
Prevalence, Antimicrobial Resistance, and Characterization of Staphylococcus aureus Isolated from Subclinical Bovine Mastitis in East Coast Malaysia

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
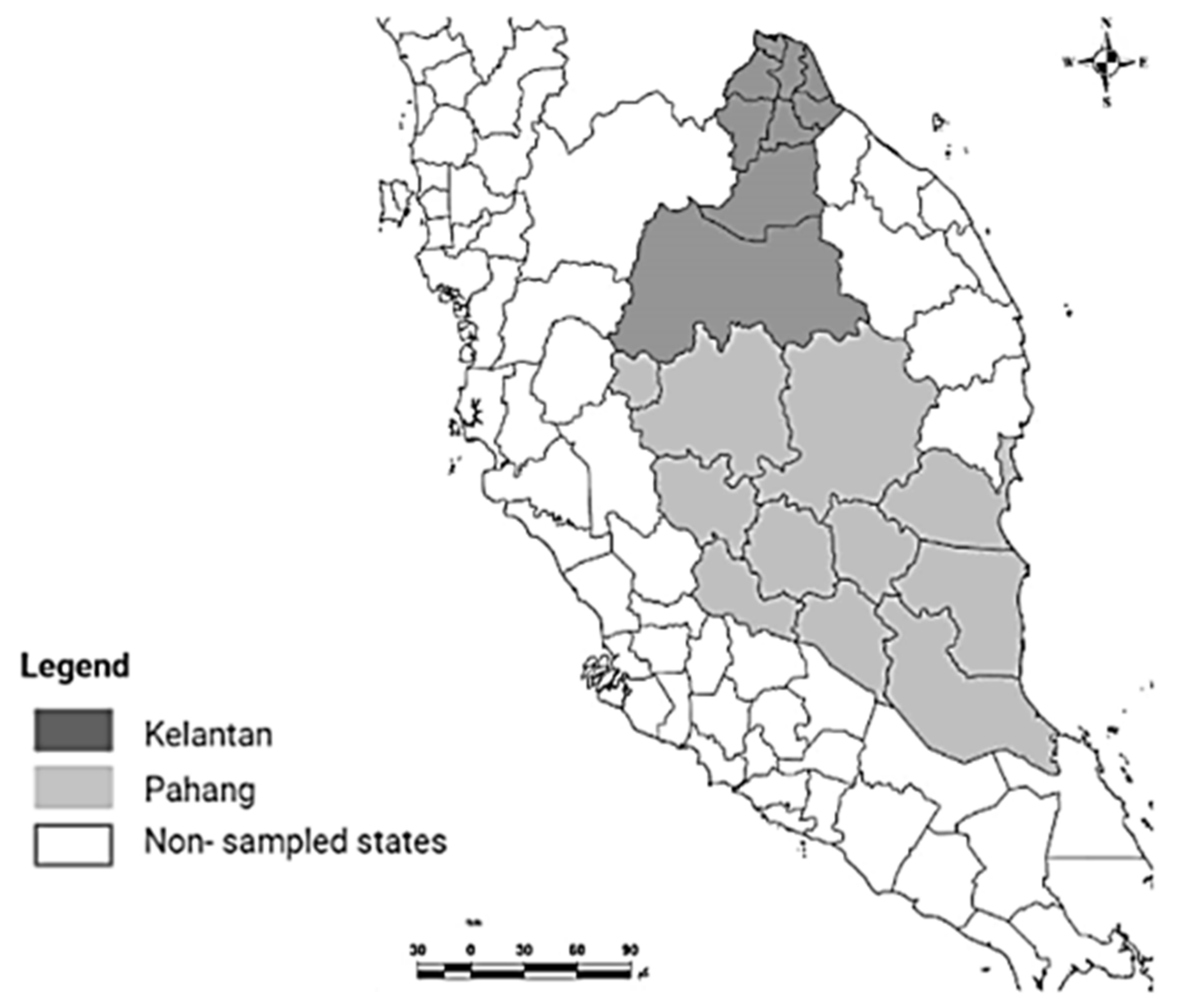
2.1. Study Area and Sampling
2.2. Determining the Somatic Cell Count (SCC)
2.3. Isolation and Identification of S. aureus
2.4. Antimicrobial Susceptibility Tests
2.5. Mammary Epithelial Cell Cultures (MAC-T Cells) and Growth Condition
2.6. Intracellular Invasions of MAC-T Cells by S. aureus
2.7. Biofilm Formation
2.8. Statistical Analysis
3. Results
3.1. Prevalance of Subclinical Mastitis and S. aureus
3.2. Antimicrobial Susceptibility Testing
3.3. S. aureus AMR Profile and Multiple Antimicrobial Resistance (MAR) Indexes
3.4. S. aureus Biofilm Formation and Their Association with AMR Profile
3.5. Intracellular Invasion of MAC-T Cell by S. aureus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Li, H.; Pan, J.; Dong, J.; Zhou, X.; Niu, X.; Deng, X. Oligopeptide targeting sortase a as potential anti-infective therapy for Staphylococcus aureus. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Zhou, K.; Li, C.; Chen, D.; Pan, Y.; Tao, Y.; Qu, W.; Liu, Z.; Wang, X.; Xie, S. A review on nanosystems as an effective approach against infections of Staphylococcus aureus. Int. J. Nanomed. 2018, 13, 7333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunert, T.; Stessl, B.; Wolf, F.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M. Distinct phenotypic traits of Staphylococcus aureus are associated with persistent, contagious bovine intramammary infections. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdi, R.D.; Gillespie, B.E.; Ivey, S.; Pighetti, G.M.; Almeida, R.A.; Dego, O.K. Antimicrobial resistance of major bacterial pathogens from dairy cows with high somatic cell count and clinical mastitis. Animals 2021, 11, 131. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.; Henriques, M. Control of Bovine Mastitis: Old and Recent Therapeutic Approaches. Curr. Microbiol. 2016, 72, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Molineri, A.I.; Camussone, C.; Zbrun, M.V.; Suárez Archilla, G.; Cristiani, M.; Neder, V.; Calvinho, L.; Signorini, M. Antimicrobial resistance of Staphylococcus aureus isolated from bovine mastitis: Systematic review and meta-analysis. Prev. Vet. Med. 2021, 188, 105261. [Google Scholar] [CrossRef]
- Jamali, H.; Paydar, M.; Radmehr, B.; Ismail, S.; Dadrasnia, A. Prevalence and antimicrobial resistance of Staphylococcus aureus isolated from raw milk and dairy products. Food Control. 2015, 54, 383–388. [Google Scholar] [CrossRef]
- Varela-Ortiz, D.F.; Barboza-Corona, J.E.; González-Marrero, J.; León-Galván, M.F.; Valencia-Posadas, M.; Lechuga-Arana, A.A.; Sánchez-Felipe, C.G.; Ledezma-García, F.; Gutiérrez-Chávez, A.J. Antibiotic susceptibility of Staphylococcus aureus isolated from subclinical bovine mastitis cases and in vitro efficacy of bacteriophage. Vet. Res. Commun. 2018, 42, 243–250. [Google Scholar] [CrossRef]
- Sasidharan, S.; Prema, B.; Latha, L.Y. Antimicrobial drug resistance of Staphylococcus aureus in dairy products. Asian Pac. J. Trop. Biomed. 2011, 1, 130–132. [Google Scholar] [CrossRef] [Green Version]
- León-Galván, M.F.; Barboza-Corona, J.E.; Lechuga-Arana, A.A.; Valencia-Posadas, M.; Aguayo, D.D.; Cedillo-Pelaez, C.; Martínez-Ortega, E.A.; Gutierrez-Chavez, A.J. Molecular detection and sensitivity to antibiotics and bacteriocins of pathogens isolated from bovine mastitis in family dairy herds of central mexico. Biomed. Res. Int. 2015, 2015, 9. [Google Scholar] [CrossRef]
- Dai, J.; Wu, S.; Huang, J.; Wu, Q.; Zhang, F.; Zhang, J.; Wang, J.; Ding, Y.; Zhang, S.; Yang, X.; et al. Prevalence and Characterization of Staphylococcus aureus Isolated from Pasteurized Milk in China. Front. Microbiol. 2019, 10, 641. [Google Scholar] [CrossRef] [PubMed]
- Clement, S.; Vaudaux, P.; Francois, P.; Schrenzel, J.; Huggler, E.; Kampf, S.; Chaponnier, C.; Lew, D.; Lacroix, J.S. Evidence of an intracellular reservoir in the nasal mucosa of patients with recurrent Staphylococcus aureus rhinosinusitis. J. Infect. Dis. 2005, 192, 1023–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monack, D.M.; Mueller, A.; Falkow, S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat. Rev. Microbiol. 2004, 2, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Mayowa, O.O.; Pour, S.H.; Shahid, S.; Mohsenipour, M.; Harun, S.B.; Heryansyah, A.; Ismail, T. Trends in rainfall and rainfall-related extremes in the east coast of peninsular Malaysia. J. Earth Syst. Sci. 2015, 124, 1609–1622. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.P.; Gonzalez, R.N.; Hogan, J.S.; Jayarao, B.M.; Owens, W.E. Microbiological procedures for the diagnosis of bovine udder infection and determination of milk quality. Verona Natl. Mastit. Counc. 2004, 47. [Google Scholar]
- Damm, M.; Holm, C.; Blaabjerg, M.; Bro, M.N.; Schwarz, D. Differential somatic cell count—A novel method for routine mastitis screening in the frame of Dairy Herd Improvement testing programs. J. Dairy Sci. 2017, 100, 4926–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wayne, P.A. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing. Inf. Suppl. 2021, 31, 100–121. [Google Scholar]
- Davis, R.; Brown, P.D. Multiple antibiotic resistance index, fitness and virulence potential in respiratory Pseudomonas aeruginosa from Jamaica. J. Med. Microbiol. 2016, 65, 261–271. [Google Scholar] [CrossRef]
- Kamaruzzaman, N.F.; Chong, S.Q.; Edmondson-Brown, K.M.; Ntow-Boahene, W.; Bardiau, M.; Good, L. Bactericidal and anti-biofilm effects of polyhexamethylene Biguanide in models of intracellular and biofilm of Staphylococcus aureus isolated from bovine mastitis. Front. Microbiol. 2017, 11, 1518. [Google Scholar] [CrossRef]
- Kamal, R.M.; Bayoumi, M.A.; Abd El Aal, S.F. Correlation between some direct and indirect tests for screen detection of subclinical mastitis. Int. Food Res. J. 2014, 1, 1249–1254. [Google Scholar]
- Sharma, N.; Pandey, V.; Sudhan, N.A. Comparison of some indirect screening tests for detection of subclinical mastitis in dairy cows. Bulg. J. Vet. Med. 2010, 1, 13. [Google Scholar]
- Marimuthu, M.; Abdullah, F.F.J.; Mohammed, K.; Poshpum, S.S.; Adamu, L.; Osman, A.Y.; Abba, Y.; Tijjani, A. Prevalence and antimicrobial resistance assessment of subclinical mastitis in milk samples from selected dairy farms. Am. J. Anim. Vet. Sci. 2014, 9, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Othman, N.; Bahaman, A.R. Prevalence of Subclinical Mastitis and Antibiotic Resistant Bacteria in Three. Malaysian J. Vet. Res. 2005, 17, 27–31. [Google Scholar]
- Liu, H.; Li, S.; Meng, L.; Dong, L.; Zhao, S.; Lan, X.; Wang, J.; Zheng, N. Prevalence, antimicrobial susceptibility, and molecular characterization of Staphylococcus aureus isolated from dairy herds in northern China. J. Dairy Sci. 2017, 100, 8796–8803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rall, V.L.M.; Miranda, E.S.; Castilho, I.G.; Camargo, C.H.; Langoni, H.; Guimarães, F.F.; Júnior, J.A.; Júnior, A.F. Diversity of Staphylococcus species and prevalence of enterotoxin genes isolated from milk of healthy cows and cows with subclinical mastitis. J. Dairy Sci. 2014, 97, 829–837. [Google Scholar] [CrossRef]
- Murat, K.; Mehmet, N.; Burhan, C. Investigation of Toxin Genes by Polymerase Chain Reaction in Staphylococcus aureus Strains Isolated from Bovine Mastitis in Turkey. Foodborne Pathog. Dis. 2009, 6, 1029–1035. [Google Scholar]
- Wang, D.; Jia, D.; He, R.; Lian, S.; Wang, J.; Wu, R. Association between serum selenium level and subclinical mastitis in dairy cattle. Biol. Trace Elem. Res. 2021, 199, 1389–1396. [Google Scholar] [CrossRef]
- El-Sayed, A.; Kamel, M. Bovine mastitis prevention and control in the post-antibiotic era. Trop. Anim. Health Prod. 2021, 53, 236. [Google Scholar] [CrossRef]
- Chai, M.H.; Faiq, T.A.; Ariffin, S.M.; Suhaili, Z.; Sukiman, M.Z.; Ghazali, M.F. Prevalence of methicillin resistant Staphylococcus aureus in raw goat milks from selected farms in Terengganu, Malaysia. Trop. Anim. Sci. J. 2020, 43, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Elemo, K.K.; Sisay, T.; Shiferaw, A.; Fato, M.A. Prevalence, risk factors and multidrug resistance profile of Staphylococcus aureus isolated from bovine mastitis in selected dairy farms in and around Asella town, Arsi Zone, South Eastern Ethiopia. Afr. J. Microbiol. Res. 2017, 11, 1632–1642. [Google Scholar]
- Anaya-López, J.L.; Contreras-Guzmán, O.E.; Cárabez-Trejo, A.; Baizabal-Aguirre, V.M.; López-Meza, J.E.; Valdez-Alarcón, J.J.; Ochoa-Zarzosa, A. Invasive potential of bacterial isolates associated with subclinical bovine mastitis. Res. Vet. Sci. 2006, 81, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Fraunholz, M.; Sinha, B. Intracellular Staphylococcus aureus: Live-in and let die. Front. Cell. Infect. Microbiol. 2012, 24, 43. [Google Scholar] [CrossRef] [Green Version]
- Günther, J.; Petzl, W.; Bauer, I.; Ponsuksili, S.; Zerbe, H.; Schuberth, H.J.; Brunner, R.M.; Seyfert, H.M. Differentiating Staphylococcus aureus from Escherichia coli mastitis: S. aureus triggers unbalanced immune-dampening and host cell invasion immediately after udder infection. Sci. Rep. 2017, 6, 4811. [Google Scholar] [CrossRef] [PubMed]
- Notcovich, S.; DeNicolo, G.; Flint, S.H.; Williamson, N.B.; Gedye, K.; Grinberg, A.; Lopez-Villalobos, N. Biofilm-forming potential of Staphylococcus aureus isolated from bovine mastitis in New Zealand. Vet. Sci. 2018, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, F.; Saavedra, M.J.; Henriques, M. Bovine mastitis disease/pathogenicity: Evidence of the potential role of microbial biofilms. Pathog Dis. 2016, 74, 74. [Google Scholar] [CrossRef] [Green Version]
- Fox, L.K.; Zadoks, R.N.; Gaskins, C.T. Biofilm production by Staphylococcus aureus associated with intramammary infection. Vet. Microbiol. 2005, 107, 295–299. [Google Scholar] [CrossRef]
- Kamaruzzaman, N.F.; Tan, L.P.; Yazid, K.A.M.; Saeed, S.I.; Hamdan, R.H.; Choong, S.S.; Wong, W.K.; Chivu, A.; Gibson, A.J. Targeting the Bacterial protective armour; challenges and novel strategies in the treatment of microbial biofilm. Materials 2018, 11, 1705. [Google Scholar] [CrossRef] [Green Version]
- Ranieri, M.R.; Whitchurch, C.B.; Burrows, L.L. Mechanisms of biofilm stimulation by subinhibitory concentrations of antimicrobials. Curr. Opin. Opin. Opin. Microbiol. 2018, 1, 164–169. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| State/locality | No. of samples | Subclinical Mastitis (%) | S. aureus isolates (%) |
|---|---|---|---|
| Kelantan | |||
| Pasir Putih | 37 | 10 (27%) | 14 (37.8%) |
| Kota Bharu | 12 | 2 (16.6%) | 1(8.3%) |
| Pasir Mas | 11 | 1 (9.0%) | 3 (27%) |
| Total | 60 | 13 (21.6%) | 18 (30%) |
| Pahang | |||
| Bentong | 84 | 38 (45.2%) | 13 (15.5%) |
| Raub | 91 | 23 (25.2%) | 8 (8.7%) |
| Total | 175 | 61 (34.8) | 21 (12%) |
| Overall | 235 | 74 (31.4%) | 39 (16.5%) |
| Antimicrobials | Dose (µg) | Breakpoint | |||||
|---|---|---|---|---|---|---|---|
| S (mm) | R (mm) | * n/39 | R (%) | I (%) | S (%) | ||
| P | 10 | ≥29 | ≤28 | 18 | 46 | 0 | 54 |
| AMP | 10 | ≥29 | ≤28 | 17 | 43.6 | 0 | 56.4 |
| CN | 10 | ≥23 | ≤17 | 0 | 0 | 0 | 100 |
| E | 15 | ≥23 | ≤13 | 7 | 18 | 0 | 82 |
| OX | 1 | ≥13 | ≤10 | 12 | 31 | 0 | 69 |
| SXT | 25 | ≥16 | ≤10 | 0 | 0 | 0 | 100 |
| TE | 30 | ≥23 | ≤17 | 10 | 26 | 0 | 74 |
| CIP | 5 | ≥21 | ≤15 | 0 | 0 | 0 | 100 |
| C | 30 | ≥18 | ≤12 | 0 | 0 | 0 | 100 |
| S. aureus Isolates Code | AMR Profile | No. of Antimicrobial Class | MAR Index |
|---|---|---|---|
| F51B | AMP, P | 1 | 0.11 |
| F02B | AMP, P | 1 | 0.11 |
| F30B | AMP, P | 1 | 0.11 |
| F32A | AMP, P | 1 | 0.11 |
| PBF14C | AMP, P | 1 | 0.11 |
| F31A | AMP, P, OXA | 1 | 0.11 |
| PBF38C | AMP, P, OXA | 1 | 0.11 |
| F04B | AMP, P, TE, OXA | 2 | 0.22 |
| PBF39C | AMP, P, TE, OXA | 2 | 0.22 |
| PBF18A | AMP, P, TE, OXA | 2 | 0.22 |
| F31D | AMP, E, P, TE, OXA | 3 | 0.33 |
| F32C | AMP, E, P, TE, OXA | 3 | 0.33 |
| F32B | AMP, E, P, TE, OXA | 3 | 0.33 |
| F41A | AMP, E, P, TE, OXA | 3 | 0.33 |
| F41B | AMP, E, P, TE, OXA | 3 | 0.33 |
| PBF2IB | AMP, E, P, TE, OXA | 3 | 0.33 |
| PBF33C | AMP, E, P, TE, OXA | 3 | 0.33 |
| NO | S. aureus Isolates | Biofilm Production Assay | Biofilm Production Ability | Antimicrobial Susceptibility | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P | AMP | CN | E | OX | SXT | TE | CIP | C | ||||
| 1. | F04B | 0.17 | + | R | R | S | S | R | S | R | S | S |
| 2. | PBF38C | 0.2 | + | R | R | S | S | R | S | S | S | S |
| 3. | F31D | 0.4 | + | R | R | S | R | R | S | R | S | S |
| 4. | PBF18A | 0.4 | + | R | R | S | S | R | S | R | S | S |
| 5. | F51B | 0.82 | ++ | R | R | S | S | S | S | S | S | S |
| 6. | F11A | 0.94 | ++ | S | S | S | S | S | S | S | S | S |
| 7. | F10A | 0.94 | ++ | S | S | S | S | S | S | S | S | S |
| 8. | F31A | 1.0 | ++ | R | R | S | R | S | S | S | S | S |
| 9. | F13C | 1.1 | ++ | S | S | S | S | S | S | S | S | S |
| 10. | PBF39C | 1.15 | ++ | R | R | S | S | R | S | R | S | S |
| 11. | F10B | 1.4 | ++ | S | S | S | S | S | S | S | S | S |
| 12. | PBF14C | 1.4 | ++ | R | R | S | S | S | S | S | S | S |
| 13. | F32B | 1.6 | ++ | R | R | S | R | R | S | R | S | S |
| 14. | F32C | 1.6 | ++ | R | R | S | R | R | S | R | S | S |
| 15. | F41A | 1.8 | ++ | R | R | S | R | R | S | R | S | S |
| 16. | F13A | 1.8 | ++ | S | S | S | S | S | S | S | S | S |
| 17. | F02B | 2.0 | ++ | R | R | S | S | S | S | S | S | S |
| 18. | F30B | 2.0 | ++ | R | R | S | S | S | S | S | S | S |
| 19. | F32A | 2.0 | ++ | R | R | S | S | S | S | S | S | S |
| 20. | PBF21B | 2.0 | ++ | R | R | S | R | R | S | R | S | S |
| 21. | PBF13B | 2.0 | ++ | S | S | S | S | S | S | S | S | S |
| 22. | PBF22B | 2.0 | ++ | S | S | S | S | S | S | S | S | S |
| 23. | PBF33C | 2.1 | +++ | R | R | S | R | R | S | R | S | S |
| 24. | F11C | 2.2 | +++ | S | S | S | S | S | S | S | S | S |
| 25. | F11C | 2.2 | +++ | S | S | S | S | S | S | S | S | S |
| 26. | F31C | 2.3 | +++ | S | S | S | S | S | S | S | S | S |
| 27. | PBF39A | 2.4 | +++ | S | S | S | S | S | S | S | S | S |
| 28. | F41B | 2.4 | +++ | R | R | S | R | R | S | R | S | S |
| 29. | F03B | 2.5 | +++ | S | S | S | S | S | S | S | S | S |
| 30. | F53D | 2.5 | +++ | S | S | S | S | S | S | S | S | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, S.I.; Mat Yazid, K.A.; Hashimy, H.A.; Dzulkifli, S.K.; Nordin, F.; Nik Him, N.A.; Omar, M.F.F.b.; Aklilu, E.; Mohamad, M.; Zalati, C.W.S.; et al. Prevalence, Antimicrobial Resistance, and Characterization of Staphylococcus aureus Isolated from Subclinical Bovine Mastitis in East Coast Malaysia. Animals 2022, 12, 1680. https://doi.org/10.3390/ani12131680
Saeed SI, Mat Yazid KA, Hashimy HA, Dzulkifli SK, Nordin F, Nik Him NA, Omar MFFb, Aklilu E, Mohamad M, Zalati CWS, et al. Prevalence, Antimicrobial Resistance, and Characterization of Staphylococcus aureus Isolated from Subclinical Bovine Mastitis in East Coast Malaysia. Animals. 2022; 12(13):1680. https://doi.org/10.3390/ani12131680
Chicago/Turabian StyleSaeed, Shamsaldeen Ibrahim, Khairun Anisa Mat Yazid, Hidayatul Athirah Hashimy, Siti Khadijah Dzulkifli, Fatihah Nordin, Nik Azmi Nik Him, Mohd Fikry Fahmi bin Omar, Erkihun Aklilu, Maizan Mohamad, Che Wan Salma Zalati, and et al. 2022. "Prevalence, Antimicrobial Resistance, and Characterization of Staphylococcus aureus Isolated from Subclinical Bovine Mastitis in East Coast Malaysia" Animals 12, no. 13: 1680. https://doi.org/10.3390/ani12131680