.it/pagina/esiti-consultazione-pubblica-sette-piani-di-gestione (accessed on 17 March 2022).
- Nerva, L.; Iannucci, A.; Menchetti, M.; Andreoni, A.; Chitarra, W.; Martini, M.; Mueller, N.; Peeters, T.M.J.; Pesenti, E.; Verbeylen, G.; et al. Where do Chip and Dale come from? Origins of invasive populations of the Siberian chipmunk in Europe. Mamm. Res. 2021, 66, 525–529. [Google Scholar] [CrossRef]
- Scalera, R.; Bevilacqua, G.; Carnevali, L.; Genovesi, P. Le Specie Esotiche Invasive: Andamenti, Impatti e Possibili Risposte; ISPRA: Rome, Italy, 2018.
- Pârâu, L.G.; Strubbe, D.; Mori, E.; Menchetti, M.; Ancillotto, L.; van Kleunen, A.; White, R.L.; Luna, A.; Hernández-Brito, D.; Le Louarn, M.; et al. Rose-ringed Parakeet Psittacula krameri populations and numbers in Europe: A complete overview. Open Ornithol. J. 2016, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- IUCN. Guidance for Interpretation of the CBD Categories of Pathways for the Introduction of Invasive Alien Species; Technical Note Prepared by IUCN for the European Commission; European Commision Publications: Luxembourg, 2018. [CrossRef]
- Wauters, L.A.; Tosi, G.; Gurnell, J. A review of the competitive effects of alien grey squirrels on behaviour, activity and habitat use of red squirrels in mixed, deciduous woodland in Italy. Hystrix It. J. Mamm. 2005, 16, 27–40. [Google Scholar] [CrossRef]
- Zwerschke, N.; van Rein, H.; Harrod, C.; Reddin, C.; Emmerson, M.C.; Roberts, D.; O’Connor, N.E. Competition between co-occurring invasive and native consumers switches between habitats. Funct. Ecol. 2018, 32, 2717–2729. [Google Scholar] [CrossRef]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D. Hybridization between native and introduced wildlife species: Importance for conservation. Wildl. Biol. 1996, 2, 143–150. [Google Scholar] [CrossRef]
- Largiadèr, C.R. Hybridization and introgression between native and alien species. Biol. Invasions 2008, 193, 275–292. [Google Scholar] [CrossRef]
- Foster, R.; Peeler, E.; Bojko, J.; Clark, P.F.; Morritt, D.; Roy, H.E.; Stebbing, P.; Tidbury, H.J.; Wood, L.E.; Bass, D. Pathogens co-transported with invasive non-native aquatic species: Implications for risk analysis and legislation. NeoBiota 2021, 69, 79–102. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Cattadori, I.M.; Newborn, D.; Haydon, D.T.; Shaw, D.; Benton, T.G.; Grenfell, B.T. Trophic interactions and population growth rates: Describing patterns and identifying mechanisms. Phil. Trans. R. Soc. Lond. B 2002, 37, 1259–1272. [Google Scholar] [CrossRef]
- Dobson, A.P.; Hudson, P.J. Regulation and stability of a free-living host-parasite system, Trichostrongylus tenuis in red grouse. II. Population models. J. Anim. Ecol. 1992, 61, 487–498. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Newborn, D. Do parasites make prey vulnerable to predation? Red grouse and parasites. J. Anim. Ecol. 1992, 61, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Hudson, P.J.; Newborn, D.; Dobson, A.P. Regulation and stability of a free-living host-parasite system, Trichostrongylus tenuis in red grouse. I. Monitoring and parasite reduction experiments. J. Anim. Ecol. 1992, 61, 477–486. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Newborn, D. Population cycles and parasitism. Science 1999, 286, 2425. [Google Scholar] [CrossRef] [Green Version]
- Newey, S.; Thirgood, S.J.; Hudson, P.J. Do parasite burdens in spring influence condition and fecundity of female mountain hares Lepus timidus? Wildl. Biol. 2004, 10, 171–176. [Google Scholar] [CrossRef]
- Halvorsen, O.; Bye, K. Parasites, biodiversity and population dynamics in an ecosystem in the High Arctic. Vet. Parasitol. 1999, 84, 205–227. [Google Scholar] [CrossRef]
- Irvine, R.J.; Stien, A.; Halvorsen, O.; Langvatn, R.; Albon, S.D. Life-history strategies and population dynamics of abomasal nematodes in Svalbard reindeer (Rangifer tarandus platyrhynchus). Parasitology 2000, 120, 297–311. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife: Threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Kelly, D.W.; Paterson, R.A.; Townsend, C.R.; Poulin, R.; Tompkins, D.M. Parasite spillback: A neglected concept in invasion ecology? Ecology 2009, 90, 2047–2056. [Google Scholar] [CrossRef]
- Doria, G. Silvilago o minilepre Sylvilagus floridanus (Allen). In Animali e Piante dalle Americhe all’Europa; Cappadocia Orsini, L., Doria, G., Eds.; Sagep: Genova, Italy, 1991. [Google Scholar]
- Spagnesi, M. Silvilago Sylvilagus floridanus (J.A. Allen, 1890). In Quaderni di Conservazione della Natura. Mammiferi d’Italia; Spagnesi, M., De Marinis, A.M., Eds.; Ministero delle Politiche Agricole e Forestali e L’istituto Nazionale per la Fauna Selvatica: Rome, Italy, 2002; Volume 14, pp. 156–157. [Google Scholar]
- Andreotti, A.; Baccetti, N.; Perfetti, A.; Besa, M.; Genovesi, P.; Guberti, V. Mammiferi e uccelli esotici in Italia: Analisi del fenomeno, impatto sulla biodiversità e linee guida gestionali. In Quaderni di Conservazione della Natura; Ministero delle Politiche Agricole e Forestali e L’istituto Nazionale per la Fauna Selvatica: Rome, Italy, 2001; Volume 14. [Google Scholar]
- Tizzani, P.; Catalano, S.; Rossi, L.; Duignan, P.J.; Meneguz, P.G. Invasive species and their parasites: Eastern cottontail rabbit Sylvilagus floridanus and Trichostrongylus affinis (Graybill, 1924) from Northwestern Italy. Parasitol. Res. 2014, 113, 1301–1303. [Google Scholar] [CrossRef] [Green Version]
- Tizzani, P.; Andrade, D.; Molinar Min, A.R.; Peano, A.; Meneguz, P.G. Does the introduction of alien species represent a sanitary threat for native species? The case of the eastern cottontail Sylvilagus floridanus in Italy. Life 2020, 10, 142. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.K.; Vaughan Jennings, N.; Harris, S. A quantitative analysis of the abundance and demography of European hares Lepus europaeus in relation to habitat type, intensity of agriculture and climate. Mamm. Rev. 2005, 35, 1–24. [Google Scholar] [CrossRef]
- Malacarne, G.; Cucco, M.; Boano, G. Progetto di Piano Faunistico-Venatorio; Provincia di Alessandria: Alessandria, Italy, 1999.
- Canova, L.; Gazzola, A.; Pollini, L.; Balestrieri, A. Surveillance and habitat diversity affect European brown hare (Lepus europaeus) density in protected breeding areas. Eur. J. Wildl. Res. 2020, 66, 66. [Google Scholar] [CrossRef]
- Naldi, L.; Greco, I.; Ferretti, M.; Zaccaroni, M. Density estimates and habitat preferences of the European Hare (Lepus europaeus) on mountainous areas in Italy. Mammal Study 2020, 45, 123–131. [Google Scholar] [CrossRef]
- Silvano, F.; Acquarone, C.; Cucco, M. Distribution of the eastern cottontail Sylvilagus floridanus in the province of Alessandria. Hystrix It. J. Mamm. 2000, 66, 75–78. [Google Scholar] [CrossRef]
- Bertolino, S.; Ingegno, B.; Girardello, M. Modelling the habitat requirements of invasive Eastern Cottontail (Sylvilagus floridanus) introduced to Italy. Eur. J. Wildl. Res. 2011, 57, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Tizzani, P.; Dematteis, A. Linee Guida per la Gestione della Lepre Europea (Lepus europeus); Provincia di Cuneo: Cuneo, Italy, 2009.
- Skrjabin, K.I.; Shikhobalova, N.P.; Schulz, R.S. (Eds.) Essential of Nematodology. In Trichostrongylids of Animals and Man; The Academy of Sciences of the USSR: Moscow, Russia, 1954; Volume 3. [Google Scholar]
- QGIS.org. QGIS Geographic Information System. QGIS Association. 2022. Available online: http://www.qgis.org (accessed on 17 March 2022).
- Tizzani, P.; Catalano, S.; Rossi, L.; Meneguz, P.G. A new tool for the measurement of parasitic structures: A GIS approach. Parassitologia 2010, 52, 335. [Google Scholar]
- Margolis, L.; Esch, G.W.; Holmes, J.C.; Kuris, A.M.; Schad, G.A. The use of ecological terms in parasitology (report of an ad hoc committee of the American Society of Parasitologists). J. Parasitol. 1982, 68, 131–133. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, 2020. Available online: https://www.R-project.org/ (accessed on 15 March 2022).
- Kassambara, A. Ggpubr: ‘Ggplot2′ Based Publication Ready Plots. R Package Version 0.4.0. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 15 March 2022).
- Chen, H. VennDiagram: Generate High-Resolution Venn and Euler Plots. R Package Version 1.7.1. 2021. Available online: https://CRAN.R-project.org/package=VennDiagram (accessed on 15 March 2022).
- Smith, K.F.; Sax, D.F.; Lafferty, K.D. Evidence for the role of infectious disease in species extinction and endangerment. Conserv. Biol. 2006, 20, 1349–1357. [Google Scholar] [CrossRef]
- Delogu, M.; Ghetti, G.; Gugiatti, A.; Cotti, C.; Piredda, I.; Frasnelli, M.; De Marco, M.A. Virological investigation of avian influenza virus on postglacial species of Phasianidae and Tetraonidae in the Italian Alps. Int. Sch. Res. Not. 2013, 2013, 601732. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, R.; Melville, L.; Sargison, F.; Kenyon, F.; Nussey, D.; Watt, K.; Sargison, K. Gastrointestinal nematode species diversity in Soay sheep kept in a natural environment without active parasite control. Vet. Parasitol. 2016, 227, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Leivesley, J.A.; Bussière, L.F.; Pemberton, J.M.; Pilkington, J.G.; Wilson, K.; Hayward, A.D. Survival costs of reproduction are mediated by parasite infection in wild Soay sheep. Ecol. Lett. 2019, 22, 1203–1213. [Google Scholar] [CrossRef]
- Iacopelli, F.; Fanelli, A.; Tizzani, P.; Berriatua, E.; Prieto, P.; Martínez-Carrasco, C.; León, L.; Rossi, L.; Candela, M.G. Spatio-temporal patterns of sarcoptic mange in red deer and Iberian ibex in a multi-host natural park. Res. Vet. Sci. 2020, 128, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Carrau, T.; Martínez-Carrasco, C.; Garijo, M.M.; Alonso, F.; de Ybáñez, R.R.; Tizzani, P. Multivariate abundance analysis of multi-host/multi-parasite lungworms in a sympatric wild ruminant population. Diversity 2021, 13, 227. [Google Scholar] [CrossRef]
- Carrau, T.; Martínez-Carrasco, C.; Garijo, M.M.; Alonso, F.; Vizcaíno, L.L.; Herrera-Russert, J.; Tizzani, P.; de Ybáñez, R.R. Epidemiological approach to nematode polyparasitism occurring in a sympatric wild ruminant multi-host scenario. J. Helminthol. 2021, 95, E29. [Google Scholar] [CrossRef]
- Fanelli, A.; Menardi, G.; Chiodo, M.; Giordano, O.; Ficetto, G.; Bessone, M.; Lasagna, A.; Carpignano, M.G.; Min, A.M.; Gugiatti, A.; et al. Gastroenteric parasite of wild Galliformes in the Italian Alps: Implication for conservation management. Parasitology 2020, 147, 471–477. [Google Scholar] [CrossRef]
- Tizzani, P.; Menzano, A.; Catalano, S.; Rossi, L.; Meneguz, P.G. First report of Obeliscoides cuniculi in European brown hare (Lepus europaeus). Parasitol. Res. 2011, 109, 963–966. [Google Scholar] [CrossRef]
- Alzaga, V.; Tizzani, P.; Acevedo, P.; Ruiz-Fons, F.; Vicente, J.; Gortázar, C. Deviance partitioning of host factors affecting parasitization in the European brown hare (Lepus europaeus). Naturwissenschaften 2009, 96, 1157–1168. [Google Scholar] [CrossRef]
- Gontero, C.; Fanelli, A.; Zanet, S.; Meneguz, P.G.; Tizzani, P. Exotic species and autochthonous parasites: Trichostrongylus retortaeformis in Eastern cottontail. Life 2020, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Soveri, T.; Valtonern, M. Endoparasites of hares (Lepus timidus L. and L. europaeus Pallas) in Finland. J. Wild. Dis. 1983, 19, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordes, F.; Langand, J.; Feliu, C.; Morand, S. Helminth communities of an introduced hare (Lepus granatensis) and a native hare (Lepus europaeus) in southern France. J. Wild. Dis. 2007, 43, 747–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; McKenzie, V.J.; Kuris, A.M. Introduced species and their missing parasites. Nature 2003, 421, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Kołodziej-Sobocińska, M. Factors affecting the spread of parasites in populations of wild European terrestrial mammals. Mamm. Res. 2019, 64, 301–318. [Google Scholar] [CrossRef] [Green Version]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: New York, NY, USA, 1992; ISBN 978-0198577416. [Google Scholar]
- Poulin, R. Evolutionary Ecology of Parasites, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007; ISBN 9781400840809. [Google Scholar]
- Keymer, A.E. Density-dependent mechanisms in the regulation of the intestinal helminth populations. Parasitology 1982, 84, 573–587. [Google Scholar] [CrossRef]
- Shostak, A.W.; Dick, T.A. Individual variability in reproductive success of Triaenophorus crassus Forel (Cestoda: Pseudophyllidea), with comments on the use of the Lorenz curve and Gini coefficient. Can. J. Zool. 1987, 65, 2878–2885. [Google Scholar] [CrossRef]
- Szalai, A.J.; Dick, T.A. Differences in numbers and inequalities in mass and fecundity during the egg-producing period for Raphidascaris acus (Nematoda: Anisakidae). Parasitology 1989, 98, 489–495. [Google Scholar] [CrossRef]
- Dove, A.D.M. Richness patterns in the parasite communities of exotic poeciliid fishes. Parasitology 2000, 120, 609–623. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M. Sudden oak death: Endangering California and Oregon forest ecosystems. Front. Ecol. Environ. 2003, 1, 197–204. [Google Scholar] [CrossRef]
- De Castro, F.; Bolker, B. Mechanisms of disease-induced extinction. Ecol. Lett. 2005, 8, 117–126. [Google Scholar] [CrossRef]
- Tompkins, D.M.; Sainsbury, A.W.; Nettleton, P.; Buxton, D.; Gurnell, J. Parapoxvirus causes a deleterious disease in red squirrels associated with UK population declines. Proc. Biol. Sci. 2002, 269, 529–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tompkins, D.M.; White, A.R.; Boots, M. Ecological replacement of native red squirrels by invasive greys driven by disease. Ecol. Lett. 2003, 6, 189–196. [Google Scholar] [CrossRef]
- Rushton, S.P.; Lurz, P.W.W.; Gurnell, J.; Nettleton, P.; Bruemmer, C.; Shirley, M.D.F.; Sainsbury, A.W. Disease threats posed by alien species: The role of a poxvirus in the decline of the native red squirrel in Britain. Epidemiol. Infect. 2006, 134, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Romeo, C.; Piscitelli, A.P.; Santicchia, F.; Martinoli, A.; Ferrari, N.; Wauters, L.A. Invading parasites: Spillover of an alien nematode reduces survival in a native species. Biol. Invasions 2021, 23, 3847–3857. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Prevalence % | Abundance | Intensity | |
|---|---|---|---|---|
| Lepus europaeus (Alessandria—N = 21) | Obeliscoides cuniculi | 40.0 [16.8–68.7] | 10.5 [−3.7–24.7] | 26.2 [−13.4–26.2] |
| Trichostrongylus calcaratus | 33.3 [17.2–54.6] | 27.3 [−0.4–55.0] | 39 [0.9–77.1] | |
| Trichostrongylus retortaeformis | 19.0 [7.7–40.0] | 15.5 [−0.3–31.3] | 77.5 [31.8–123.2] | |
| Trichostrongylus affinis | 25.0 [11.2–46.9] | 19.3 [−8.3–46.9] | 77.2 [−52.7–207.1] | |
| Trichuris leporis | 20.0 [8.1–41.6] | 0.3 [0–0.7] | 1.8 [0.3–3.2] | |
| Trichostrongylus colubriformis | 4.8 [0.8–22.7] | 13 [NA] | 260 [NA] | |
| Sylvilagus floridanus (Alessandria—N = 16) | Obeliscoides cuniculi | 86.7 [62.1–96.3] | 96.8 [−14.2–207.8] | 149 [−22.6–320.6] |
| Trichostrongylus calcaratus | 85.7 [60.1–96.0] | 963 [117–1809] | 1123.5 [151.5–2095.5] | |
| Trichostrongylus affinis | 68.8 [44.4–85.8] | 79.1 [−11.6–169.8] | 115 [−17.6–247.6] | |
| Lepus timidus varronis (Cuneo—N = 7) | Trichostrongylus retortaeformis | 42.9 [15.8–74.9] | 1.4 [−1.0–3.8] | 3.3 [−4.6–11.2] |
| Trichuris leporis | 14.3 [2.6–51.3] | 0.7 [NA] | 5 [NA] | |
| Trichostrongylus colubriformis | 14.3 [2.6–51.3] | 26.1 [NA] | 183 [NA] | |
| Cestoda | 71.4 [35.9–91.8] | 91.7 [−49.7–233.1] | 128.4 [−83.7–340.5] | |
| Nematodirus sp. | 14.3 [2.6–51.3] | 0.57 [NA] | 4 [NA] | |
| Lepus europaeus (Cuneo—N = 30) | Obeliscoides cuniculi | 6.7 [1.8–21.3] | 0.4 [−0.4–1.2] | 6.5 [−63.6–76.6] |
| Trichostrongylus calcaratus | 3.3 [0.6–16.7] | 0.2 [NA] | 7 [NA] | |
| Trichostrongylus retortaeformis | 40.0 [24.6–57.9] | 12.3 [2.6–22.0] | 30.8 [9.2–52.4] | |
| Trichostrongylus affinis | 3.3 [0.6–16.7] | 1 [NA] | 30 [NA] | |
| Trichuris leporis | 40.0 [24.6–57.9] | 5.6 [1.3–9.9] | 13.9 [4.5–23.3] | |
| Trichostrongyluscolubriformis | 3.3 [0.6–16.7] | 14.3 [NA] | 430 [NA] | |
| Cestoda | 26.7 [14.2–44.4] | 2.9 [−0.2–6.0] | 10.7 [−1.1–22.5] | |
| Passalurus Ambiguus | 13.3 [5.3–29.7] | 26.4 [−14.4–67.0] | 197.7 [–223.5–618.9] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catalano, S.; La Morgia, V.; Molinar Min, A.R.; Fanelli, A.; Meneguz, P.G.; Tizzani, P. Gastrointestinal Parasite Community and Phenotypic Plasticity in Native and Introduced Alien Lagomorpha. Animals 2022, 12, 1287. https://doi.org/10.3390/ani12101287
Catalano S, La Morgia V, Molinar Min AR, Fanelli A, Meneguz PG, Tizzani P. Gastrointestinal Parasite Community and Phenotypic Plasticity in Native and Introduced Alien Lagomorpha. Animals. 2022; 12(10):1287. https://doi.org/10.3390/ani12101287
Chicago/Turabian StyleCatalano, Stefano, Valentina La Morgia, Anna Rita Molinar Min, Angela Fanelli, Pier Giuseppe Meneguz, and Paolo Tizzani. 2022. "Gastrointestinal Parasite Community and Phenotypic Plasticity in Native and Introduced Alien Lagomorpha" Animals 12, no. 10: 1287. https://doi.org/10.3390/ani12101287
APA StyleCatalano, S., La Morgia, V., Molinar Min, A. R., Fanelli, A., Meneguz, P. G., & Tizzani, P. (2022). Gastrointestinal Parasite Community and Phenotypic Plasticity in Native and Introduced Alien Lagomorpha. Animals, 12(10), 1287. https://doi.org/10.3390/ani12101287