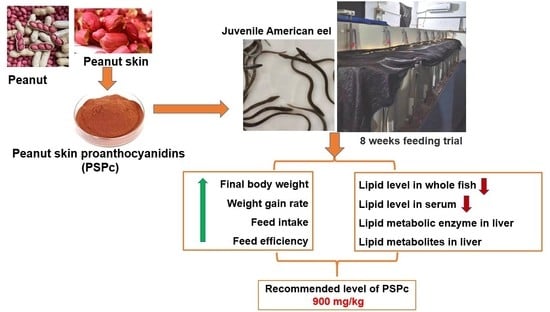
4.1. Effects of Dietary PSPc Supplementation on Growth Performance and Proximate Composition of Whole Fish of the Juvenile American Eel
In the present study, appropriate levels of dietary PSPc supplementation increased the WGR, SGR, FE, and FR of the juvenile American eel. Our results indicate that dietary PSPc supplementation might have a positive impact on growth performance. Wang et al. [
15] demonstrated that dietary 400 mg/kg GSP could improve the WGR of the American eel, which is in line with the results of the present study. Similarly, the WGR and FE of tilapia were improved by 200–800 mg/kg dietary GSP [
4]. In the studies of rainbow trout, 1000 mg/kg grape seed extract supplemented in the diet improved WGR, SGR, and FE [
6], while 1000 mg/kg grape seed oil increased WGR and SGR [
5]. Mehrinakhi et al. [
8] found that 10–30 g/kg dietary grape seed extract improved the WGR and FE of common carp. Furthermore, dietary GSP supplementation could ameliorate the growth retardation of the American eel exposed to dietary histamine stress [
7] and pearl gentian grouper (
Epinephelus fuscoguttatus female ×
Epinephelus lanceolatus male) and tilapia exposed to dietary cadmium stress [
20,
21]. The possible reasons for the growth-promoting effect of PSPc might be the same as the GSP—PSPc could be absorbed in the anterior intestine and quickly metabolized to induce the synthesis of proteins associated with cytoskeletal function, resulting in an increase in the small intestine absorptive surface, and further promote the absorption of nutrients [
22]. The absorbed PSPc could improve the intestinal digestive enzyme activities and positively regulate the intestinal microbiota to maintain intestinal health and eventually improve growth performance [
10]. In the present trial, the best growth performance of the American eel was found in the group fed 900 mg/kg PSPc, which was higher than the optimal level of GSP at 400 mg/kg. The difference in optimal level between PSPc and GSP might be due to the different structure and composition of proanthocyanidins. The catechin and epicatechin are the basic structural units of proanthocyanidins. The quantity and degree of polymerization of catechin and epicatechin are different between PSPc and GSP [
23]. The proanthocyanidins in PSPc belong to the A-type, and the proanthocyanidins in GSP belong to the B-type. The main difference between A-type and B-type proanthocyanidins is the interflavanic linkages, and the A-type at least has one double linkage consisting of a C-C bond and an additional ether bond.
Lower levels of crude lipids in whole fish were found in juvenile American eels in PSPc600, PSPc900, and PSPc1200 groups. This result was similar to the study of tilapia fed 200–800 mg/kg GSP [
4]. The decreasing lipid levels in whole fish indicated that the PSPc might have a hypolipidemic effect in juvenile American eel. However, 200 mg/kg dietary GSP could increase the fillet protein content of common carp, and 500 mg/kg grape seed extract supplemented in the diet enhanced the protein level of the whole body of rainbow trout [
6,
9]. The difference in the above results might be due to fish species, resources and supplementation levels of proanthocyanidins, and dietary nutrient levels.
4.2. Effects of Dietary PSPc Supplementation on Serum Lipids and Lipid Metabolic Enzymes of the Juvenile American Eel
The levels of TC, TG, LDL-C, and HDL-C in serum are relevant to lipid metabolism; excessive TC and TG in the blood are considered to result in metabolic diseases [
3]. In the present study, there were lower levels of TC, TG, and LDL-C and higher HDL-C levels in the PSPc-treated groups, and there was no other report about the effect of PSPc on serum lipid in fish. The results of the present trial were consistent with the study of American eels fed 400 mg/kg GSP [
3]. Similar results were also found in common carp and tilapia fed 200 mg/kg or 400 kg/kg GSP; the levels of TC and TG in the serum of those two fish species were found to be decreased significantly [
4,
9]. However, GSP could not reduce the TG levels in the serum of rainbow trout [
24]. The lipid-reduced mechanisms in serum of PSPc might be related to inhibiting cholesterol absorption and synthesis, stimulating LDL-C receptor transcription, and downregulating certain genes of TG synthesis, along with upregulating some genes of lipid catabolism [
11,
25].
The liver is an important organ for fatty acid oxidation [
26]. The LPL and HL are parts of lipolytic enzymes in the fish liver and are collectively known as TL. The LPL can hydrolyze TG-rich lipoprotein and produce free fatty acid. The HL can be used as a ligand to promote LDL and chylomicron remnant entry into liver cells [
26,
27]. The FAS and ACC are related to lipid synthesis in the liver. The FAS is a key enzyme to regulate the de novo biosynthesis of long-chain fatty acids. The ACC is mainly responsible for the first step of de novo fatty acid biosynthesis [
28]. In the present study, there were increasing activities of LPL, HL, and TL and decreasing levels of FAS and ACC in the liver of the PSPc-treated groups, and little information was available about the effect of PSPc on lipid metabolism enzymes in the fish liver. However, Lu et al. revealed that GSP might reduce fat accumulation and fatty acid synthesis by upregulating the LPL gene expression and downregulating the gene expression of FAS and ACC in the liver after gavage of grass carp with 250 mg/kg GSP for 3 h [
11].
4.3. Effects of Dietary PSPc Supplementation on Metabolites and Metabolic Pathways Involved in Lipid Metabolism in the Liver of the Juvenile American Eel
In recent years, lipidomics has been widely applied to reveal the internal changes in organisms through the comprehensive analysis of subtle changes in lipids at the molecular level [
29]. The liver is the central organ of lipid metabolism in fish species [
30]. In the present study, 900 mg/kg dietary PSPc upregulated the levels of PC and PE, downregulated the levels of Cer and 2-LPC, and mainly affected the pathways of autophagy, sphingolipid metabolism, glycerophospholipid metabolism, and linoleic acid metabolism in the liver.
Autophagy is an intracellular degradation system and is key to maintaining homeostasis in the cells [
31]. A previous study reported that the autophagosome could transfer lipid droplets to lysosomes and inhibit lipid accumulation [
32]. On the contrary, the inhibition of autophagy triggered the TG increase and lipid droplet accumulation in hepatocytes [
33]. The LC3 is an autophagy marker protein that reflects the procession of autophagy. The PE plays a role in autophagy by covalently attaching to modify LC3 and is located on the autophagosome membrane, consequently mediating autophagosome formation and elongation [
34,
35]. Therefore, the upregulated PE level in the autophagy pathway is likely to induce adipocyte autophagy and reduce lipid accumulation in the liver. Furthermore, a study on zebrafish (
Danio rerio) showed that polyphenols might induce the autophagy process, reduce lipid droplet formation, and promote lipid droplet breakdown in the liver [
35], which was similar to the present study. These results suggest that PSPc might have positive regulatory roles in lipid metabolism through autophagy.
Sphingolipids are the bioactive lipids that regulate the major biological processes and contribute to the arrangement of membrane lipid domains [
36], including four subclasses: phosphosphingolipid, glycosphingolipids, sphingoid bases, and Cer. A lipidomic analysis showed that sphingolipid content (especially Cer) might be strongly correlated with the TG level in the liver, and a significant increase in hepatic Cer content was observed in rats fed with a high-fat diet [
37]. In addition, ceramide could activate the pro-inflammatory pathways in macrophages and secrete pro-inflammatory cytokines TNF-α and interleukin-1 [
36]. The downregulation of Cer in the liver of the PSPc900 group suggested that there might be a reduction in the liver lipid accumulation and inflammation by inhibiting the sphingolipid metabolism.
Glycerophospholipids, such as PC and PE, are major components of biological membranes. PC seems to be the predominant phospholipid class in lipoproteins of fish [
38,
39]. PE is one of the components of the cell membrane system, and the majority of PE is located in the inner lipid bilayer structure [
40]. A recent study showed that PC could increase the expression of LPL and hormone-sensitive lipase and decrease the FAS expression in the liver of tilapia [
41]. PE is required for the function and integrity of mitochondrial membranes [
34] and is used to help TC metabolism by transforming into PC [
42]. LPC is an important signaling molecule for many diseases and could produce pro-inflammatory actions, including the release of chemotactic factors and the production of reactive oxygen species [
43,
44]. In this study, the glycerophospholipid metabolism pathway was significantly affected by involving the downregulated level of the 2-LPC in the liver of the American eels in the PSP900 group. Similarly, the downregulation of hepatic LPC was also found in black seabream fed a diet with polyphenol supplementation [
44]. In general, 900 mg/kg dietary PSPc could modulate the glycerophospholipid metabolism by upregulating levels of PC and PE and downregulating 2-LPC levels in the liver of juvenile American eels, which might be beneficial in decreasing liver lipid accumulation.
As an essential fatty acid for vertebrates, linoleic acid was found to accelerate fatty acid oxidation [
45]. It has been reported that the content of PC in rat liver microsomes is positively proportional to the level of linoleic acid [
46]. Therefore, the upregulation of PC might have enhanced the anabolism of linoleic acid in the present study, and the increased linoleic acid level could further promote the proliferation and differentiation of fatty cells to exert hypolipidemic effects [
45,
46]. In addition, linoleic acid could increase the number of LDL receptors and enhance their affinity with LDL to accelerate LDL degradation to reduce the TC level [
46], which supports the results of the decreased TC level in the serum of the PSPc900 group.
In addition, PACs from grape seed were reported to interfere with lipid metabolism involved in intestinal absorption of lipids, changes in the gut microbiota to enhance microbial propionate production, and the secretion of chylomicrons and lipoproteins by the intestine and liver of terrestrial animals [
1,
47]. The mechanism of lipid metabolism regulation by dietary PSPc supplementation should be clarified in this regard in future studies on the American eel.






{kind=link}
{kind=link}
{kind=link}
{kind=link}