
Growth Curves and Body Condition of Young Cats and Their Relation to Maternal Body Condition


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Data Collection
2.3. Statistical Analysis
3. Results
3.1. Relationship of BCS at 8 Months and Maternal Phenotype to Weight Development
3.1.1. General
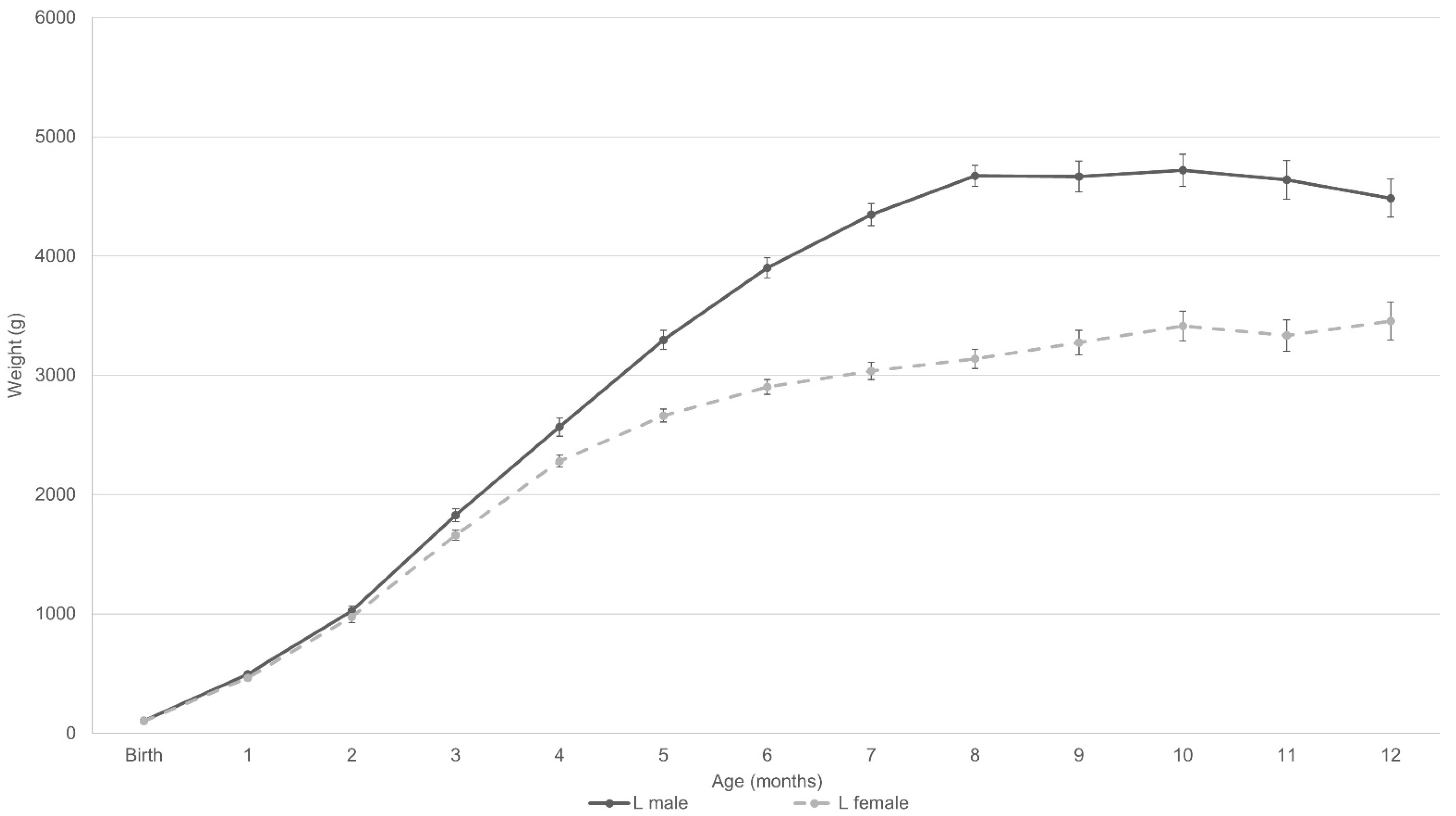
3.1.2. According to Sex
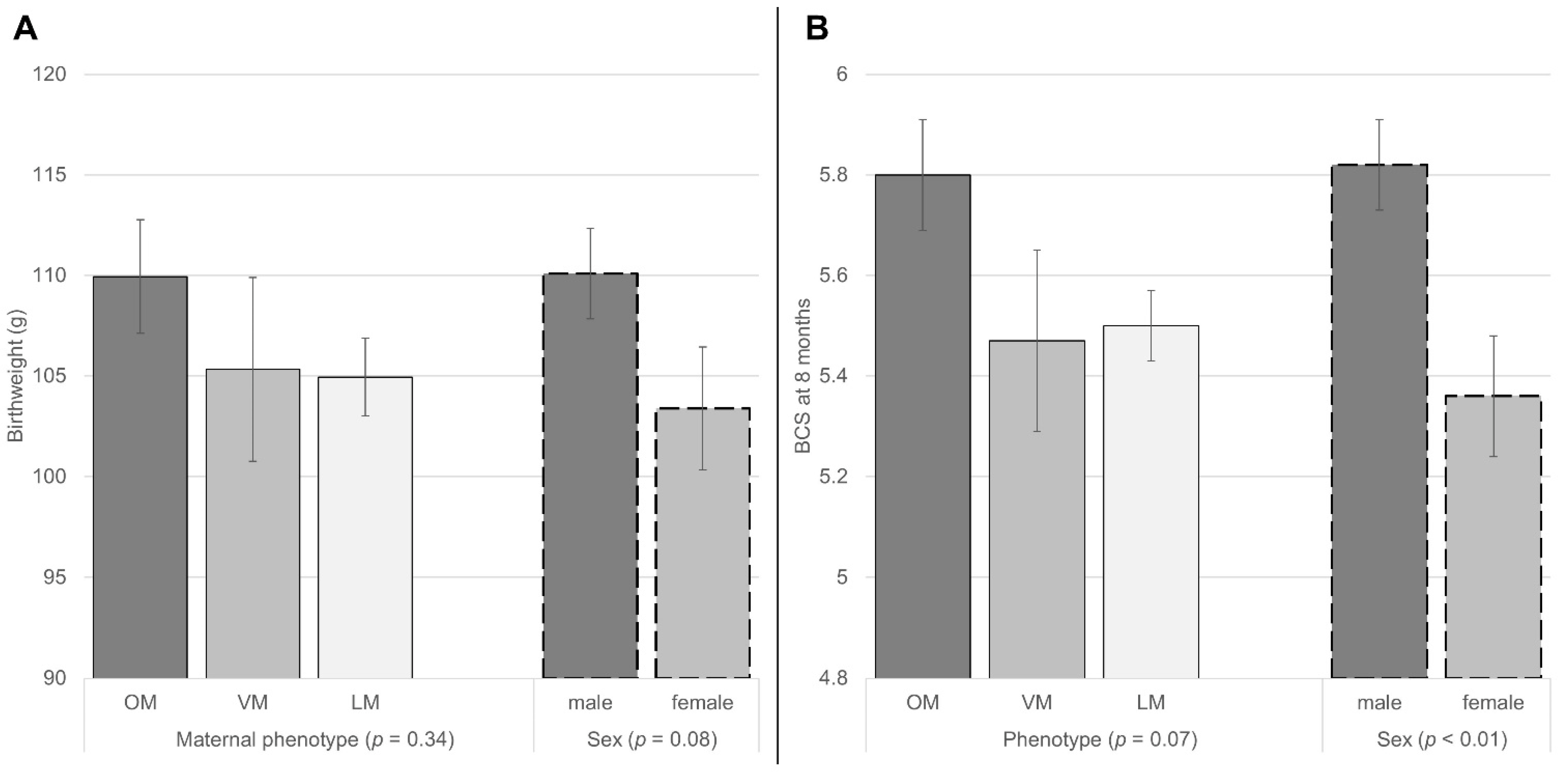
3.2. Relationship of Litter Traits and Birthweight with Kitten’s BCS at 8 Months
3.3. Relationship of Sex and Maternal Phenotype to Litter Characteristics, Birthweight and BCS at 8 Months
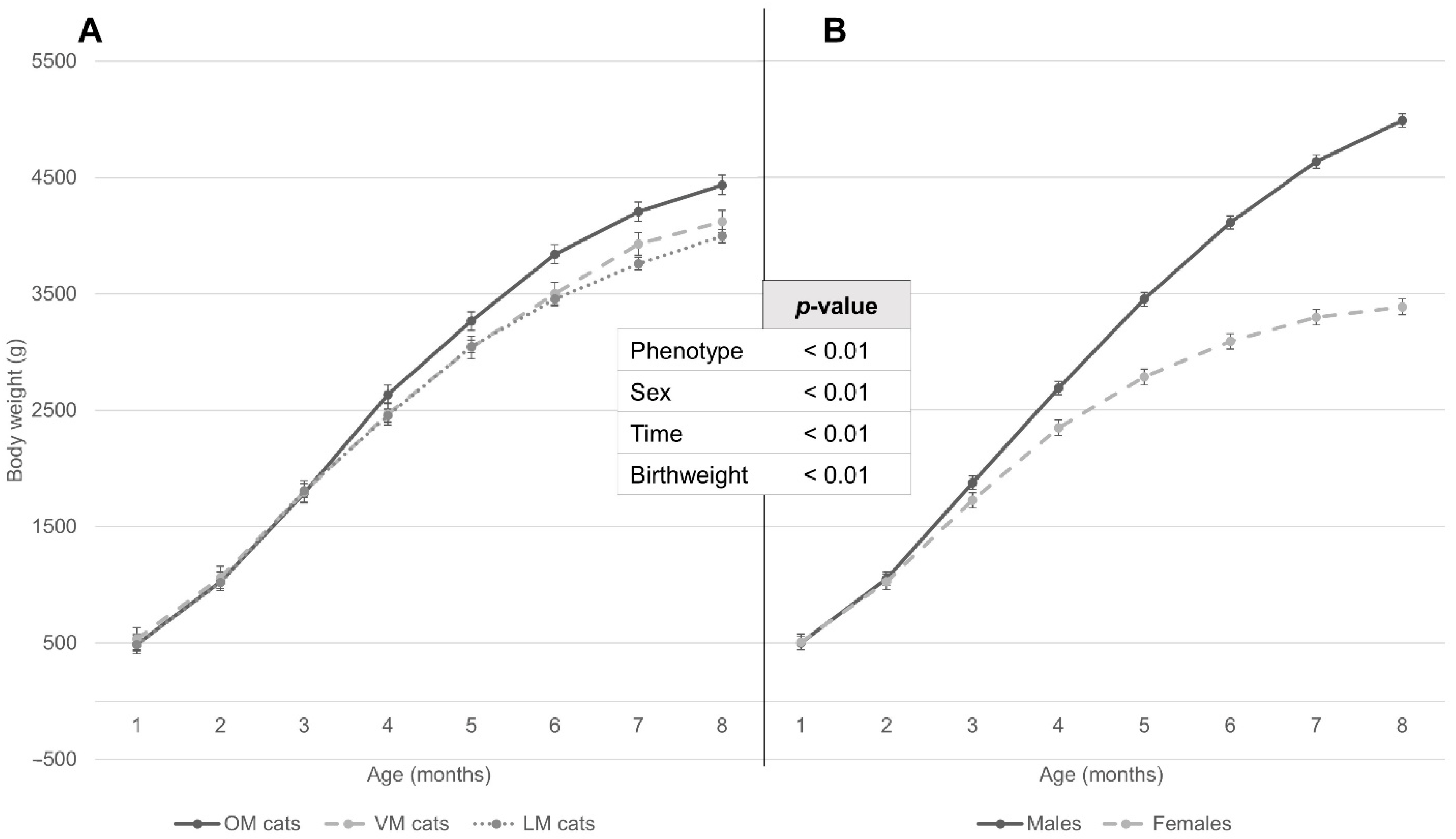
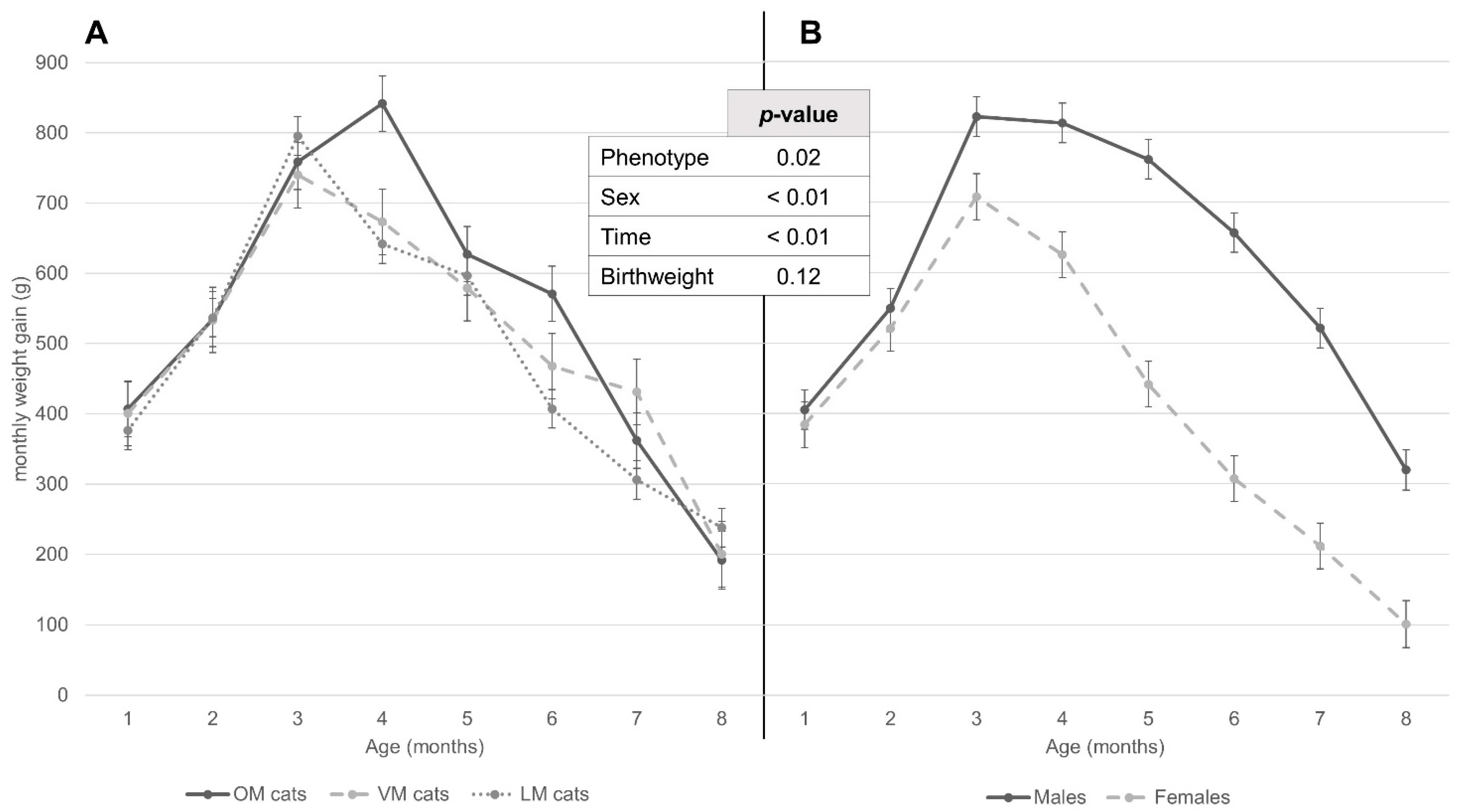
3.4. Weight Development according to Repeated Mixed Measures Model in Relation to Sex and Maternal Phenotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teng, K.T.; McGreevy, P.D.; Toribio, J.A.L.M.L.; Raubenheimer, D.; Kendall, K.; Dhand, N.K. Associations of Body Condition Score with Health Conditions Related to Overweight and Obesity in Cats. J. Small Anim. Pract. 2018, 59, 603–615. [Google Scholar] [CrossRef] [PubMed]
- German, A.J. The Growing Problem of Obesity in Dogs and Cats. J. Nutr. 2006, 136, 1940S–1946S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.M. The Growing Problem of Obesity: Mechanisms, Consequences, and Therapeutic Approaches. Arterioscler. Thromb. Vasc. Biol. 2015, 35, e19–e23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarkosova, D.; Story, M.M.; Rand, J.S.; Svoboda, M. Feline Obesity-Prevalence, Risk Factors, Pathogenesis, Associated Conditions and Assessment: A Review. Vet. Med. 2016, 61, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Deagle, G.; Holden, S.L.; Biourge, V.; Morris, P.J.; German, A.J. Long-Term Follow-up after Weight Management in Obese Cats. J. Nutr. Sci. 2014, 3, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kienzle, E.; Bergler, R. Human-Animal Relationship of Owners of Normal and Overweight Cats. J. Nutr. 2006, 136, 1947S–1950S. [Google Scholar] [CrossRef] [Green Version]
- Colliard, L.; Paragon, B.-M.; Lemuet, B.; Bénet, J.-J.; Blanchard, G. Prevalence and Risk Factors of Obesity in an Urban Population of Healthy Cats. J. Feline Med. Surg. 2009, 11, 135–140. [Google Scholar] [CrossRef]
- Cave, N.J.; Allan, F.J.; Schokkenbroek, S.L.; Metekohy, C.A.M.; Pfeiffer, D.U. A Cross-Sectional Study to Compare Changes in the Prevalence and Risk Factors for Feline Obesity between 1993 and 2007 in New Zealand. Prev. Vet. Med. 2012, 107, 121–133. [Google Scholar] [CrossRef]
- Rowe, E.C.; Browne, W.J.; Casey, R.A.; Gruffydd-Jones, T.J.; Murray, J.K. Early-Life Risk Factors Identified for Owner-Reported Feline Overweight and Obesity at around Two Years of Age. Prev. Vet. Med. 2017, 143, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Wall, M.; Cave, N.J.; Vallee, E. Owner and Cat-Related Risk Factors for Feline Overweight or Obesity. Front. Vet. Sci. 2019, 6, 266. [Google Scholar] [CrossRef]
- Larsen, J.A. Risk of Obesity in the Neutered Cat. J. Feline Med. Surg. 2017, 19, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Fettman, M.J.; Stanton, C.A.; Banks, L.L.; Hamar, D.W.; Johnson, D.E.; Hegstad, R.L.; Johnston, S. Effects of Neutering on Bodyweight, Metabolic Rate and Glucose Tolerance of Domestic Cats. Res. Vet. Sci. 1997, 62, 131–136. [Google Scholar] [CrossRef]
- Flynn, M.F.; Hardie, E.M.; Armstrong, P.J. Effect of Ovariohysterectomy on Maintenance Energy Requirement in Cats. J. Am. Vet. Med. Assoc. 1996, 209, 1572–1581. [Google Scholar] [PubMed]
- Courcier, E.A.; Mellor, D.J.; Pendlebury, E.; Evans, C.; Yam, P.S. An Investigation into the Epidemiology of Feline Obesity in Great Britain: Results of a Cross-Sectional Study of 47 Companion Animal Practises. Vet. Rec. 2012, 171, 560. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.D.; Poston, L. Developmental Programming of Obesity in Mammals. Exp. Physiol. 2007, 92, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Barness, L.A.; Opitz, J.M.; Gilbert-Barness, E. Obesity: Genetic, Molecular, and Environmental Aspects. Am. J. Med. Genet. 2007, 143, 3016–3034. [Google Scholar] [CrossRef]
- Cave, N.J.; Bridges, J.P.; Weidgraaf, K.; Thomas, D.G. Nonlinear Mixed Models of Growth Curves from Domestic Shorthair Cats in a Breeding Colony, Housed in a Seasonal Facility to Predict Obesity. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1390–1400. [Google Scholar] [CrossRef]
- Yu, Z.B.; Han, S.P.; Zhu, G.Z.; Zhu, C.; Wang, X.J.; Cao, X.G.; Guo, X.R. Birth Weight and Subsequent Risk of Obesity: A Systematic Review and Meta-Analysis. Obes. Rev. 2011, 12, 525–542. [Google Scholar] [CrossRef]
- Gondret, F.; Lefaucheur, L.; Juin, H.; Louveau, I.; Lebret, B. Low Birth Weight Is Associated with Enlarged Muscle Fiber Area and Impaired Meat Tenderness of the Longissimus Muscle in Pigs. J. Anim. Sci. 2006, 84, 93–103. [Google Scholar] [CrossRef]
- Mugnier, A.; Morin, A.; Cellard, F.; Devaux, L.; Delmas, M.; Adib-Lesaux, A.; Flanagan, J.; Laxalde, J.; Chastant, S.; Grellet, A. Association between Birth Weight and Risk of Overweight at Adulthood in Labrador Dogs. PLoS ONE 2020, 15, e0243820. [Google Scholar] [CrossRef]
- Parra-Vargas, M.; Ramon-Krauel, M.; Lerin, C.; Jimenez-Chillaron, J.C. Size Does Matter: Litter Size Strongly Determines Adult Metabolism in Rodents. Cell Metab. 2020, 32, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Mozeš, Š.; Šefčíková, Z.; Raček, L. Long-Term Effect of Altered Nutrition Induced by Litter Size Manipulation and Cross-Fostering in Suckling Male Rats on Development of Obesity Risk and Health Complications. Eur. J. Nutr. 2014, 53, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Tummaruk, P.; Lundeheim, N.; Einarsson, S.; Dalin, A.M. Effect of Birth Litter Size, Birth Parity Number, Growth Rate, Backfat Thickness and Age at First Mating of Gilts on Their Reproductive Performance as Sows. Anim. Reprod. Sci. 2001, 66, 225–237. [Google Scholar] [CrossRef]
- Estany, J.; Villalba, D.; Tibau, J.; Soler, J.; Babot, D.; Noguera, J.L. Correlated Response to Selection for Litter Size in Pigs: I. Growth, Fat Deposition, and Feeding Behavior Traits. J. Anim. Sci. 2002, 80, 2556–2565. [Google Scholar] [CrossRef]
- Serisier, S.; Feugier, A.; Venet, C.; Biourge, V.; German, A.J. Faster Growth Rate in Ad Libitum-Fed Cats: A Risk Factor Predicting the Likelihood of Becoming Overweight during Adulthood. J. Nutr. Sci. 2013, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, A.H.; Rogers, K.; Henley, W.E.; Gunn-Moore, D.A.; May, J.M.; Gruffydd-Jones, T.J.; Bessant, C. A Questionnaire-Based Study of Gestation, Parturition and Neonatal Mortality in Pedigree Breeding Cats in the UK. J. Feline Med. Surg. 2006, 8, 145–157. [Google Scholar] [CrossRef]
- Merenda, M.E.Z.; Sato, J.; Scheibel, S.; Uemoto, A.T.; Rossoni, D.F.; dos Santos, M.P.; Pereira, L.C.; Ribeiro, L.B.; Vasconcellos, R.S. Growth Curve and Energy Intake in Male and Female Cats. Top. Companion Anim. Med. 2021, 44, 100518. [Google Scholar] [CrossRef]
- Gatel, L.; Rosset, E.; Chalvet-Monfray, K.; Buff, S.; Rault, D.N. Relationships between Fetal Biometry, Maternal Factors and Birth Weight of Purebred Domestic Cat Kittens. Theriogenology 2011, 76, 1716–1722. [Google Scholar] [CrossRef]
- Gast, A.-S. Approche Zootechnique de La Croissance Du Chaton. Ph.D. Thesis, École Nationale Vétérinaire d’Alfort, Maisons-Alfort, France, 2011. [Google Scholar]
- Häring, T.; Wichert, B.; Dolf, G.; Haase, B. Segregation Analysis of Overweight Body Condition in an Experimental Cat Population. J. Hered. 2011, 102, S28–S31. [Google Scholar] [CrossRef]
- FEDIAF. Nutritional Guidelines for Complete and Complementary Pet Food for Cats and Dogs; The European Pet Food Industry Federation: Bruxelles, Belgium, 2021. [Google Scholar]
- Laflamme, D. Development and Validation of a Body Condition Score System for Cats: A Clinical Tool. Feline Pract. 1997, 25, 13–18. [Google Scholar]
- Ghielmetti, V.; Wichert, B.; Rüegg, S.; Frey, D.; Liesegang, A. Food Intake and Energy Expenditure in Growing Cats with and without a Predisposition to Overweight. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1401–1410. [Google Scholar] [CrossRef] [PubMed]
- van Lent, D.; Vernooij, J.C.M.; Corbee, R.J. Kittens That Nurse 7 Weeks or Longer Are Less Likely to Become Overweight Adult Cats. Animals 2021, 11, 3434. [Google Scholar] [CrossRef] [PubMed]
- Moik, K.; Kienzle, E. Birth Weight and Postnatal Growth of Pure-Bred Kittens. Br. J. Nutr. 2011, 106, S32–S34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugnier, A.; Chastant, S.; Saegerman, C.; Gaillard, V.; Grellet, A.; Mila, H. Management of Low Birth Weight in Canine and Feline Species: Breeder Profiling. Animals 2021, 11, 2953. [Google Scholar] [CrossRef]
- Van De Velde, H.; Janssens, G.P.J.; De Rooster, H.; Polis, I.; Peters, I.; Ducatelle, R.; Nguyen, P.; Buyse, J.; Rochus, K.; Xu, J.; et al. The Cat as a Model for Human Obesity: Insights into Depot-Specific Inflammation Associated with Feline Obesity. Br. J. Nutr. 2013, 110, 1326–1335. [Google Scholar] [CrossRef] [Green Version]
- Hoenig, M. The Cat as a Model for Human Obesity and Diabetes. J. Diabetes Sci. Technol. 2012, 6, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Kil, D.Y.; Swanson, K.S. Endocrinology of Obesity. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 205–219. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Sex Differences in the Physiology of Eating. Am. J. Physiol.Regul. Integr. Comp. Physiol. 2013, 305, 1215–1267. [Google Scholar] [CrossRef] [Green Version]
- Olson, P.N.; Husted, P.W.; Allen, T.A.; Nett, T.M. Reproductive Endocrinology and Physiology of the Bitch and Queen. Vet. Clin. N. Am. Small Anim. Pract. 1984, 14, 927–946. [Google Scholar] [CrossRef]
- Feldman, E.C.; Nelson, R.W. Canine and Feline Endocrinology and Reproduction, 3rd ed.; Saunders: St. Louis, MO, USA, 2004; ISBN 0721693156. [Google Scholar]
- Axnér, E. Updates on Reproductive Physiology, Genital Diseases and Artificial Insemination in the Domestic Cat. Reprod. Domest. Anim. 2008, 43, 144–149. [Google Scholar] [CrossRef]
- Asarian, L.; Bächler, T. Neuroendocrine Control of Satiation. Horm. Mol. Biol. Clin. Investig. 2014, 19, 163–192. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, N.B.; Monaghan, P. Compensation for a Bad Start: Grow Now, Pay Later? Trend Ecol. Evol. 2001, 16, 254–260. [Google Scholar] [CrossRef]
- Bazhan, N.; Zelena, D. Food-Intake Regulation during Stress by the Hypothalamo-Pituitary-Adrenal Axis. Brain Res. Bull. 2013, 95, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Hendriks, W.H.; Wamberg, S. Milk Intake of Suckling Kittens Remains Relatively Constant from One to Four Weeks of Age. J. Nutr. 2000, 130, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Owen, C.G.; Martin, R.M.; Whincup, P.H.; Smith, G.D.; Cook, D.G. Effect of Infant Feeding on the Risk of Obesity across the Life Course: A Quantitative Review of Published Evidence. Pediatrics 2005, 115, 1367–1377. [Google Scholar] [CrossRef]
- Iben, C.; Liesegang, A.; Wichert, B.; Wolf, P. Allgemeine Grundlagen. In Ernährung der Katze; Iben, C., Liesegang, A., Wichert, B., Wolf, P., Eds.; Thieme: Stuttgart, Germany, 2021; pp. 15–86. [Google Scholar]
- Dobenecker, B.; Kienzle, E.; Zentek, J. Milk Yield and Milk Composition of Lactating Queens. J. Anim. Physiol. Anim. Nutr. 1998, 80, 173–178. [Google Scholar] [CrossRef]
- Chester-Jones, H.; Heins, B.J.; Ziegler, D.; Schimek, D.; Schuling, S.; Ziegler, B.; de Ondarza, M.B.; Sniffen, C.J.; Broadwater, N. Relationships between Early-Life Growth, Intake, and Birth Season with First-Lactation Performance of Holstein Dairy Cows. J. Dairy Sci. 2017, 100, 3697–3704. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.; Denk, H.; Bayram, D. Effects of Lambing Season, Sex and Birth Type on Growth Performance in Norduz Lambs. Small Rumin. Res. 2007, 68, 336–339. [Google Scholar] [CrossRef]
- Phillips, D.I.W.; Young, J.B. Birth Weight, Climate at Birth and the Risk of Obesity in Adult Life. Int. J. Obes. 2000, 24, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Yang, Y.; Wei, Q.; Zhang, Y.; Shi, H. Longitudinal Association of Maternal Pre-Pregnancy BMI and Third-Trimester Glycemia with Early Life Growth of Offspring: A Prospective Study among GDM-Negative Pregnant Women. Nutrients 2021, 13, 3971. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regression Model | Parameter Estimates | p Values | R2 | |
|---|---|---|---|---|
| A | ||||
| O males | Y = −134.07 + b1x for x ≤ XB | XB, 7.40 ± 0.35 | <0.01 | 0.99 |
| * Y = 6722.70 + b2x for x > XB | YB, 5444.32 ± 214.44 | <0.01 | ||
| b1, 753.47 ± 43.30 | <0.01 | |||
| * b2, −172.67 ± 87.68 | 0.07 | |||
| L males | Y = −64.66 + b1x for x ≤ XB | XB, 7.44 ± 0.17 | <0.01 | 1.00 |
| Y = YB for x > XB | YB, 4740.02 ± 88.98 | <0.01 | ||
| b1, 645.86 ± 15.77 | <0.01 | |||
| O females | Y = −64.66 + b1x for x ≤ XB | XB, 6.53 ± 0.49 | <0.01 | 1.00 |
| Y = YB for x > XB | YB, 3685.44 ± 199.75 | <0.01 | ||
| b1, 560.00 ± 46.43 | <0.01 | |||
| L females | Y = 29.76 + b1x for x ≤ XB | XB, 5.71 ± 0.51 | <0.01 | 0.99 |
| Y = YB for x > XB | YB, 3077.78 ± 173.91 | <0.01 | ||
| b1, 537.60 ± 55.17 | <0.01 | |||
| B | ||||
| OM males | Y = −140.98 + b1x for x ≤ XB | XB, 7.44 ± 0.18 | <0.01 | 1.00 |
| Y = 5979.85 + b2x for x > XB | YB, 5264.98 ± 104.83 | <0.01 | ||
| b1, 726.95 ± 19.68 | <0.01 | |||
| b2, −96.13 ± 40.34 | 0.03 | |||
| VM males | Y = −99.35 + b1x for x ≤ XB | XB, 7.40 ± 0.39 | <0.01 | 0.99 |
| Y = 6551.24 + b2x for x > XB | YB, 4955.41 ± 219.05 | <0.01 | ||
| b1, 682.91 ± 46.68 | <0.01 | |||
| b2, −215.60 ± 94.50 | 0.04 | |||
| LM males | Y = −56.36 + b1x for x ≤ XB | XB, 7.44 ± 0.17 | <0.01 | 1.00 |
| Y = YB for x > XB | YB, 4866.72 ± 87.88 | <0.01 | ||
| b1, 662.06 ± 15.55 | <0.01 | |||
| C | ||||
| OM females | Y = 11.71 + b1x for x ≤ XB | XB, 6.22 ± 0.49 | <0.01 | 1.00 |
| Y = YB for x > XB | YB, 3713.65 ± 227.96 | <0.01 | ||
| b1, 595.10 ± 50.65 | <0.01 | |||
| VM females | Y = 138.10 + b1x for x ≤ XB | XB, 7.30 ± 0.27 | <0.01 | 0.99 |
| Y = 4126.60 + b2x for x > XB | YB, 3483.33 ± 103.35 | <0.01 | ||
| b1, 458.15 ± 19.55 | <0.01 | |||
| b2, −88.10 ± 40.07 | 0.05 | |||
| LM females | Y = 9.52 + b1x for x ≤ XB | XB, 5.72 ± 0.54 | <0.01 | 1.00 |
| Y = 2590.93 + b2x for x > XB | YB, 3075.42 ± 182.50 | <0.01 | ||
| b1, 535.81 ± 56.21 | <0.01 | |||
| b2, 84.67 ± 39.93 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opsomer, H.; Liesegang, A.; Brugger, D.; Wichert, B. Growth Curves and Body Condition of Young Cats and Their Relation to Maternal Body Condition. Animals 2022, 12, 1373. https://doi.org/10.3390/ani12111373
Opsomer H, Liesegang A, Brugger D, Wichert B. Growth Curves and Body Condition of Young Cats and Their Relation to Maternal Body Condition. Animals. 2022; 12(11):1373. https://doi.org/10.3390/ani12111373
Chicago/Turabian StyleOpsomer, Han, Annette Liesegang, Daniel Brugger, and Brigitta Wichert. 2022. "Growth Curves and Body Condition of Young Cats and Their Relation to Maternal Body Condition" Animals 12, no. 11: 1373. https://doi.org/10.3390/ani12111373