
Proteomics Insights into the Gene Network of cis9, trans11-Conjugated Linoleic Acid Biosynthesis in Bovine Mammary Gland Epithelial Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Treatment and Real-Time PCR Analysis
2.3. Fatty Acids Analysis
2.4. Identification of Differentially Expressed Proteins by Tandem Mass Tag (TMT)-Based Quantitative Proteomics
2.5. Protein Functional Annotation and Bioinformatic Analysis
2.6. Parallel Reaction Monitoring (PRM) Validation
2.7. Small RNA Interference
2.8. Statistical Analysis
3. Results
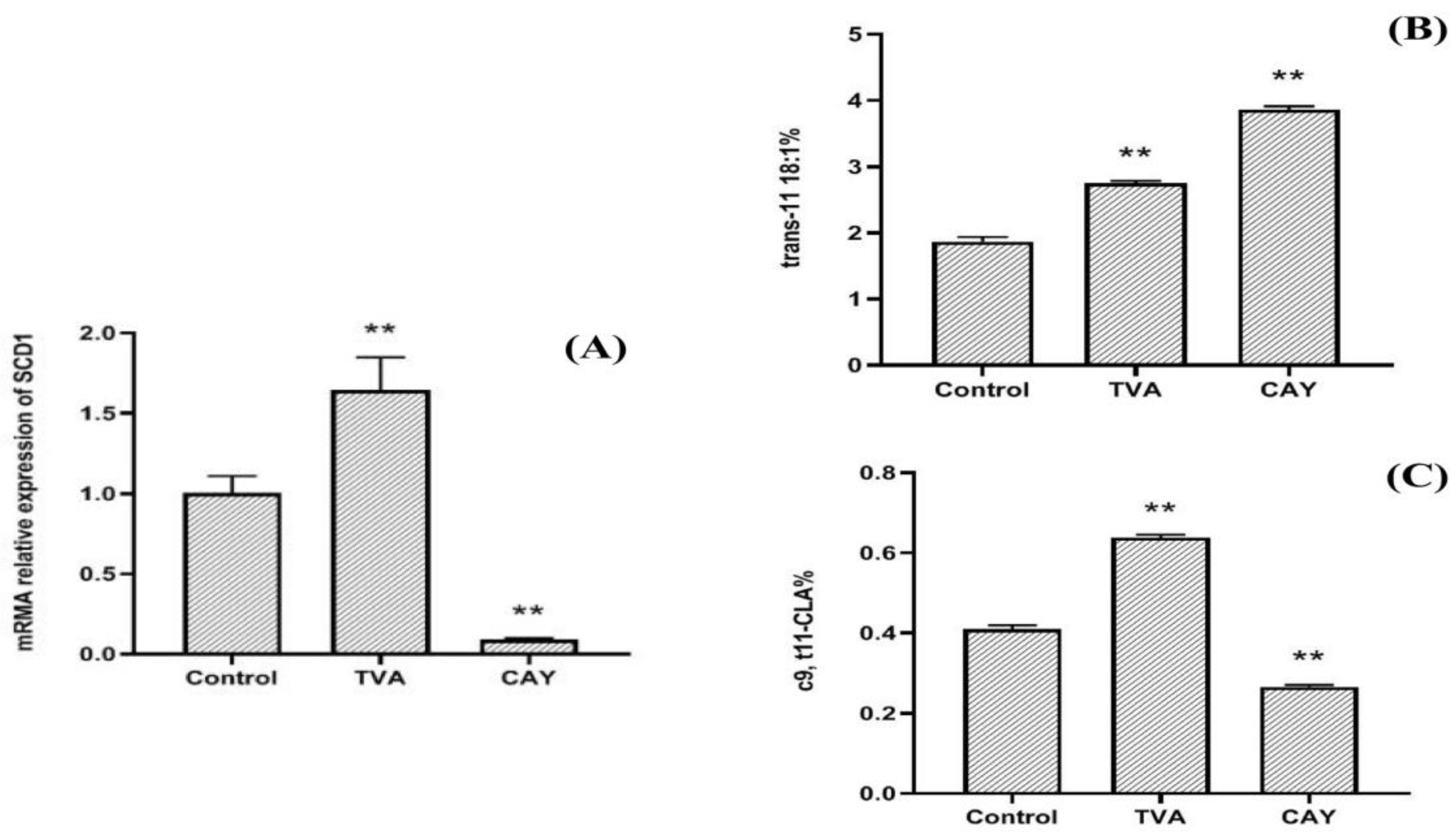
3.1. Effects of CAY and TVA on SCD1 mRNA Expression
3.2. Effects of CAY and TVA on TVA Accumulation and c9, t11-CLA Synthesis
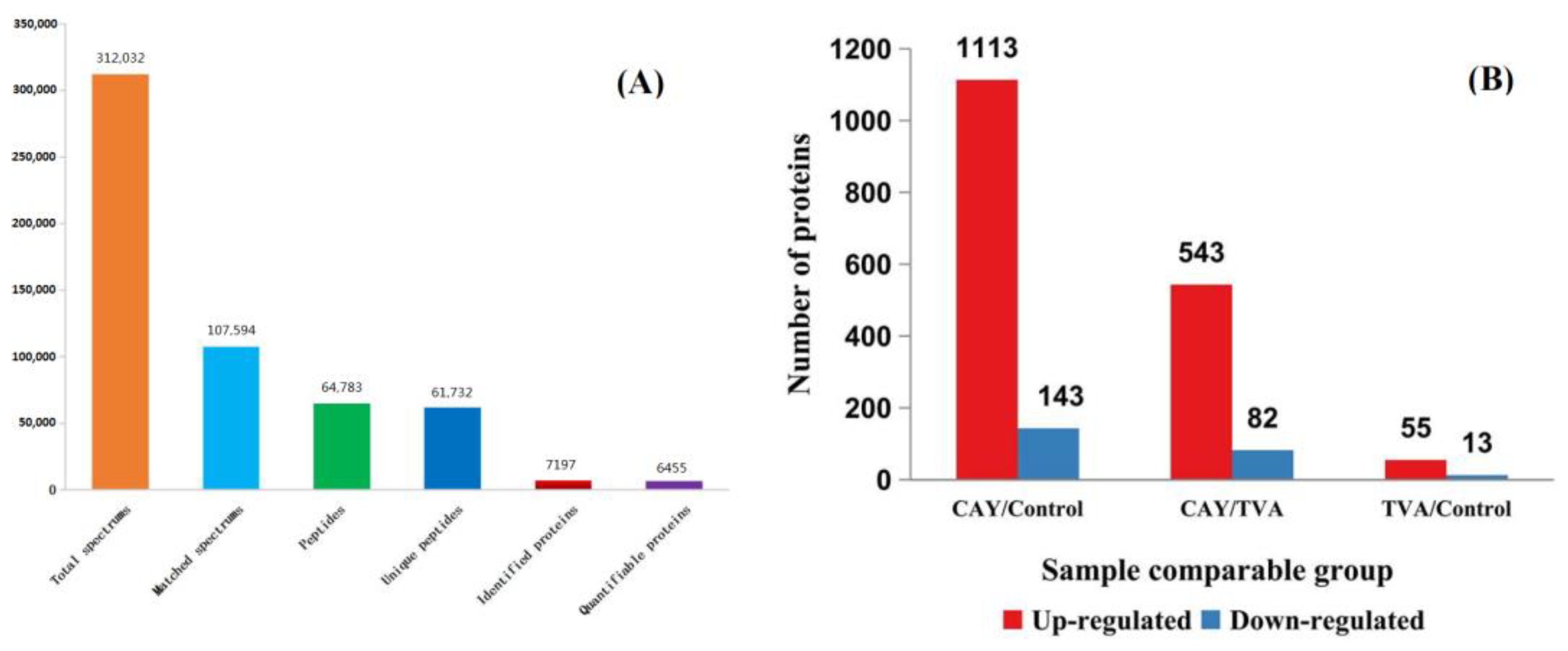
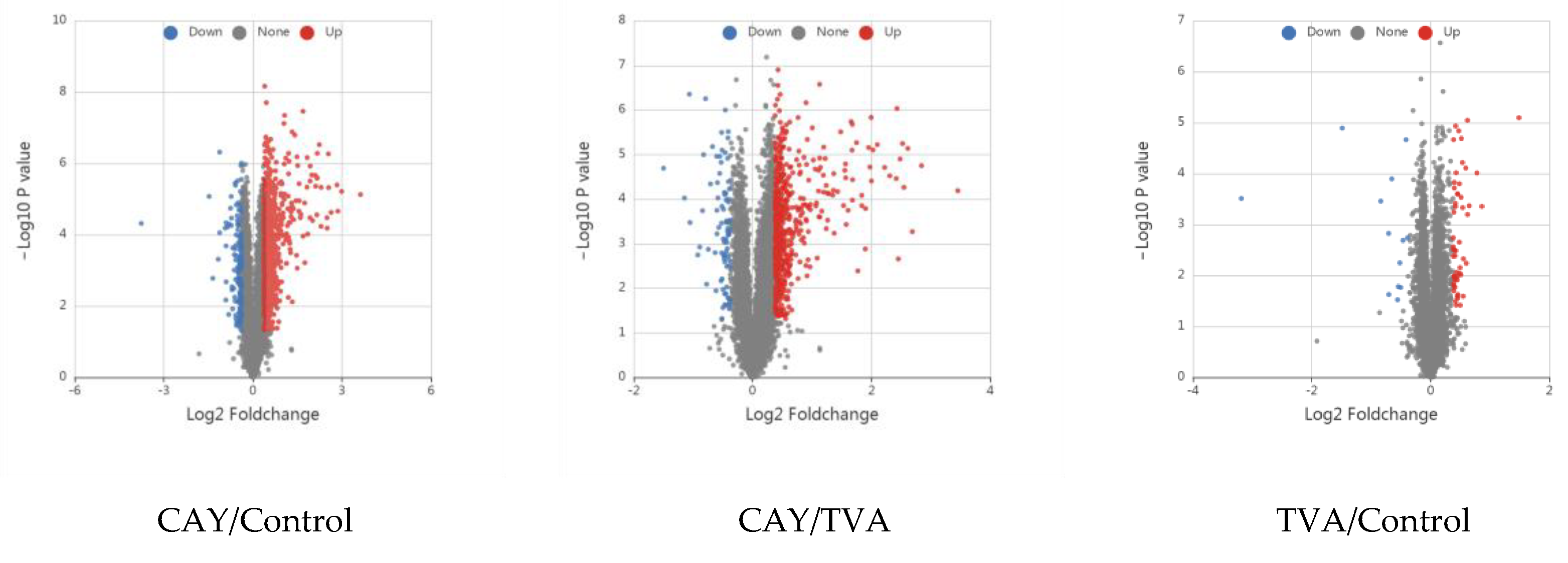
3.3. Identification of Differentially Expressed Proteins
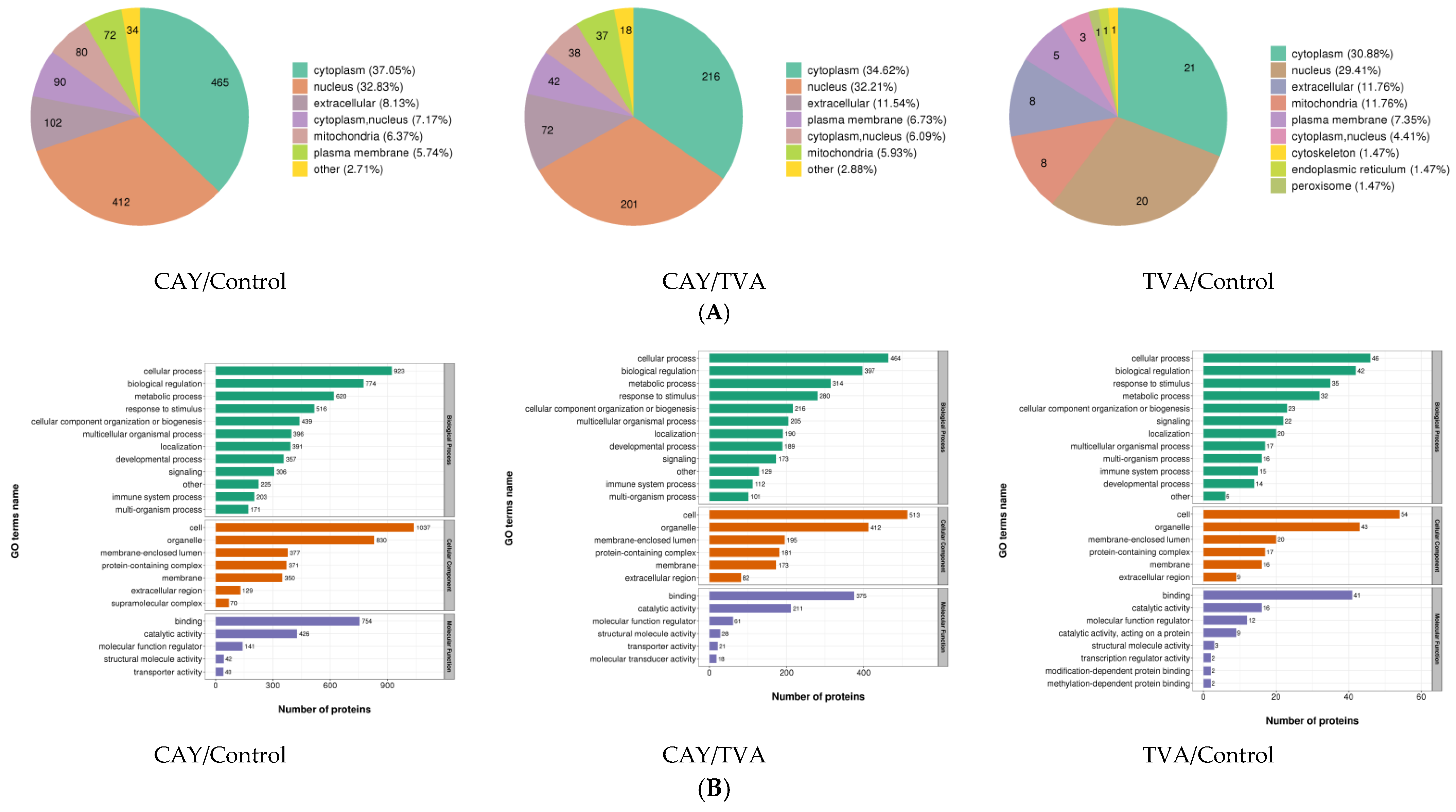
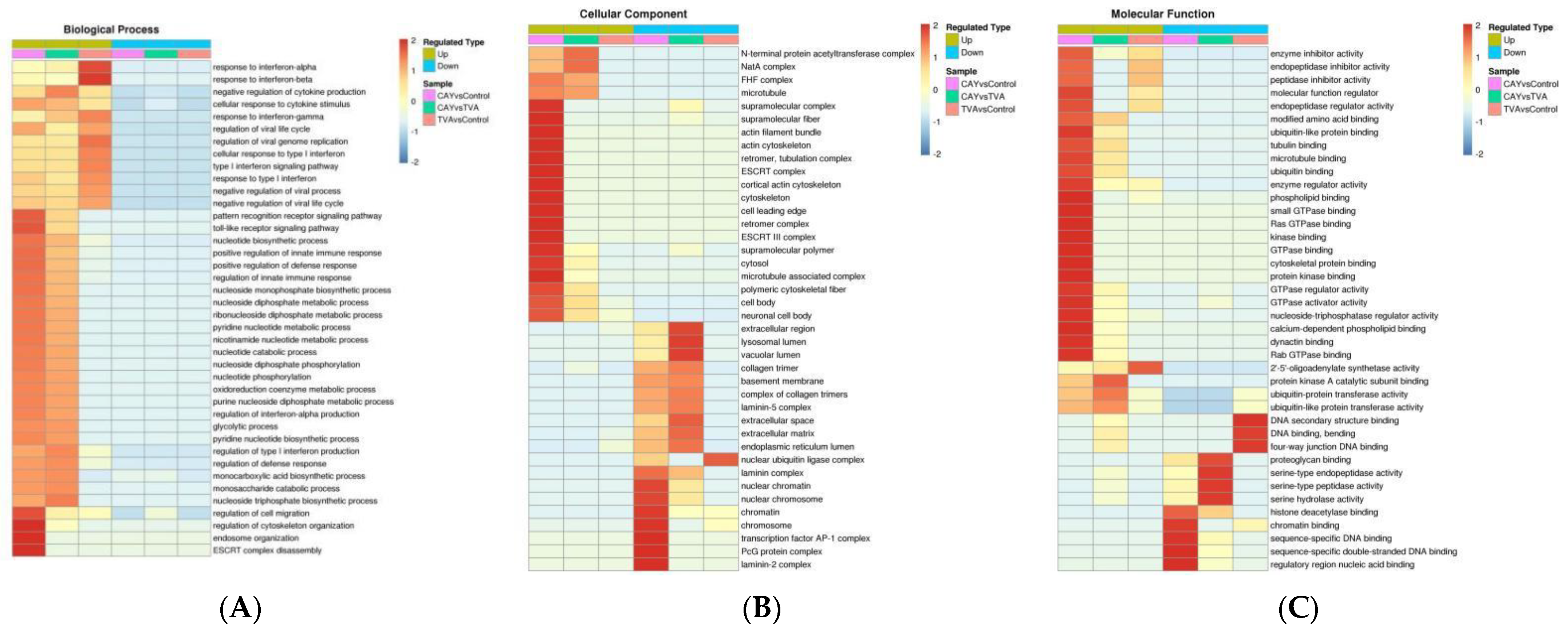
3.4. Functional Annotation of Differentially Expressed Proteins
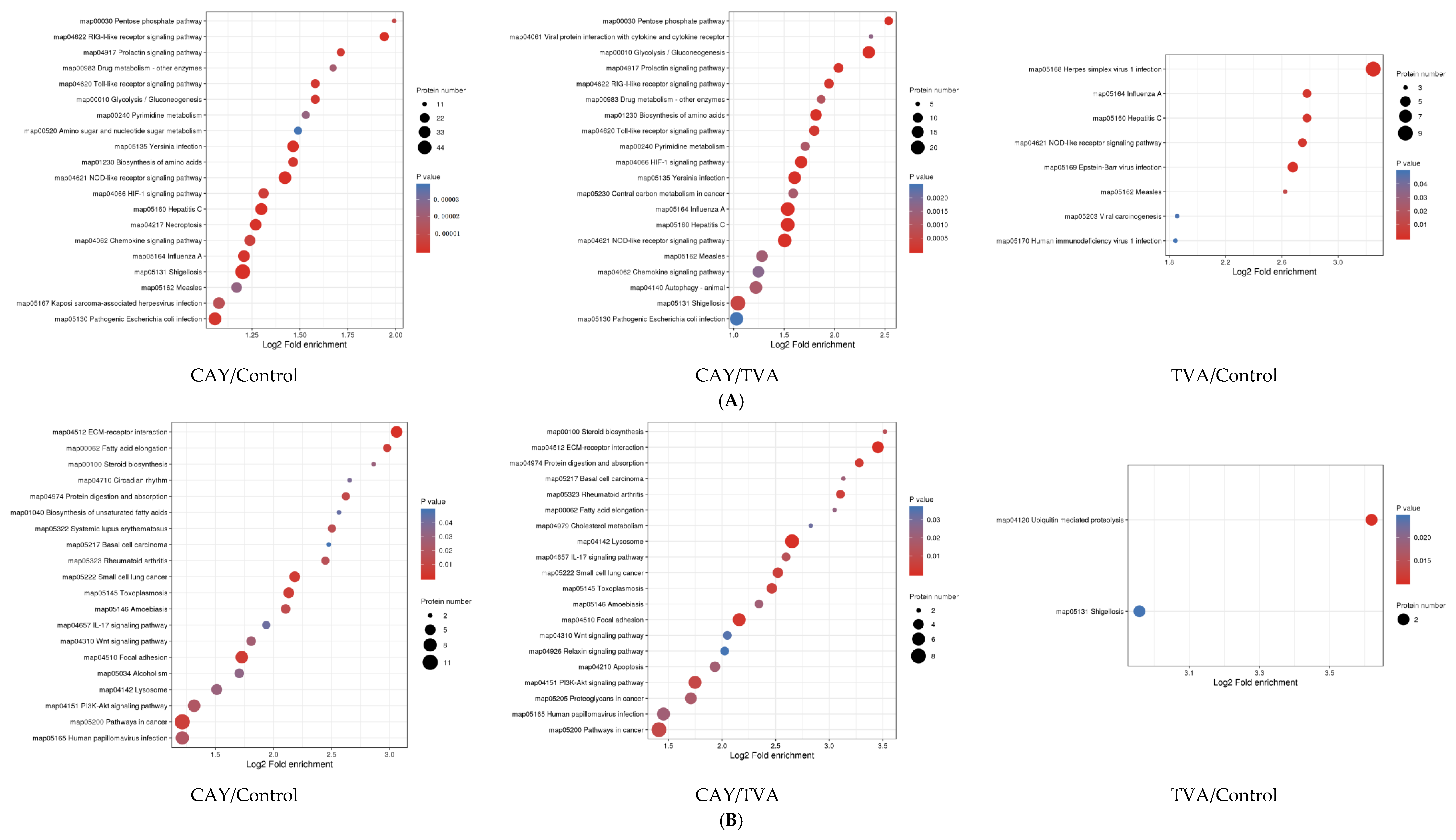
3.5. GO and KEGG Analysis of Differentially Expressed Proteins
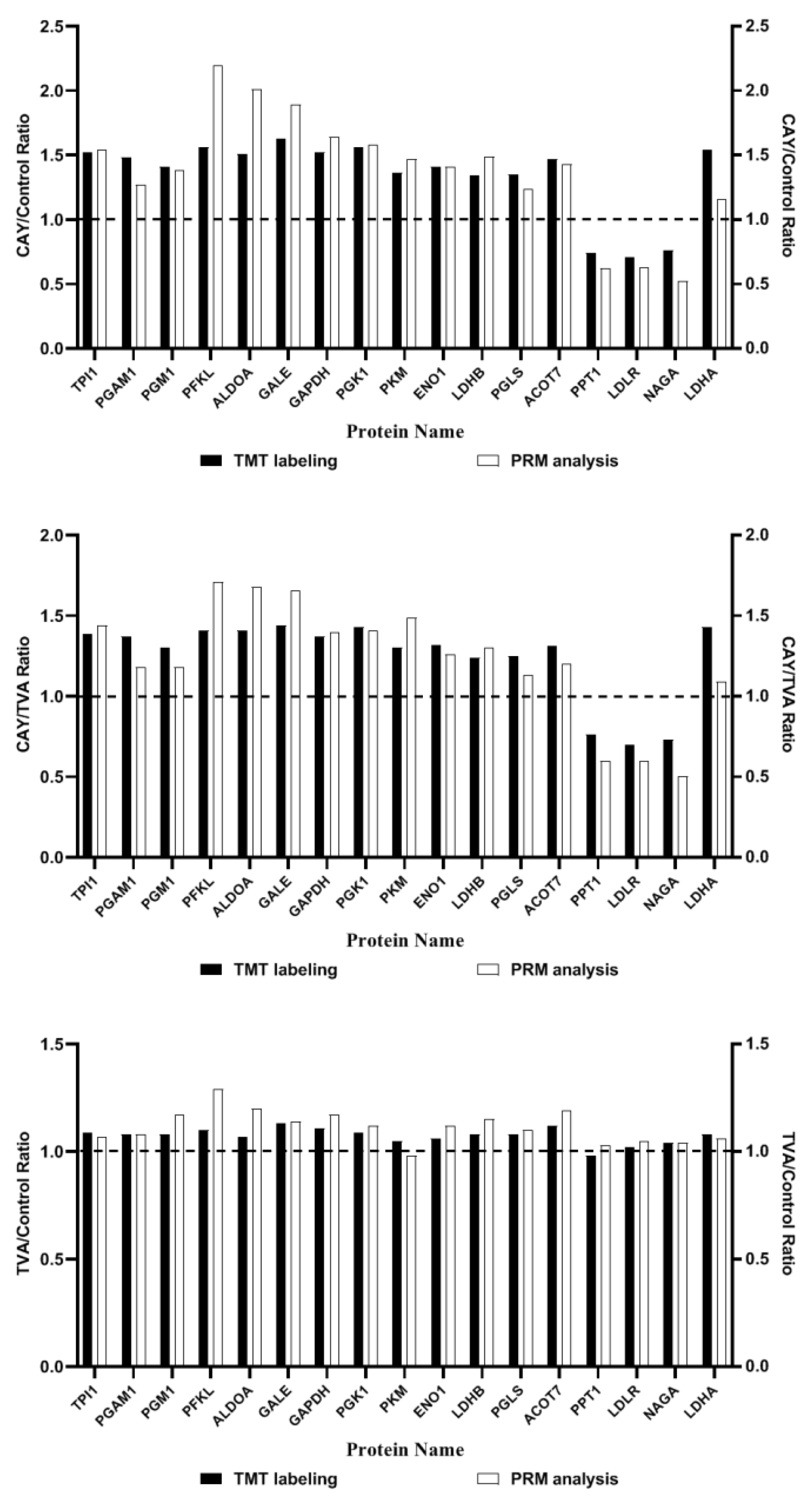
3.6. PRM Validation
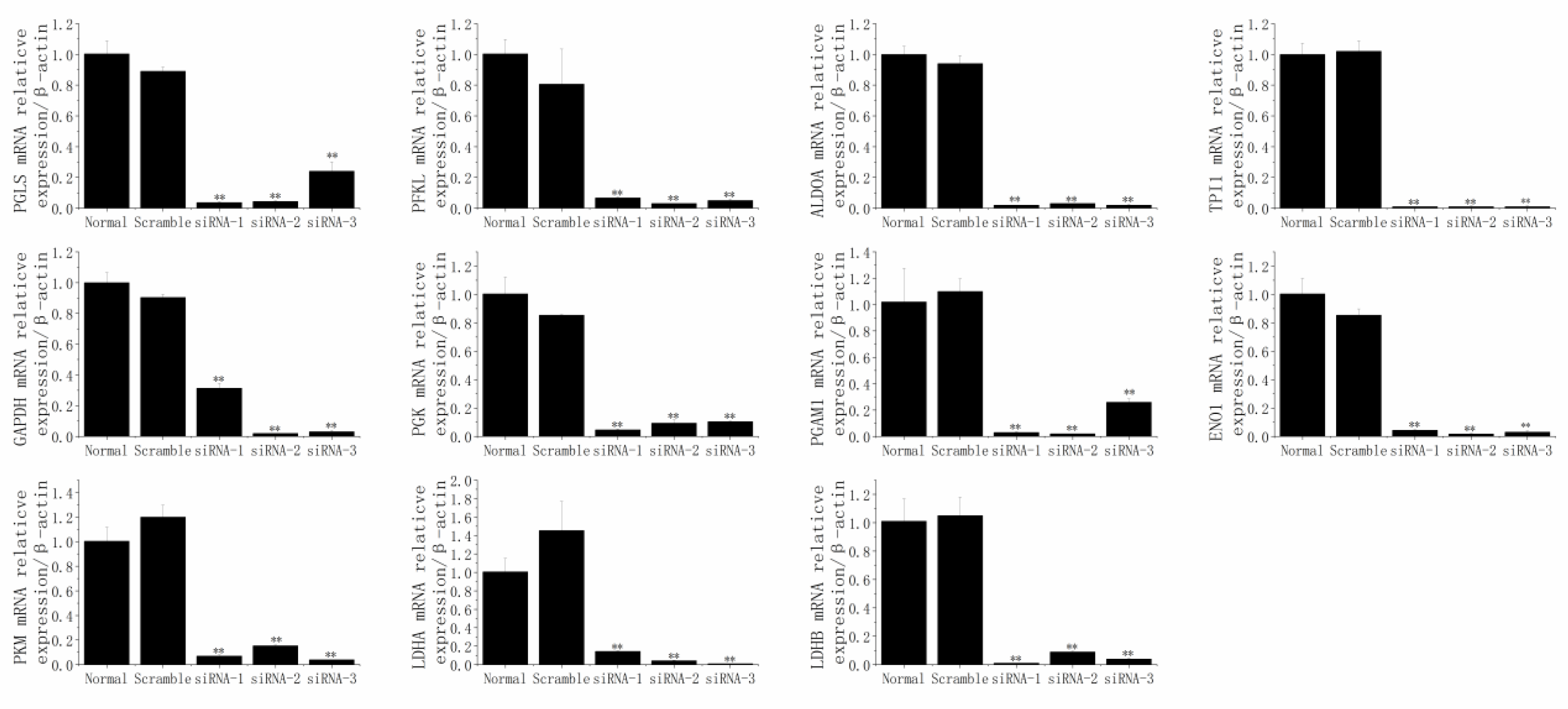
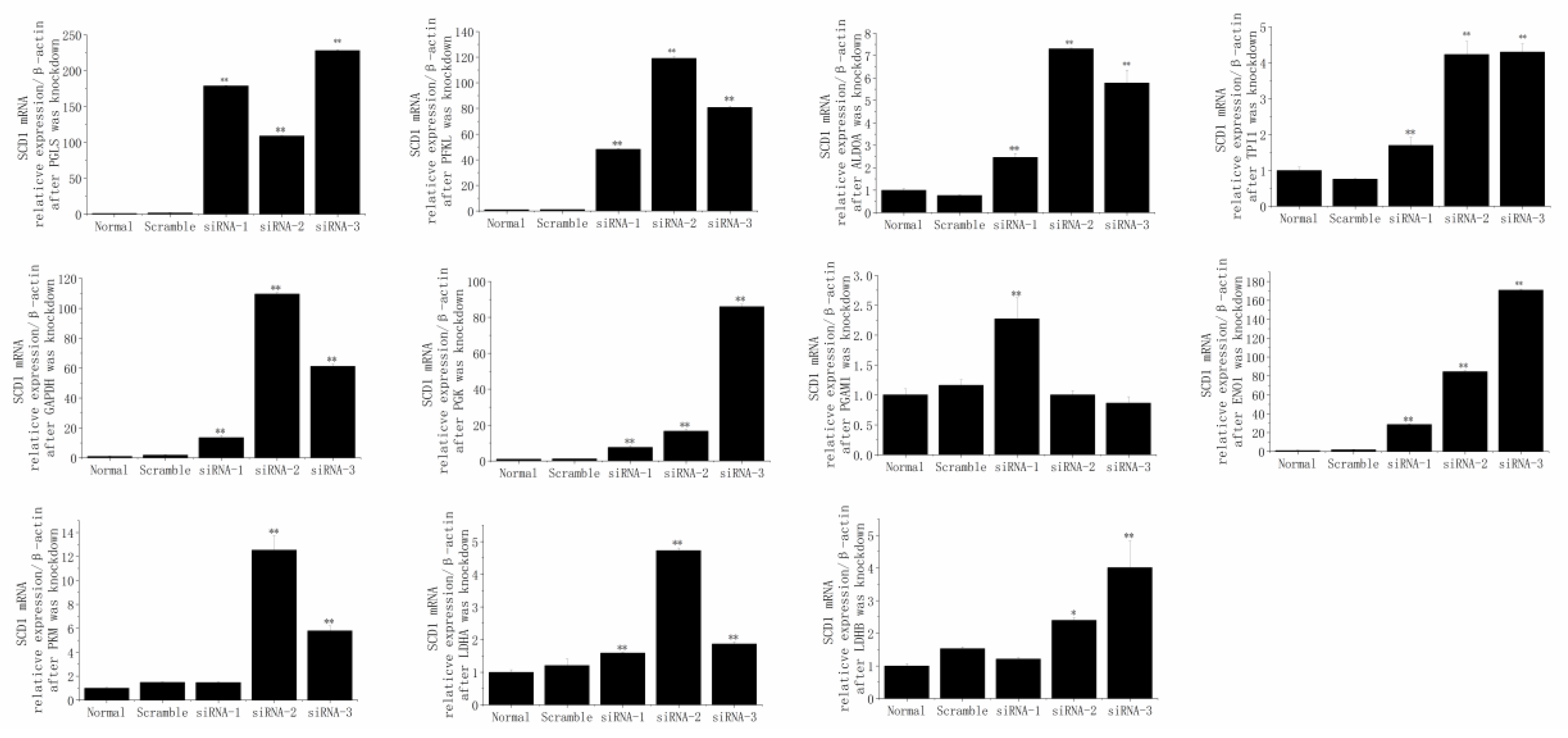
3.7. The Responses of SCD1 after the Knockdown of PGLS, PFKL, ALDOA, TPI1, GAPDH, PGK1, PGAM1, ENO1, PKM, LDHA, and LDHB
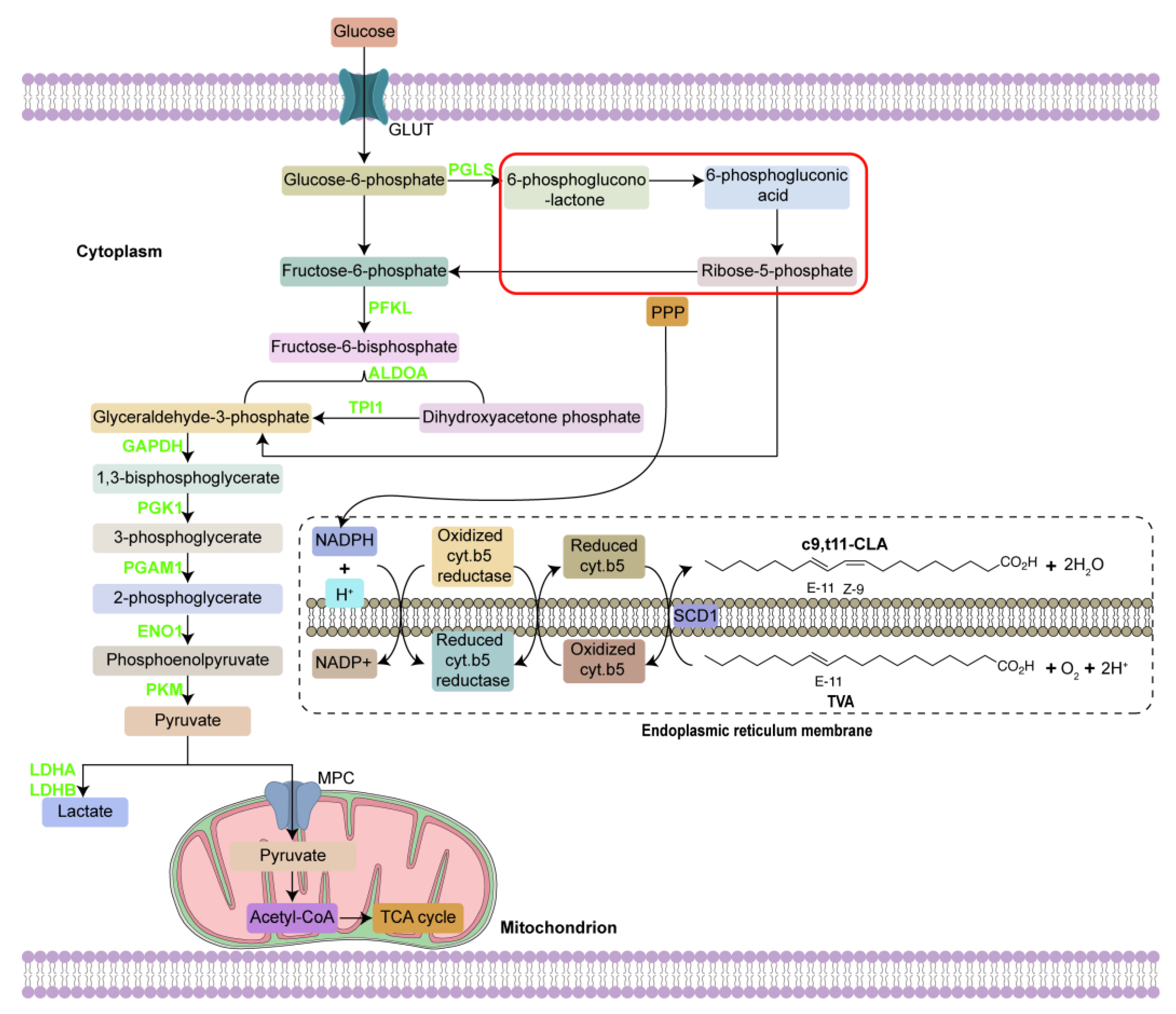
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavillonniere, F.; Chajes, V.; Martin, J.C.; Sébédio, J.L.; Lhuillery, C.; Bougnoux, P. Dietary purified cis9, trans11 conjugated linoleic acid isomer has anticarcinogenic properties in chemically induced mammary tumors in rats. Nutr. Cancer 2003, 45, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Huebners, S.M.; Olson, J.M.; Campbell, J.P.; Bishop, J.W.; Crump, P.M.; Cook, M.E. Low dietary c9t11-conjugated linoleic acid intake from dairy fat or supplements reduces inflammation in collagen-induced arthritis. Lipids 2016, 51, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Schaftenaa, F.; Frodermann, V.; Kuiper, J.; Lutgens, E. Atherosclerosis: The interplay between lipids and immune cells. Curr. Opin. Lipidol. 2016, 27, 209–215. [Google Scholar] [CrossRef]
- Wang, T.; Lee, H.G. Advances in Research on cis9, trans11 conjugated linoleic acid: A major functional conjugated linoleic acid isomer. Crit. Rev. Food Sci. 2015, 55, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Naito, T.; Nagao, Y.; Kabuyama, Y.; Hashimoto, K.; Azuma, N.; Maeda, I. Effect of increased feeding of dietary α-linolenic acid by grazing on formation of the cis9, trans11-18:2 isoform of conjugated linoleic acid in bovine milk. Anim. Sci. J. 2017, 88, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Robyn, B.; Robyn, F.; Orina, B. Atheroprotective effects of conjugated linoleic acid. Br. J. Clin. Pharmacol. 2017, 83, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Mosley, E.E.; Shafii, B.; Moate, P.J.; McGuire, M.A. cis9, trans11 Conjugated linoleic acid is synthesized directly from vaccenic acid in lactating dairy cattle. J. Nutr. 2006, 136, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Griinari, J.M.; Corl, B.A.; Lacy, S.H.; Chouinard, P.Y.; Nurmela, K.V.; Bauman, D.E. Conjugated linoleic acid is synthesized endogenously in lactating dairy cows by D9-desaturase. J. Nutr. 2000, 130, 2285–2291. [Google Scholar] [CrossRef]
- Kay, J.K.; Mackle, T.R.; Auldist, M.J.; Thomson, N.A.; Bauman, D.E. Endogenous synthesis of cis9, trans11 conjugated linoleic acid in dairy cows fed fresh pasture. J. Dairy Sci. 2004, 87, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.H.; Hyeong, S.K.; Young, Y.B.; Yeonhwa, P. Overview of conjugated linoleic acid formation and accumulation in animal products. Livest. Sci. 2016, 195, 105–111. [Google Scholar] [CrossRef]
- Iwai, T.; Kume, S.; Chin-Kanasaki, M.; Kuwagata, S.; Araki, H.; Takeda, N.; Sugaya, T.; Uzu, T.; Maegawa, H.; Araki, S.I. Stearoyl-CoA desaturase-1 protects cells against lipotoxicity-mediated apoptosis in proximal tubular cells. Int. J. Mol. Sci. 2016, 17, 1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengi, A.J.; Corl, B.A. Identification and characterization of a novel bovine stearoyl-CoA desaturase isoform with homology to human SCD5. Lipids 2007, 42, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Rezamand, P.; Watts, J.S.; Yavah, K.M.; Mosley, E.E.; Ma, L.Y.; Corl, B.A.; McGuire, M.A. Relationship between stearoyl-CoA desaturase 1 gene expression, relative protein abundance, and its fatty acid products in bovine tissues. J. Dairy Res. 2014, 81, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M.; Buhrow, S.A.; Kaestner, K.H.; Christy, R.J.; Sibley, E.; KellyJr, T.J.; Lane, M.D. Differentiation-induced gene expression in 3T3-L1 preadipocytes. Characterization of a differentially expressed gene encoding stearoyl-CoA desaturase. J. Biol. Chem. 1988, 263, 17291–17300. [Google Scholar] [CrossRef]
- Wang, L.Z.; You, J.H.; Zhong, B.S.; Ren, C.F.; Zhang, Y.L.; Li, M.; Zhang, G.M.; Jia, R.X.; Ying, S.J.; Wang, F. Scd1 mammary-specific vector constructed and overexpressed in goat fibroblast cells resulting in an increase of palmitoleic acid and oleic acid. Biochem. Biophys. Res. Commun. 2013, 443, 389–394. [Google Scholar] [CrossRef]
- Yao, D.W.; Luo, J.; He, Q.Y.; Shi, H.B.; Li, J.; Wang, H.; Xu, H.F.; Chen, Z.; Yi, Y.Q.; Loor, J.J. SCD1 alters long-chain fatty acid (LCFA) composition and its expression is directly regulated by SREBP-1 and PPARgamma 1 in dairy goat mammary cells. J. Cell Physiol. 2017, 232, 635–649. [Google Scholar] [CrossRef]
- Lounis, M.A.; Escoula, Q.; Veillette, C.; Bergeron, K.F.; Ntambi, J.M.; Mounier, C. SCD1 deficiency protects mice against ethanol-induced liver injury. BBA-Mol. Cell Biol. Lipids 2016, 1861, 1662–1670. [Google Scholar] [CrossRef] [Green Version]
- Paton, C.M.; Ntambi, J.M. Biochemical and physiological function of stearoyl-CoA desaturase. Am. J. Physiol.-Endoc. Metab. 2009, 297, E28–E37. [Google Scholar] [CrossRef] [Green Version]
- Jedrzejczak, M.; Szatkowska, I. Bovine mammary epithelial cell cultures for the study of mammary gland functions. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Lee, S.B.; Hwang, J.H.; Lim, J.N.; Jung, U.S.; Kim, M.J.; Kim, H.S.; Kang, S.H.; Choi, J.S.; Lee, S.G.; et al. Proteomic analysis reveals PGAM1 altering cis9, trans11 conjugated linoleic acid synthesis in bovine mammary gland. Lipids 2015, 50, 469–481. [Google Scholar] [CrossRef]
- Mitz, C.A.; Viloria-Petit, A.M. Contrasting effects of transforming growth factor β1 on programmed cell death of bovine mammary epithelial cell lines MAC-T and BME-UV1. J. Dairy Sci. 2020, 103, 5532–5549. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Lim, J.N.; Bok, J.D.; Kim, J.H.; Kang, S.K.; Lee, S.B.; Hwang, J.H.; Lee, K.H.; Kang, H.S.; Choi, Y.J.; et al. Association of protein expression in isolated milk epithelial cells and cis9, trans11 conjugated linoleic acid proportions in milk from dairy cow. J. Sci. Food Agric. 2013, 94, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Lee, H.G. Responses of MAC-T cells to inhibited stearoyl-CoA desaturase 1 during cis9, trans11 conjugated linoleic acid synthesis. Lipids 2018, 53, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.L.; Fujimoto, B.A.; Jiménez-Flores, R.; Peterson, D.G. Growth hormone alters lipid composition and increases the abundance of casein and lactalbumin mRNA in the MAC-T cell line. J. Dairy Res. 2010, 77, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, L.; Leroux, C.; Bonnet, M.; Rouel, J.; Martin, P.; Chilliard, Y. Expression and nutritional regulation of lipogenic genes in mammary gland and adipose tissues of lactating goats. J. Dairy Res. 2005, 72, 250–255. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Lim, J.N.; Choi, S.H.; Kang, H.S.; Lee, H.G. Comparative studies on derivatization methods of single or mixed fatty acids. Food Sci. Biotechnol. 2013, 22, 1573–1579. [Google Scholar] [CrossRef]
- Wang, T.; Oh, J.J.; Lim, J.N.; Hong, J.E.; Kim, J.H.; Kim, J.H.; Kang, H.S.; Choi, Y.J.; Lee, H.G. Effects of lactation stage and individual performance on milk cis9, trans11 conjugated linoleic acids content in dairy cows. Asian-Australas. J. Anim. Sci. 2013, 26, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Min, C.W.; Park, J.; Bae, J.W.; Agrawal, G.K.; Rakwal, R.; Kim, Y.; Yang, P.F.; Kim, S.T.; Gupta, R. In-depth investigation of low-abundance proteins in matured and filling stages seeds of glycine max employing a combination of protamine sulfate precipitation and TMT-based quantitative proteomic analysis. Cells 2020, 9, 1517. [Google Scholar] [CrossRef]
- Duan, Z.Q.; Yuan, C.; Han, Y.F.; Zhou, L.; Zhao, J.F.; Ruan, Y.; Chen, J.Q.; Ni, M.M.; Ji, X.Q. TMT-based quantitative proteomics analysis reveals the attenuated replication mechanism of Newcastle disease virus caused by nuclear localization signal mutation in viral matrix protein. Virulence 2020, 11, 607–635. [Google Scholar] [CrossRef]
- Zhao, L.; Geng, T.T.; Sun, K.C.; Su, S.; Zhao, Y.; Bao, N.; Pan, L.; Sun, H.; Li, M.Y. Proteomic analysis reveals the molecular mechanism of Hippophae rhamnoides polysaccharide intervention in LPS-induced inflammation of IPEC-J2 cells in piglets. Int. J. Biol. Macromol. 2020, 164, 3294–3304. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Pascovici, D.; Wu, J.X.; Chick, J.; Wu, Y.; Cooke, B.; Haynes, P.; Molloy, M.P. TMT one-stop shop: From reliable sample preparation to computational analysis platform. In Proteome Bioinformatics; Keerthikumar, S., Mathivanan, S., Eds.; Humana Press: New York, NY, USA, 2010; Volume 1549. [Google Scholar] [CrossRef]
- Ma, C.; Wang, W.W.; Wang, Y.D.; Sun, Y.; Kang, L.; Zhang, Q.; Jiang, Y.L. TMT-labeled quantitative proteomic analyses on the longissimus dorsi to identify the proteins underlying intramuscular fat content in pigs. J. Proteom. 2019, 213, 103630. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.Y.; Guo, D.Q.; Yu, X.; Wang, S.Y.; Liu, T.H. TMT-based proteomics analysis of the anti-hepatocellular carcinoma effect of combined dihydroartemisinin and sorafenib. Biomed. Pharmacother. 2020, 126, 109862. [Google Scholar] [CrossRef]
- Sun, J.; Jiang, T.C.; Gu, F.; Ma, D.H.; Liang, J.M. TMT-based proteomic analysis of plasma from children with rolandic epilepsy. Dis. Markers 2020, 2020, 8840482. [Google Scholar] [CrossRef]
- Jia, B.Y.; Xiang, D.C.; Liu, S.N.; Zhang, B.; Shao, Q.Y.; Hong, Q.H.; Quan, G.B.; Wu, G.Q. TMT-based quantitative proteomic analysis of cumulus cells derived from vitrified porcine immature oocytes following in vitro maturation. Theriogenology 2020, 152, 8–17. [Google Scholar] [CrossRef]
- Wang, Z.K.; Liu, F.J.; Ye, S.L.; Jiang, P.; Yu, X.C.; Xu, J.; Du, X.; Ma, L.; Cao, H.G.; Yuan, C.; et al. Plasma proteome profiling of high-altitude polycythemia using TMT-based quantitative proteomics approach. J. Proteom. 2018, 194, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W.; Bandeira, N.; Sharma, V.; Perez-Riverol, Y.; Carver, J.J.; Kundu, D.J.; García-Seisdedos, D.; Jarnuczak, A.F.; Hewapathirana, S.; Pullman, B.S.; et al. The ProteomeXchange consortium in 2020: Enabling ‘big data’ approaches in proteomics. Nucleic Acids Res. 2020, 48, D1145–D1152. [Google Scholar] [CrossRef] [Green Version]
- Kan, L.B.; Cui, D.Z.; Chai, Y.Y.; Ma, L.; Li, X.Y.; Zhao, M. TMT-based quantitative proteomic analysis of antitumor mechanism of Sporisorium reilianum polysaccharide WM-NP-60 against HCT116 cells. Int. J. Biol. Macromol. 2020, 165, 1755–1764. [Google Scholar] [CrossRef]
- Chen, C.; Hou, J.; Tanner, J.J.; Cheng, J.L. Bioinformatics methods for mass spectrometry-based proteomics data analysis. Int. J. Mol. Sci. 2020, 21, 2873. [Google Scholar] [CrossRef] [Green Version]
- Xue, T.; Liu, P.; Zhou, Y.; Liu, K.; Yang, L.; Moritz, R.L.; Yan, W.; Xu, L.X. Interleukin-6 induced “Acute” phenotypic microenvironment promotes Th1 anti-tumor immunity in cryo-thermal therapy revealed by shotgun and parallel peaction monitoring proteomics. Theranostics 2016, 6, 773–794. [Google Scholar] [CrossRef]
- Yan, X.Y.; Wu, Y.J.; Zhong, F.J.; Jiang, Q.Y.; Zhou, T.T.; Guo, Y.F.; Yang, X.R.; Liang, J.; Liao, D.J.; Lan, G.Q. iTRAQ and PRM-based quantitative proteomics in T2DM-susceptible and-tolerant models of Bama mini-pig. Gene 2018, 675, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Singh, N.; Long, C.; Smith, S.B. The lentiviral system construction for high expressed porcine stearoyl CoA desaturase-l and functional characterization in stably transduced porcine swine kidney cells. Lipids 2018, 53, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, M.; Bharathi, M.; Reddy, M.R.G.; Kumar, M.S.; Putcha, U.K.; Vajreswari, A.; Jeyakumar, S.M. Carrot juice administration decreases liver stearoyl-CoA desaturase I and improves docosahexaenoic acid levels but not steatosis in high fructose diet-fed weanling wistar rats. Prev. Nutr. Food Sci. 2016, 21, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.; Miyazaki, M.; Socci, N.D.; Hagge-Greenberg, A.; Liedtke, W.; Soukas, A.A.; Sharma, R.; Hudgins, L.C.; Ntambi, J.M.; Friedman, J.M. Role for stearoyl-coa desaturase eptin-mediated weight loss. Science 2002, 297, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wang, Y.N.; Ma, H.; He, Z.H.; Tang, Y.; Guo, L.; Liu, Y.; Ding, M.; Qian, S.W.; Tang, Q.Q. SCD1 promotes lipid mobilization in subcutaneous white adipose tissue. J. Lipid Res. 2020, 61, 1589–1604. [Google Scholar] [CrossRef]
- Teng, Z.X.; Guo, M.Z.; Liu, X.Y.; Tian, Z.; Che, K. Revealing protein functions based on relationships of interacting proteins and GO terms. J. Biomed. Semant. 2017, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Conte, G.; Mele, M.; Chessa, S.; Castiglioni, B.; Serra, A.; Pagnacco, G.; Secchiari, P. Diacylglycerol acyltransferase 1, stearoyl-CoA desaturase 1, and sterol regulatory element binding protein 1 gene polymorphisms and milk fatty acid composition in Italian Brown cattle. J. Dairy Sci. 2010, 93, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Yan, P.; Eng, O.C.; Yu, C.J. A review on the expression and metabolic features of orphan human cytochrome P450 2S1 (CYP2S1). Curr. Drug Metab. 2018, 19, 917–929. [Google Scholar] [CrossRef]
- Bray, G.A.; Champagne, C.M. Obesity and the metabolic syndrome: Implications for dietetics practitioners. J. Am. Diet. Assoc. 2004, 104, 86–89. [Google Scholar] [CrossRef]
- Ebert, K.; Witt, H. Fructose malabsorption. Mol. Cell. Pediatr. 2016, 3, 10. [Google Scholar] [CrossRef]
- Roy, K.; Wu, Y.; Meitzler, J.L.; Juhasz, A.; Liu, H.; Jiang, G.J.; Lu, J.M.; Antony, S.; Doroshow, J.H. NADPH oxidases and cancer. Clin. Sci. 2015, 128, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Opielka, M.; Sobocki, B.; Mierzejewska, P.; Smolenski, R.T. The effect of trehalose on intracellular and extracellular nucleotide metabolism. A pilot study. Nucleosides Nucleotides Nucleic Acids 2020, 39, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Kuhl, W.; Gelbart, T. 6-Phosphogluconolactonase deficiency, a hereditary erythrocyte enzyme deficiency: Possible interaction with glucose-6-phosphate dehydrogenase deficiency. Proc. Natl. Acad. Sci. USA 1985, 82, 3876–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.Z.; Chen, J.; Li, Y.S.; Ye, Z.T.; Tian, X.B.; Wei, Y.; Hao, Z.C.; Pan, Y.; Zhou, H.L. 6-Phosphogluconolactonase promotes hepatocellular carcinogenesis by activating pentose phosphate pathway. Front. Cell Dev. Biol. 2021, 9, 753196. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.L.; Zhang, Y.; Cai, Y.; Liu, R.J.; Lu, M.L.; Li, T.Z.M.; Fu, Y.; Guo, M.; Huang, H.C.; Ou, Y.F.; et al. A20 targets PFKL and glycolysis to inhibit the progression of hepatocellular carcinoma. Cell Death Dis. 2020, 11, 89. [Google Scholar] [CrossRef]
- Mor, I.; Cheung, E.C.; Vousden, K.H. Control of glycolysis through regulation of PFK1: Old friends and recent additions. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 211–216. [Google Scholar] [CrossRef]
- Moreno-Sanchez, R.; Rodriguez-Enriquez, S.; Marin-Hernandez, A.; Saavedra, E. Energy metabolism in tumor cells. FEBS J. 2007, 274, 1393–1418. [Google Scholar] [CrossRef]
- Huang, Z.K.; Hua, Y.B.; Tian, Y.; Qin, C.; Qian, J.; Bao, M.L.; Liu, Y.Y.; Wang, S.Q.; Cao, Q.; Ju, X.B.; et al. High expression of fructose-bisphosphate aldolase A induces progression of renal cell carcinoma. Oncol. Rep. 2018, 39, 2996–3006. [Google Scholar] [CrossRef] [Green Version]
- Vladimir, F.L.; Irina, V.G.; Boris, A.M. Glyceraldehyde-3-phosphate dehydrogenase is a multifaceted therapeutic target. Pharmaceutics 2020, 12, 416. [Google Scholar] [CrossRef]
- Qian, X.; Li, X.J.; Cai, Q.S.; Zhang, C.B.; Yu, Q.J.; Jiang, Y.H.; Lee, J.H.; Hawke, D.; Wang, Y.G.; Xia, Y.; et al. Phosphoglycerate kinase 1 phosphorylates beclin1 to induce autophagy. Mol. Cell 2017, 65, 917–931. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.H.; Jiang, L.L.; Cao, Y.; Ye, D.Y.; Zhou, L. The design and synthesis of N-xanthone benzenesulfonamides as novel phosphoglycerate mutase 1 (PGAM1) inhibitors. Molecules 2018, 23, 1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, G.; Wu, A.G.; Chen, X.L.; Tian, Y.; Lin, X.K. Enolase 1, a moonlighting protein, as a potential target for cancer treatment. Int. J. Biol. Sci. 2021, 17, 3981–3992. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Sugito, N.; Shinohara, H.; Kuranaga, Y.; Inomata, Y.; Komura, K.; Uchiyama, K.; Akao, Y. Organ-specific microRNAs (MIR122, 137, and 206) contribute to tissue characteristics and carcinogenesis by regulating pyruvate kinase M1/2 (PKM) expression. Int. J. Mol. Sci. 2018, 19, 1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilpe, S.V.; Koornstra, R.; Brok, M.D.; Groot, J.W.D.; Blank, C.; Vries, J.D.; Gerritsen, W.; Mehra, N. Lactate dehydrogenase: Actate dehydrogenase: A marker of diminished antitumor immunity. OncoImmunology 2020, 9, 1731942. [Google Scholar] [CrossRef] [PubMed] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, L.; Bai, G.; Wang, C.; Dong, J.; Liu, Y.; Sun, Z.; Zhen, Y.; Qin, G.; Zhang, X.; Demelash, N.; et al. Proteomics Insights into the Gene Network of cis9, trans11-Conjugated Linoleic Acid Biosynthesis in Bovine Mammary Gland Epithelial Cells. Animals 2022, 12, 1718. https://doi.org/10.3390/ani12131718
Peng L, Bai G, Wang C, Dong J, Liu Y, Sun Z, Zhen Y, Qin G, Zhang X, Demelash N, et al. Proteomics Insights into the Gene Network of cis9, trans11-Conjugated Linoleic Acid Biosynthesis in Bovine Mammary Gland Epithelial Cells. Animals. 2022; 12(13):1718. https://doi.org/10.3390/ani12131718
Chicago/Turabian StylePeng, Liying, Ge Bai, Chunzheng Wang, Jianan Dong, Yongjun Liu, Zhe Sun, Yuguo Zhen, Guixin Qin, Xuefeng Zhang, Natnael Demelash, and et al. 2022. "Proteomics Insights into the Gene Network of cis9, trans11-Conjugated Linoleic Acid Biosynthesis in Bovine Mammary Gland Epithelial Cells" Animals 12, no. 13: 1718. https://doi.org/10.3390/ani12131718
APA StylePeng, L., Bai, G., Wang, C., Dong, J., Liu, Y., Sun, Z., Zhen, Y., Qin, G., Zhang, X., Demelash, N., & Wang, T. (2022). Proteomics Insights into the Gene Network of cis9, trans11-Conjugated Linoleic Acid Biosynthesis in Bovine Mammary Gland Epithelial Cells. Animals, 12(13), 1718. https://doi.org/10.3390/ani12131718